荔枝無核和焦核機理的研究進展
2016-05-30 07:04:05李明芳盧誠劉興地王向社鄭學勤
熱帶作物學報 2016年5期
李明芳 盧誠 劉興地 王向社 鄭學勤

摘 要 對荔枝無核和焦核機理的研究概況作了綜述。指出大孢子發育異常無法完成受精和單性結實是導致荔枝無核的主要原因,今后對無核荔枝無核機理的研究應該著重從大孢子敗育和單性結實發生分子機理的角度展開,重點闡明無核荔枝發生大孢子敗育、受精被阻和單性結實的分子途徑。科技界對荔枝焦核機理的研究主要從組織細胞水平、生理生化角度和分子水平進行:從細胞水平看,主要是胚乳過早解體和胚在胚囊外發育引起合子或胚早期敗育,進而導致胚珠敗育形成焦核;從生理生化角度看,植物內源激素、多胺、酚類物質以及可溶性蛋白含量都與胚珠敗育、焦核形成有關;從分子水平看,兩條特異片段OPL-12-1 645 bp與OPL-12-722 bp,可能與荔枝的焦核基因相關。認為今后對荔枝焦核機理的研究應該著重于合子期到心形胚期,從分子水平上闡明究竟是哪些基因異常表達使合子分裂受阻、使胚乳過早解體,使球形胚、心形胚形成受阻。
關鍵詞 荔枝;無核荔枝;單性結實;焦核荔枝
中圖分類號 S667.1 文獻標識碼 A
Abstract This paper reviews the research progress of seedless and stenospermocarpic mechanism in litchi(Litchi chinensis Sonn.). It points out that the abnormal development of megaspores(or embryo sacs), the failure of egg fertilization and parthenocarpy are the primary cause contributing to seedless litchi formation, therefore from now on, the study on the seedless mechanism of litchi should focus on the molecular one of the megaspores abortion and parthenocarpy. The most important thing is to clarify their molecular pathway. The stenospermocarpic mechanism of litchi was investigated by scientists from three aspects. At the cellular level, it is endosperms disintegrating ahead of time and embryos growing out of embryo sac that cause zygotes or embryos to abort from the early time of embryonic development, further resulting in abortion of ovules and seeds. From physiological and biochemical angle, the changes of plant endogenous hormones, polyamines, phenolic inhibitors and soluble protein content are closely related to abortion of ovules and formation of stenospermocarpy. At the molecular level, two specific amplified bands OPL-12-1 645 bp and OPL-12-722 bp have been considered as markers of seed-abortion genes in litchi. This paper suggests that in the future the study on the aborted-seeded mechanism of litchi should focus on zygote till heart-shape embryo stages so that it will be elucidated that which genesexpressing abnormally gives rise to zygote division blocked, endosperm disintegrating very early, ball-and heart-shape embryos formation blocked.
