李斯特菌噬菌體裂解酶的研究進(jìn)展
2016-04-27 03:30:55劉珊娜范志華孫溪付榮霞吳海清
天津農(nóng)學(xué)院學(xué)報(bào) 2016年1期
關(guān)鍵詞:研究進(jìn)展
劉珊娜,范志華,孫溪,付榮霞,吳海清
(天津農(nóng)學(xué)院 食品科學(xué)與生物工程學(xué)院,天津市農(nóng)副產(chǎn)品深加工技術(shù)工程中心,天津 300384)
?
李斯特菌噬菌體裂解酶的研究進(jìn)展
劉珊娜,范志華,孫溪,付榮霞,吳海清
(天津農(nóng)學(xué)院 食品科學(xué)與生物工程學(xué)院,天津市農(nóng)副產(chǎn)品深加工技術(shù)工程中心,天津 300384)
摘 要:單核細(xì)胞增生李斯特菌是重要的食源性致病菌,可通過污染的食品進(jìn)入人體,引發(fā)食品安全事件。噬菌體作為李斯特菌生物防治的有效手段,能夠特異性識(shí)別菌體細(xì)胞,造成細(xì)菌裂解死亡。噬菌體發(fā)揮抑菌活性的關(guān)鍵在于其雙鏈DNA所編碼的裂解酶(肽聚糖水解酶類)能夠在增殖過程的后期有效裂解宿主菌的細(xì)胞壁,將子代噬菌體釋放到胞外環(huán)境中。本文綜述了李斯特菌噬菌體裂解酶的種類、作用原理和結(jié)構(gòu)特征,并探討了裂解酶的應(yīng)用領(lǐng)域和未來發(fā)展方向。
關(guān)鍵詞:李斯特菌;噬菌體;裂解酶;研究進(jìn)展
李斯特菌(Listeria)是一種兼性厭氧的革蘭氏染色陽性短桿菌,存在于土壤、污水、飼料、動(dòng)物和污染的食品中。在目前已確定的李斯特菌8個(gè)種屬中,單核細(xì)胞增生李斯特菌(Listeria monocytogenes,LM)是致病力最強(qiáng)的細(xì)菌,也是迄今發(fā)現(xiàn)的唯一對(duì)人致病的菌種,其主要針對(duì)孕婦、新生兒、老年人和免疫功能低下的人群[1-2],致死率高達(dá)25%~30%。目前已知至少有13種血清型,引起食物中毒的血清型多為1/2和4b[3]。多種食品包括魚和海產(chǎn)品、生香腸、奶酪、加工的肉制品等易受到LM的污染[4],由于該菌對(duì)各種應(yīng)激(低溫、低pH、高鹽、低水分活度、氧化應(yīng)激等)有一定程度的耐受性,因而進(jìn)一步增加了對(duì)人類健康的危害性。
噬菌體能夠特異性地識(shí)別菌體細(xì)胞,侵入宿主體內(nèi),完成增殖過程后裂解宿主細(xì)胞,釋放子代噬菌體。噬菌體具有只針對(duì)活體細(xì)胞進(jìn)行侵染的特點(diǎn),因此在病原菌的檢測(cè)和食品安全的生物控制方面起著越來越積極的作用[5-6]。國外對(duì)于噬菌體的開發(fā)研究逐漸成為熱點(diǎn),已有包括美國Omnilytics和EBI等公司相關(guān)研究產(chǎn)品的報(bào)道[7]。我國的研究起步較晚,仍處于實(shí)驗(yàn)室階段。噬菌體發(fā)揮抑菌活性的關(guān)鍵在于其雙鏈DNA所編碼的裂解酶(肽聚糖水解酶類)能夠在噬菌體增殖過程的后期有效裂解宿主菌的細(xì)胞壁,將子代噬菌體釋放到胞外環(huán)境中[8-9]。對(duì)于噬菌體裂解酶的認(rèn)識(shí),將更有利于揭示噬菌體特異性的來源和進(jìn)化規(guī)律,以及其與宿主菌的作用方式,為食源性LM的檢測(cè)和感染的治療提供新的途徑和策略。
1 李斯特菌噬菌體
噬菌體是環(huán)境中大量存在的自我復(fù)制單元,可作用于特定的宿主細(xì)胞,其增殖過程可分為裂解途徑和溶原途徑。裂解途徑中,噬菌體通過裂解細(xì)胞釋放子代噬菌體;而在溶原途徑中,噬菌體的DNA與宿主染色體整合,感染菌體后,不造成細(xì)胞死亡。噬菌體被認(rèn)為是可生物降解的食品級(jí)無毒殺菌物質(zhì),開發(fā)的相關(guān)產(chǎn)品也被美國FDA 和EPA認(rèn)可為GRAS級(jí)[10]。與抗生素相比,噬菌體來源廣泛,增殖速度快,特異性好,不影響腸道正常菌群,安全性較高,使用成本較低[11]。1945年,Schultz[12]首次報(bào)道了噬菌體可以侵染李斯特菌屬細(xì)菌。直至今日,超過400多種李斯特菌噬菌體從500多種環(huán)境來源中獲得,基本屬于長尾噬菌體家族(Siphoviridae)或肌尾噬菌體家族(Myoviridae)。大部分李斯特菌噬菌體表現(xiàn)出圓形排列的基因組結(jié)構(gòu),大小在30~65 kb之間,含有雙鏈DNA負(fù)責(zé)編碼結(jié)構(gòu)基因,以及重組、復(fù)制、修復(fù)等功能基因,裂解基因和溶原控制基因(針對(duì)溫和噬菌體)[13]等。李斯特菌噬菌體可應(yīng)用于LM污染的控制,清除環(huán)境中的LM,如美國FDA批準(zhǔn)的ListShieldTM、LMP-102和ListexTMP100等噬菌體試劑用于即食食品中LM的控制[14-15]。
2 裂解酶的種類和作用原理
噬菌體雙鏈DNA編碼的裂解酶基因在噬菌體增殖過程中表達(dá),可以降解細(xì)菌細(xì)胞壁的肽聚糖層,在細(xì)胞壁上形成孔洞,最終使細(xì)胞裂解而釋放子代噬菌體。不同噬菌體的裂解酶在蛋白結(jié)構(gòu)和酶活性上存在較大差異,通常一種裂解酶只表現(xiàn)出一種水解活性。根據(jù)酶在細(xì)胞壁上的作用方式可分為5大類:(1)N-乙酰胞壁酸酶(N-acetylmuramidases),與溶菌酶類的機(jī)理相同;(2)N-乙酰氨基葡萄糖苷酶(endo-β-N-acetylglucosaminidases),可以在多糖鏈中切斷兩個(gè)β-1,4糖苷鍵中的一個(gè);(3)轉(zhuǎn)糖基酶(lytic transglycosylases),切斷肽聚糖中的多糖部分;(4)肽鏈內(nèi)切酶(endopeptidases),切斷多肽之間的連接;(5)N-乙酰胞壁酰-L-丙氨酸酰胺酶(N-acetylmuramoyl-L-alanine amidases),切斷糖苷和多肽之間的化學(xué)鍵[8,16]。裂解酶不含有分泌信號(hào)肽,依靠疏水性跨膜蛋白holin的作用通過細(xì)胞內(nèi)膜到達(dá)肽聚糖層。雖然holin的作用機(jī)理還未完全得到驗(yàn)證,但已證實(shí)裂解酶和holin分子都是裂解菌體、釋放子代噬菌體所必需的,它們的編碼基因位置相鄰組成了裂解基因簇[17]。