Key words Litchi(Litchi chinensis Sonn.);Seedless litchi;Parthenocarpy;Stenospermocarpic(aborted-seeded)litchi
doi 10.3969/j.issn.1000-2561.2016.05.030
荔枝(Litchi chinensis Sonn.)為無患子科(Sapindaceac)荔枝屬(litchi)植物,原產中國南部,至今已有兩千多年的栽培史。在這漫長的歷史進程中,人們通過實生選種和芽變選種選育出來的荔枝品種和株系多達200個以上。其中,最受老百性青睞的荔枝良種當屬無核荔枝和焦核荔枝。所謂無核荔枝是指果實內完全沒有核或僅有一個綠豆大小的痕跡,而焦核荔枝指的是果實內的核大小如蠶豆,但是不飽滿,呈扁平空癟狀態。無核荔枝和焦核荔枝是怎么形成的?兩者之間有何區別?關于這兩個問題科技界從上個世紀八十年代末九十年代初開始研究。三十多年過去了,荔枝無核和焦核機理的研究狀況究竟怎樣呢?現將有關的研究情況分述如下。
1 無核荔枝
1.1 已發現的無核荔枝的株系
經資料查新,沒有發現國外相關的報道。國內研究結果[1-11]表明:目前選育出來的無核荔枝株系至少有10個:分別是海南無核荔枝A4號、13號、南島無核丁香、廣東東莞無核荔枝、廣東增城假掛綠、廣東茂名茂紅糯、假香荔和禾蝦串,以及廣西欽州金科無核王和無核香荔。
1.2 無核荔枝形成機理的研究概況
總結科技界多年的研究成果,所有的證據都證實導致荔枝無核的原因是雌花中的大孢子發育異常無法完成受精,單性結實。陳健輝[1]、劉頌頌等[2]曾經以廣東東莞無核荔枝為材料、馮蔚等[7-8]以南島無核丁香為材料研究大孢子(也就是胚囊)的發育過程,發現大孢子在各個發育階段(二核胚囊期、四核胚囊期和八核胚囊期)均會出現各種敗育現象。由于胚囊內無法形成正常的卵器,卵細胞不能受精,胚無法形成,導致胚珠極早敗育所以無核。楊應華[9]以海南溫敏無核荔枝為材料研究其受精過程,發現溫敏無核荔枝由于受低溫影響,在受精過程中,精子不能進入卵膜或進入卵后雌雄核不融合,導致卵受精失敗并退化。而次生核雖正常受精,并產生核型胚乳,有的還能發育到細胞型胚乳,但到花后12 d左右也完全退化,結果導致胚珠敗育果實無核。王碧青等[10]以禾蝦串荔枝為材料研究單性結實荔枝的胚胎發育過程,發現單性結實的禾蝦串,開花后沒有雙受精過程發生及胚的發育,花后卵細胞和助細胞逐漸解體,極核核膜溶解核仁消失,胚囊內液體于花后3 d逐漸干涸;花后7 d胚珠開始漸漸萎縮,種子不發育;盡管如此,假種皮繼續發育形成無核果。以上是細胞學方面的證據,證明單性結實使荔枝結無核果。還有田間實驗證據。筆者曾經以海南無核荔枝A4號為材料做過研究,發現無核荔枝A4號在雌花未開放的時候,去雄去柱頭、套袋不授粉的情況下能結無核果,天生具有單性結實的能力;人工授粉結的也是無核果,而花粉并無敗育現象(圖1是無核荔枝A4號去雄去柱頭、套袋不授粉的結果情況)。劉頌頌等[2]也以廣東東莞無核荔枝為材料做過不授粉套袋實驗,發現該種質可以不經過授粉受精單性結實。目前所發現的10個無核荔枝株系幾乎都具有單性結實的能力。
那么究竟是什么因素使荔枝單性結實呢?從生理角度分析,可能是激素。王碧青等[11]曾經以懷枝、禾蝦串兩種荔枝品種為材料研究了荔枝開花前后子房內源激素含量變化,發現具單性結實能力的禾蝦串開花前子房中細胞分裂素(CTK)、赤霉素(GAs)含量較高,開花后一直處于較高水平;吲哚乙酸(IAA)開花前子房含量較低,但開花后上升, 第3天有一高峰; 而不具單性結果能力的懷枝(大核品種)開花前子房CTK、GAs含量低于禾蝦串。說明單性結實與荔枝開花前后子房內高水平的生長促進類激素有關。另外,邱燕萍等[12]以糯米糍、淮枝、禾蝦串3種荔枝品種為材料研究了荔枝花期內源激素平衡與單性結實的關系,發現花后3 d禾蝦串無核果生長促進類激素達到最高值,(IAA+GAs+CTK)/ABA(脫落酸)>=14.8,而大核品種淮枝的(IAA+GAs+CTK)/ABA<=1。由此可以看到,開花期相對高水平的生長促進類激素使禾蝦串單性結實子房順利進入坐果期。李偉才等[13]以海南3種無核荔枝(A4號、13號、南島無核丁香)和大核荔枝紫娘喜為材料研究了謝花后果實發育過程中內源激素的變化趨勢,發現謝花后14~21 d,3種無核荔枝果實中IAA的含量是紫娘喜的3倍;35 d時,紫娘喜果實中的IAA含量達最高值,為23 ng/g FW,IAA水平逐漸緩慢降低;而49 d時,3種無核荔枝果實中IAA含量均達到高峰值70.3 ng/g FW,約是紫娘喜最高值的3倍左右。可見,果實發育中后期高濃度的IAA是導致無核荔枝具備單性結實能力的主要原因,是果實無種胚但能繼續發育的源泉。
王碧青等[11]曾經用植物生長調節劑2,4-二氯苯氧乙酸(2,4-D)處理妃子笑和糯米糍品種未授粉的子房,結果在妃子笑上成功地獲得誘導性單性結實無核果;李再峰等[14]應用雜環脲類細胞分裂素使無單性結實能力的桂味、廣良紅和妃子笑3個品種產生了部分單性結實無核果,從而進一步證實激素能導致荔枝單性結實結無核果。
1.3 無核荔枝形成機理未來的研究重點和研究方向
由于大孢子發育異常導致卵細胞無法完成受精和單性結實是導致荔枝無核的主要原因,因此今后對無核荔枝無核機理的研究應該著重從這兩個方面展開。重點放在胚囊發育期到果實發育早期,因為這個時期恰好是荔枝形成無核的關鍵期,大量的分子事件和細胞水平的變化發生在這個時期。有兩個問題必須闡明:究竟是什么基因突變、表達使大孢子敗育、受精無法完成?什么基因突變、表達使果實在沒有受精的前提下得以發育?本課題組研究者推測:可能是與生長素、細胞分裂素、赤霉素等植物激素生物合成途徑、信號傳導途徑相關的基因發生了突變,導致開花前后子房內這些激素含量大幅提高,從而引起胚囊在發育過程中敗育、受精無法完成,種子無法形成,最后結無核果,行單性結實。當然這只是一種猜測,還需要通過實驗加以證實,最終目的是闡明無核荔枝發生大孢子敗育、受精被阻和單性結實的分子途徑。
2 焦核荔枝