在噬菌體增殖的后期,holin分子和裂解酶同時(shí)表達(dá)。裂解酶不斷在胞內(nèi)積累,通過holin分子在內(nèi)膜形成的孔道,到達(dá)肽聚糖層并迅速裂解細(xì)胞壁。但也有報(bào)道LM的A511噬菌體裂解酶基因上游并無編碼holin的基因,有可能存在其他的胞內(nèi)轉(zhuǎn)運(yùn)機(jī)理[18]。與裂解酶活性研究進(jìn)展相比,有關(guān)裂解酶在細(xì)胞壁上受體的研究較少。已有研究結(jié)果表明,LM噬菌體裂解酶PlyP35能夠識(shí)別LM細(xì)胞壁磷壁酸上的N-乙酰氨基葡萄糖(GLCNAC)殘基[19]。而對(duì)于LM噬菌體裂解酶Ply118、Ply511和PlyP40,其直接作為結(jié)合配體的則是肽聚糖自身結(jié)構(gòu)而非細(xì)胞壁磷壁酸[20]。
3 裂解酶的結(jié)構(gòu)特征
2002年,瑞士蘇黎世聯(lián)邦理工學(xué)院的Loessner課題組首次發(fā)表了LM噬菌體裂解酶的蛋白結(jié)構(gòu)域,提出裂解酶由N端的酶活性域(EAD)和C端的細(xì)胞表面結(jié)合域(CBD)組成[21]。EAD決定酶的作用方式,CBD作為一個(gè)模式結(jié)構(gòu)識(shí)別李斯特菌細(xì)胞表面的配體分子。不同LM噬菌體裂解酶的水解位點(diǎn)具有多樣性,是由N端的EAD蛋白決定的。裂解酶的作用范圍通常反映出菌株血清型的差異,這種差異性體現(xiàn)了CBD蛋白特異性識(shí)別宿主細(xì)胞配體的特點(diǎn)。對(duì)李斯特菌12種裂解酶基因序列比較研究表明,其在結(jié)合特異性、結(jié)合配體數(shù)量和親和力上存在高度多樣性[19]。如裂解酶Ply118僅對(duì)血清型為1/2的受試菌株表現(xiàn)出明顯的結(jié)合作用,而裂解酶Ply500則只針對(duì)血清型為4、5和6的受試菌株。CBD是裂解酶與李斯特菌細(xì)胞壁結(jié)合所必須的,能夠保證細(xì)胞裂解時(shí)裂解酶與底物的結(jié)合,避免裂解新的宿主細(xì)胞。CBD與李斯特菌的結(jié)合是一種非共價(jià)的快速結(jié)合,親和力甚至可以與成熟抗體的結(jié)合力相比。當(dāng)環(huán)境中存在CBD特異性抗體IgG時(shí),CBD仍能和目標(biāo)菌株細(xì)胞壁上的配體分子結(jié)合。由于酶的活性與結(jié)合蛋白的特異性可以獨(dú)立存在而互不影響,因此通過基因工程手段替換任一結(jié)構(gòu)域,可改變結(jié)合特性或酶活性。現(xiàn)已證實(shí),重組表達(dá)的裂解酶蛋白可以快速裂解并殺滅環(huán)境中的LM,同時(shí)表現(xiàn)出高度的熱穩(wěn)定性[22]。與綠色熒光蛋白融合表達(dá)的CBD具有結(jié)合LM的特性,可作為指示分子在熒光顯微鏡下檢測(cè)牛奶和奶酪樣品中人為污染的LM,起到快速、準(zhǔn)確的靶向作用[23]。
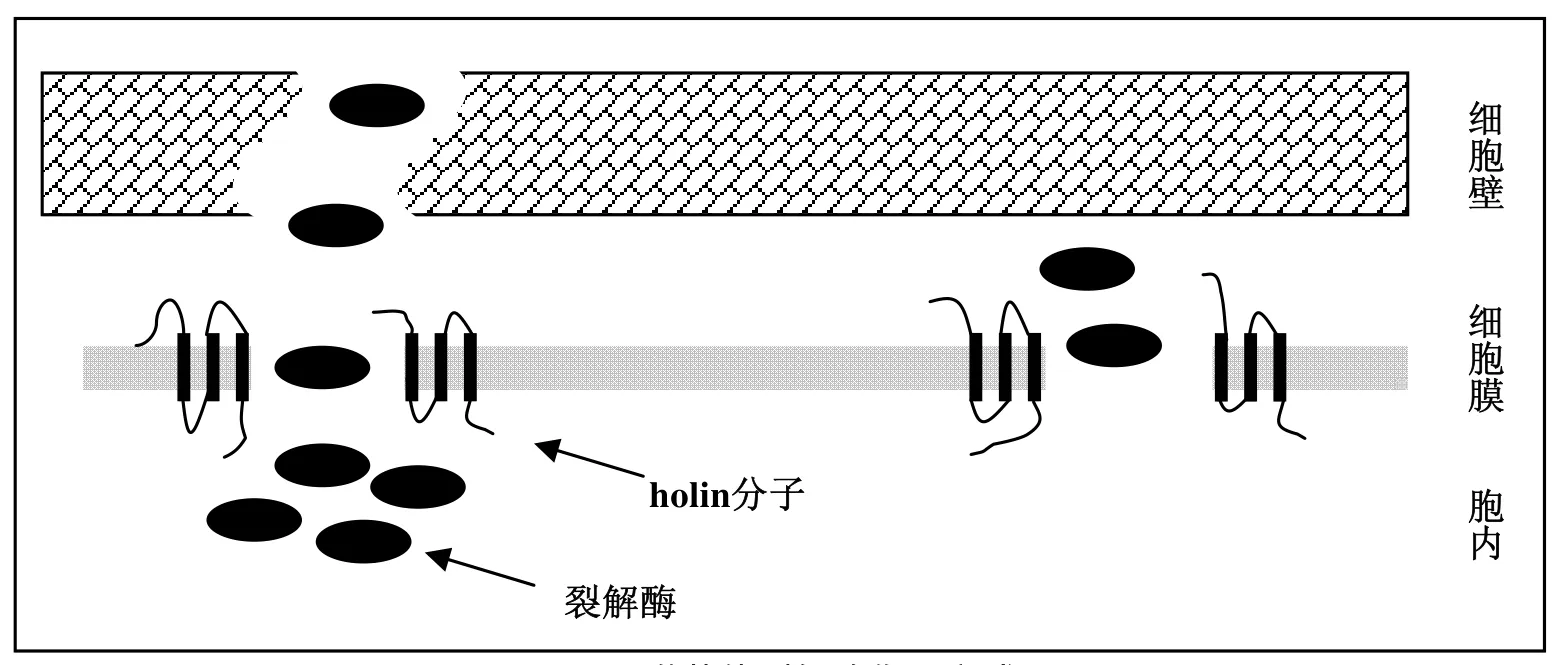
圖1 噬菌體裂解酶作用方式

圖2 李斯特菌噬菌體裂解酶結(jié)構(gòu)域和氨基酸序列相關(guān)性[23]
4 裂解酶的應(yīng)用
在國外,LM噬菌體已用于食品污染的控制、防腐保鮮、微生物檢測(cè)和感染的治療等方面,我國關(guān)于LM噬菌體方面的研究報(bào)道較少。與使用噬菌體相比,使用裂解酶對(duì)宿主菌作用更迅速,裂解譜更廣,可通過基因工程技術(shù)提高其產(chǎn)量。此外裂解酶的本質(zhì)是蛋白質(zhì),作為食品添加劑的使用更容易獲得法規(guī)許可。因此LM噬菌體裂解酶在食品和醫(yī)療等方面的探索研究也逐漸展開。如Gaeng等[24]將LM噬菌體裂解酶Ply118和Ply511在乳酸乳球菌中表達(dá),獲得了高活性的裂解酶,有效抑制了LM的生長。Schmelcher等人[23]將彩色熒光蛋白與LM噬菌體裂解酶CBD基因融合表達(dá),將這種熒光標(biāo)記的CBD蛋白加入到混合血清型的LM菌液中,可以通過多路成像技術(shù)分辨不同血清型的菌株。Kretzer等人[25]將熒光標(biāo)記技術(shù)與CBD-磁珠分離技術(shù)結(jié)合,檢測(cè)食品樣品(牛奶、奶酪)中人為污染的LM。在增菌培養(yǎng)6 h后,檢出下限在1~100 CFU/g。Walcher等[26]將CBD-磁珠與其他定量檢測(cè)方法共用,可以使檢測(cè)下限達(dá)到1 000 CFU/mL以下。目前,LM噬菌體裂解酶的應(yīng)用仍是研究的前沿領(lǐng)域,未來的研究有望為降低病原菌感染和食品安全風(fēng)險(xiǎn)提供更多的途徑。
5 結(jié)語
LM是重要的食源性致病菌,威脅人體健康并造成了嚴(yán)重經(jīng)濟(jì)損失。LM噬菌體裂解酶能夠從細(xì)胞內(nèi)水解LM菌體細(xì)胞壁,使菌體裂解。從人們開始對(duì)LM噬菌體裂解酶的研究以來,其晶體結(jié)構(gòu)、分子機(jī)制和作用特點(diǎn)不斷得到認(rèn)識(shí)。隨著對(duì)病原菌快速檢測(cè)技術(shù)和抗生素替代療法的需求不斷增加,基于噬菌體裂解酶的檢測(cè)和診斷系統(tǒng)必將得到更多關(guān)注。噬菌體基因組分析的開展,也將更有利于裂解酶安全性和作用效果的評(píng)價(jià)。未來將利用分子生物學(xué)手段進(jìn)行基因改造,闡明噬菌體裂解酶與宿主之間的相互作用,在實(shí)驗(yàn)室研究的基礎(chǔ)上進(jìn)一步評(píng)價(jià)噬菌體裂解酶在實(shí)際生產(chǎn)條件下的使用效果,拓寬對(duì)裂解酶生物學(xué)意義的認(rèn)識(shí),使噬菌體裂解酶的應(yīng)用技術(shù)得到完善和發(fā)展。
參考文獻(xiàn):
[1]Kuenne C,Billion A,Mraheil M A,et al.Reassessment of the Listeria monocytogenes pan-genome reveals dynamic integration hotspots and mobile genetic elements as major components of the accessory genome[J].BMC Genomics,2013,14:47.