與無核荔枝相比,焦核荔枝胚囊內的卵細胞完成了受精過程,但在胚胎發育過程中因受某種因素影響而中途敗育,形成的果實內種子呈扁平空癟狀態,故而稱之為焦核荔枝。
2.1 焦核荔枝的種類
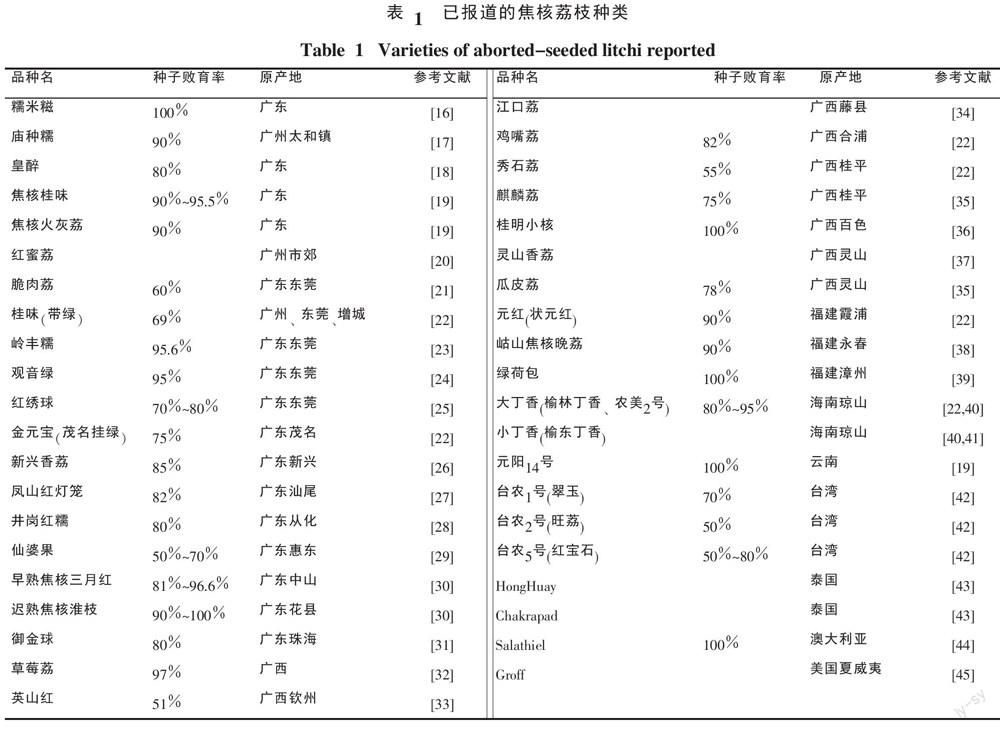
目前選育出來的焦核荔枝種類很多,國內外都有報道,表1列出已見報道的焦核荔枝的主要種類及其產地。
2.2 焦核荔枝形成機理的研究概況
為了解析焦核荔枝的形成機制,科技界主要從3個方面對其進行了研究。
2.2.1 組織細胞水平的研究 通過對大核荔枝和焦核荔枝花后胚胎發育的顯微觀察發現焦核荔枝有3種情況:一種像糯米糍、桂味等品種,其卵細胞受精形成合子以后,部分合子不分裂,解體,沒有形成胚[46];第二種像綠荷包,合子形成后,一部分合子分裂幾次就停止發育并解體,另一部分則發育到球形原胚至球形胚時停止發育[47-48];第三種像蘭竹,其部分果實的胚發育到球形胚、心形胚時停止發育。可能由于合子或胚發生敗育,導致胚珠敗育、成熟種子敗育,最后形成焦核。
總結前人的研究結果,從細胞水平看,那么是什么原因引起荔枝胚珠中合子或胚敗育呢?可能有以下兩個原因:一是胚乳過早解體。荔枝受精后,初生胚乳核立即分裂形成多個游離核,花后20~30 d,游離核數量達到最多;花后50 d,全部解體被胚吸收,這是大核荔枝品種胚乳的解體時間。而焦核荔枝品種胚乳解體時間比較早,如糯米糍的胚乳核達26個左右即停止發育,逐漸解體;綠荷包在球形胚形成之前,大部分胚乳已干枯解體[49]。胚乳過早解體,使合子或胚發育失去營養源,因而停止了發育[50]。二是胚在胚囊外發育。張健等發現綠荷包的胚在發育過程中,被胚柄細胞推出了珠孔,使之游離在內、外珠被之間的空腔里。這種胚在胚囊外發育的畸變現象使胚不能獲得發育所需的正常養分因而敗育[48]。
2.2.2 生理生化角度的研究 主要以大核荔枝和焦核荔枝的幼果、胚珠、幼胚、胚乳為材料測定內源生長調節物即內源植物激素、多胺、酚類化合物和蛋白質的含量,以研究上述生理指標的差異是否與胚珠敗育、焦核形成有關。
(1)內源植物激素與胚珠敗育、焦核形成的關系。1990年,葉明志等[47,51]率先以大核荔枝品種烏葉和焦核荔枝品種綠荷包的幼果為材料測定其內源生長調節物的含量變化,結果發現在促進物含量方面,綠荷包在合子期、球形胚期、魚雷胚至子葉胚期都低于烏葉;而抑制物的含量在合子期、心形胚期、魚雷胚至子葉胚期均高于烏葉。從這個結果可以看到內源生長調節物的水平與合子、幼胚及胚珠敗育都有關系。由于是用幼果作試驗材料,且生長調節物的含量測定采用的是小麥芽鞘垂直生長法,也許是擔心這種生物測定法的實驗結果有誤差,2000年,陳偉等[52]又以烏葉和綠荷包的胚珠為材料采用酶聯免疫吸附法(ELISA)測定了IAA、GA1+3、CTK和ABA的含量變化,結果發現綠荷包胚珠中ABA的含量在所測定的6個時期都高于烏葉,而IAA、GA1+3、CTK的含量在所有時期均低于烏葉,與葉明志的研究結果相似。