[2]Winkelstr?ter L K,De Martinis E C P.Effect of bacteriocins and conditions that mimic food and digestive tract on biofilm formation,in vitro invasion of eukaryotic cells and internalin gene expression by Listeria monocytogenes[J].Probiotics & Antimicro Prot,2013,5(3):153-164.
[3]徐冬旸.食源性單核細(xì)胞增生李斯特菌對(duì)消毒劑及鎘協(xié)同耐受機(jī)制研究[D].廣州:華南理工大學(xué),2013.
[4]Gandhi M,Chikindas M L.Listeria:A foodborne pathogenthat knows how to suvive[J].Int J Food Microbiol,2007,113(1):1-15.
[5]Mahony J,McAuliffe O,Ross R P,et al.Bacteriophage as biocontrol agents of food pathogens[J].Curr Opin Biotechnol,2011,22(2):157-163.
[6]Coffey B,Mills S,Coffey A,et al.Phage and their lysins as biocontrol agents for food safety application[J].Annu Rev Food Sci Technol,2010,1(1):449-468.
[7]O’Flaherty S,Ross R P,Coffey A.Bacteriophage and their lysins for elimination of infectious bacteria[J].FEMS Microbiol Rev,2009,33(4):801-819.
[8]Borysowski J,Weber-Dabrowska B,Gorski A.Bacteriophage endolysins as a novel class of antibacterial agents[J].Exp Biol Med(Maywood),2006,231(4):366-377.
[9]Hermoso J A,Garcia J L,Garcia P.Taking aim on bacterial pathogens:from phage therapy to enzybiotics[J].Curr Opin Microbiol,2007,10(5):461-472.
[10]Arachchi G J G,Cridge A G,Dias-Wanigasekera B M,et al.Effectiveness of phages in the decontamination of Listeria monocytogenes adhered to clean stainless steel,stainless steel coated with fish protein,and as a biofilm[J].J Ind Microbiol Biotechnol,2013,40(10):1105-1116.
[11]Oliveira J,Castilho F,Cunha A,et al.Bacteriophage therapy as a bacterial control strategy in aquaculture[J].Aquacult Int,2012,20(5):879-910.
[12]Schultz E W.Listerella Infections:A review[J].Stanford Medical Bulletin,1945,3:135-151.
[13]Klumpp J,Loessner M J.Listeria phages:Genomes,evolution,and application[J].Bacteriophage,2013,3(3):e26861.
[14]Carlton R M,Noordman W H,Biswas B,et al.Bacteriophage P100 for control of Listeria monocytogenes in foods:genome sequence,bioinformatic analyses,oral toxicity study,and application[J].Regul Toxicol Pharmacol,2005,43(3):301-312.
[15]Bren L.Bacteria-eating virus approved as food additive[J].FDA Consum,2007,41(1):20-22.
[16]Loessner M J.Bacteriophage endolysins-current state of research and applications[J].Curr Opin Microbiol,2005,8(4):480-487.
[17]Young R Y.Bacteriophage holins:deadly diversity[J].J Mol Microbiol Biotechnol,2002,4(1):21-36.
[18]Loessner M J,Wendlinger G,Scherer S.Heterogeneous endolysins in Listeria monocytogenes bacteriophages:A new class of enzymes and evidence for conserved holin genes within the siphoviral lysis cassettes[J].Mol Microbiol,1995,16(6):1231-1241.
[19]Eugster M R,Haug M C,Huwiler S G,et al.The cell wall binding domain of Listeria bacteriophage endolysin PlyP35 recognizes terminal GlcNAc residues in cell wall teichoic acid[J].Mol Microbiol,2011,81(6):1419-1432.