與此同時,陳偉[53]還以部分焦核型品種蘭竹荔枝的正常發育胚珠和敗育胚珠為材料, 研究了胚胎發育與內源激素的關系,得到的結果也類似:在胚胎敗育期,敗育胚珠中IAA 和GA1+3的含量都降至最低點,CTK的含量明顯低于正常胚珠;而ABA的含量則明顯高于正常胚珠。另外發現敗育胚珠中ABA/IAA+GA1+3與ABA/CTK兩個比值顯著高于正常胚珠[52-53]。以上結果說明在胚胎發育進程中高的生長抑制物質與低的生長促進物質使植物內源激素的平衡失調,進而使合子、幼胚及胚珠敗育,形成焦核。
不過,張以順等[54]以另外兩個荔枝品種——焦核荔枝品種糯米糍與大核荔枝品種懷枝的幼胚及胚乳為材料測定內源激素的含量變化,得到的結果略有不同。發現糯米糍幼胚及胚乳中的IAA、GAs含量低于懷枝,CTK含量卻顯著高于懷枝;在花后10 d的時候,ABA含量低于懷枝,而隨著胚胎的發育進程,糯米糍幼胚及胚乳中的ABA含量均表現為上升趨勢,尤其是在幼胚內表現最為顯著,升幅高達4.5倍;而同期懷枝幼胚內的ABA含量呈下降趨勢。由此認為,焦核荔枝品種幼胚中的ABA急劇上升改變了生長促進物質與生長抑制物質的正常配比,是導致其胚敗育的一個重要原因。
(2)多胺與胚珠敗育、焦核形成的關系。陳偉等[55]曾經以烏葉、綠荷包以及蘭竹的正常胚珠與敗育胚珠為材料測定了3種多胺含量的變化,結果發現正常發育的胚珠中腐胺(Put)、亞精胺(Spd)和精胺(Spm)的含量在極核分裂、合子分裂、球形胚、心形胚、魚雷胚和子葉胚發育階段均高于敗育胚珠;并且綠荷包敗育胚珠的多胺含量從合子期開始急劇下降,蘭竹敗育胚珠的多胺含量從球形胚期開始急劇下降,其下降速度顯著大于正常胚珠。這與形態學上觀察到的綠荷包胚珠從合子期開始敗育, 蘭竹焦核荔枝的胚珠從球形胚期開始敗育,而烏葉胚珠正常發育的結果是一致的。說明低的多胺水平也會導致合子、幼胚及胚珠敗育,從而形成焦核。
(3)酚類物質與胚珠敗育、焦核形成的關系。 陳偉等[56]研究發現綠荷包和蘭竹的敗育胚珠中有一種酚類抑制物質含量很高,與烏葉、蘭竹的正常胚珠相比,其差異達極顯著水平,經鑒定這種物質是對羥基苯甲酸(p-HBA);同時發現敗育胚珠中IAA氧化酶活性也顯著高于正常胚珠, 而IAA水平則明顯低于正常胚珠。因此認為,高含量的p-HBA是通過促進IAA側鏈的氧化進而影響生長促進物質和抑制物質的平衡最終使荔枝胚珠發生敗育、形成焦核的。
也許是所用的試驗材料不一樣,張以順等[54]的研究結果不盡相同。他們發現:焦核品種桂味和糯米糍幼胚中的多酚類物質含量略低于大核品種黑葉和懷枝,而胚乳中的多酚類物質含量顯著高于大核品種。這樣一來桂味和糯米糍的胚乳過早敗育、解體,幼胚因為得不到胚乳提供的養分因而發育受到抑制。
(4)蛋白質與胚珠敗育、焦核形成的關系。 張以順等[54]研究發現:焦核荔枝品種桂味和糯米糍的胚及胚乳中可溶性蛋白質含量顯著低于大核品種黑葉和懷枝,說明焦核荔枝品種的幼胚和胚乳中蛋白質的合成和積累受到了明顯抑制。由此推測焦核荔枝品種的幼胚和胚乳中很多基因的表達都受到了抑制,而這顯然與其胚珠敗育、焦核形成有關。
2.2.3 分子水平的研究 2002年,劉成明等[57]對荔枝品種三月紅、懷枝和它們的9個自然焦核突變體進行隨機擴增多態性DNA標記(RAPD)分析,經500個隨機引物的篩選, 發現僅1個引物OPL-12 能在原品種與突變體間擴增出多態性片段, 其中2個焦核懷枝的特異片段大小為1 645 bp, 而另外4個焦核三月紅的特異片段大小為722 bp, 初步認為這兩條特異片段可能與荔枝的焦核基因相關。但同時發現另外三個焦核突變體并沒有出現這兩條特異帶,推測可能還存在著其它控制荔枝焦核性狀的基因。
2004年,張以順[58-59]以‘桂味荔枝的正常發育胚珠和敗育胚珠為材料采用抑制消減雜交技術分離到3個在敗育胚珠中特異表達的基因片段,經Blast同源性比對,發現其中一個基因片段與泛素結合酶基因同源性較高,另外兩個基因片段與S-腺苷甲硫氨酸合成酶基因同源性較高。經分析,S -腺苷甲硫氨酸合成酶基因編碼S-腺苷甲硫氨酸合成酶,以催化甲硫氨酸和三磷酸腺苷(ATP)反應生成S-腺苷甲硫氨酸(SAM)。在生物學上,SAM是多胺和乙烯合成的前體, 而多胺和乙烯的生物合成與植物胚胎的發育密切相關, 因此推測, S-腺苷甲硫氨酸合成酶基因在荔枝敗育胚珠中大量表達可能參與了其體內乙烯合成的調控, 從而導致荔枝胚胎敗育。當然這只是推測,還沒有通過功能鑒定證明這一點。而泛素結合酶基因是否與植物的胚胎敗育有關,同樣也缺乏實驗證據。
2.3 焦核荔枝的誘導
由于內源植物激素能引起荔枝合子或胚早期敗育,促使人們想到用外源植物生長調節劑來處理大核荔枝品種以誘導焦核荔枝,這方面已有一些成功的報道。最早是1984年,邱似德等[60]應用1.0~1.5 g/L的青鮮素(MH)于花后7-13 d 噴果成功獲得淮枝品種的焦核果,焦核率達73%。1998年,刁森標等[61]報道用濃度為1 000~2 000 μg/mL的青鮮素處理淮枝,焦核率達到97.6%~100%。2002年,向旭等[60]報道用MH(0.8 g/L)+β-萘氧乙酸(BNOA, 30 mg/L)處理‘雪懷子,于雌花謝后15、25 d連續噴果2次,獲得焦核率達87.5%的優質果實。2013年,李冬波等[15]用MH(800 mg/L)+2,4-D(5 mg/L)處理大核品種‘黑葉和部分焦核品種‘靈山香荔,使靈山香荔的焦核率由50%提高到了89.77%,而黑葉的焦核誘導率只有8.79%,盡管如此,黑葉大核種子質量降低了20%以上。總之,經過多年研究證明,青鮮素能夠誘導荔枝胚早期敗育,從而形成焦核荔枝,但是它單獨使用、使用濃度過高、使用次數超過兩次,焦核率雖然提高了,但是果實偏小,沒有商業價值并且還會引起落果。而將青鮮素與其它植物生長調節劑搭配使用,青鮮素使用的濃度、次數低一點,能夠獲得具市場競爭力的焦核荔枝果。當然,荔枝品種不同,其焦核率的誘導效果也會不同。
2.4 焦核荔枝焦核機理未來的研究重點和研究方向