[20]Eugster M R,Loessner M J.Wall teichoic acid restrict access of bacteriophage endolysin Ply118,Ply511,and PlyP40 cell wall binding domains to the Listeria monocytogenes peptidoglycan[J].J Bacteriol,2012,194(23):6498-6506.
[21]Loessner M J,Kramer K,Ebel F,et al.C-terminal domains of Listeria monocytogenes bacteriophage murein hydrolases determine specific recognition and high-affinity binding to bacterial cell wall carbohydrates[J].Mol Microbiol,2002,44(2):335-349.
[22]Schmelcher M,Waldherr F,Loessner M J,et al.Listeria bacteriophage peptidoglycan hydrolases feature high thermoresistance and reveal increased activity after divalent metal cation substitution[J].Appl Microbiol Biotechnol,2012,93(2):633-643.
[23]Schmelcher M,Shabarova T,Eugster M R,et al.Rapid multiplex detection and differentiation of Listeria cells by use of fluorescent phage endolysin cell wall binding domains[J].Appl Environ Microbiol,2010,76(17):5745-5756.
[24]Gaeng S,Scherer S,Neve H,et al.Gene cloning and expression and secretion of Listeria monocytogenes bacteriophage-lytic enzymes in Lactococcus lactis[J].Appl Environ Microbiol,2000,66(7):2951-2958.
[25]Kretzer J W,Lehmann R,Schmelcher M,et al.Use of high-affinity cell wall-binding domains of bacteriophage endolysins for immobilization and separation of bacterial cells[J].Appl Environ Microbiol,2007,73(6):1992-2000.
[26]Walcher G,Stessl B,Wagner M,et al.Evaluation of paramagnetic beads coated with recombinant Listeria phage endolysin-derived cell-wall-binding domain proteins for separation of Listeria monocytogenes from raw milk in combination with culture-based and real-time polymerase chain reaction-based quantification[J].Foodborne Pathog Dis,2010,7(9):1019-1024.
Research Progress in Listeria Phage Endolysins
LIU Shan-na,FAN Zhi-hua,SUN Xi,FU Rong-xia,WU Hai-qing
(Tianjin Engineering and Technology Research Center of Agricultural Products Processing,College of Food Science and Bioengineering,Tianjin Agricultural University,Tianjin 300384,China)
Abstract:Listeria monocytogenes is an important genus of food-borne pathogens.It may transmit into human body through contaminated food and cause food safety issue.Bacteriophages are effective means for Listeria bio-control,which can specifically recognize bacterial cells and lead to bacterial lysis.The key to antibacterial activities of bacteriophages relies on double stranded DNA-encoded endolysins,the murein enzymes which degrade host cell wall and result in release of phage progeny to the extracellular environment at the end of the phage lytic multiplication cycle.This article reviews the category of Listeria phage endolysins,the mechanism and structural feature of them,and discusses the application area and direction for development of endolysins in the future.
Key words:Listeria; bacteriophage; endolysin; research progress
作者簡介:劉珊娜(1984-),女,天津市人,講師,博士,主要從事食品微生物研究。E-mail:shannaliu@tjau.edu.cn。
基金項(xiàng)目:天津市科技支撐計(jì)劃項(xiàng)目“基于噬菌體蛋白的李斯特菌快速檢測(cè)技術(shù)及試紙的研發(fā)”(14ZCZDNC00003)
收稿日期:2015-08-18
文章編號(hào):1008-5394(2016)01-0062-04
中圖分類號(hào):S852.61
文獻(xiàn)標(biāo)識(shí)碼:A
猜你喜歡
現(xiàn)代畜牧科技(2021年8期)2021-10-13 07:21:46
昆明醫(yī)科大學(xué)學(xué)報(bào)(2021年3期)2021-07-22 07:40:08
心肺血管病雜志(2019年9期)2019-12-09 08:34:04
制造技術(shù)與機(jī)床(2019年10期)2019-10-26 02:47:12
中成藥(2017年9期)2017-12-19 13:34:44
老年醫(yī)學(xué)與保健(2017年6期)2017-02-06 05:30:03
中國塑料(2016年4期)2016-06-27 06:33:22
中國醫(yī)藥導(dǎo)報(bào)(2015年27期)2015-02-28 22:08:05
西南軍醫(yī)(2015年2期)2015-01-22 09:09:38
西南軍醫(yī)(2015年1期)2015-01-22 09:08:36