從上面的敘述可以看到,過去對荔枝焦核機理的研究多集中于組織細胞水平和生理生化角度,分子水平的研究不多。由于合子或胚早期敗育是導致荔枝形成焦核的主要原因,因此今后對焦核荔枝焦核機理的研究應該著重于合子期到心形胚期,包括胚乳核分裂期、合子分裂期、球形胚期和心形胚期,焦核荔枝敗育就發生在這幾個時期。可以通過激光顯微切割技術分離大核荔枝品種正常發育胚珠和焦核荔枝品種敗育胚珠中的合子、胚乳核、球形原胚和球形胚,然后通過單細胞轉錄組測序結合熒光定量PCR獲得正常分裂的合子與不分裂的合子之間、正常分裂的胚乳核與不分裂的胚乳核之間、正常分裂的球形原胚與不分裂的球形原胚之間、正常分裂的球形胚與不分裂的球形胚之間差異表達的基因,以便從分子水平上闡明究竟是哪些基因突變或異常表達使合子分裂受阻、使胚乳過早解體,使球形胚、心形胚形成受阻。另外,對于部分焦核型荔枝品種的形成機理也要加大研究力度,為什么同一棵荔枝樹上只有一部分果實形成焦核?究竟是溫度等環境因素引起的還是基因突變引起的?對上述問題的研究將從根本上解決荔枝生產實踐上普遍存在的焦核荔枝焦核率不穩定的問題,從而提高焦核荔枝的商品價值。
參考文獻
[1] 陳健輝, 楊俊慧, 潘坤清, 等. 無核荔枝胚胎異常發育的研究[J]. 植物學通報, 1998(15)(增刊): 95-100.
[2] 劉頌頌, 葉永昌, 招曉東,等. 無核荔枝種子敗育的胚胎學研究[J]. 華南農業大學學報, 1999, 20(2): 41-46.
[3] 陳健輝,楊俊慧,潘坤清. 無核荔枝假種皮發育過程的研究[J]. 廣西植物, 2001, 21(4): 362-366.
[4] 張展薇, 邱燕萍, 向 旭. 荔枝單性結實研究初報[J]. 果樹科學, 1990, 7(4): 234-235.
[5] 張承運,張景照. 介紹幾種無核荔枝良種[N]. 福建科技報,2002-09-03.
[6]王廷標. 無核荔枝新株系的發現及溫敏無核理論和無核技術研究[J]. 熱帶作物科技, 1997(5): 1-7.
[7]馮 蔚. 海南無核荔枝胚胎發育及敗育的機理初探[D]. 海口: 海南大學碩士學位論文, 2010.
[8] 馮 蔚, 張 蕾, 李松剛, 等. 3個荔枝品種胚胎發育過程的比較[J]. 熱帶作物學報, 2010, 31(5): 736-739.
[9] 楊應華, 李 蕾,余誕年. 溫敏無核荔枝的種子敗育和果實發育研究[J]. 海南師范學院學報(自然科學版), 2002, 15(2):71-74.
[10] 王碧青, 邱燕萍,張展薇. 單性結實荔枝胚胎及果實發育觀察[J]. 廣東農業科學, 1996(2): 15-17.
[11] 王碧青, 邱燕萍, 向 旭,等. 荔枝結果過程中內源激素變化及單性結果的誘導[J]. 園藝學報, 1997, 24(1): 19-24.
[12] 邱燕萍, 向 旭,王碧青, 等. 荔枝三種結實類型內源激素的平衡與坐果機理[J]. 果樹科學, 1998, 15(1): 39-4 3.
[13] 李偉才, 魏永贊, 胡會剛, 等. 3種無核荔枝果實發育過程中內源激素含量變化動態[J]. 熱帶作物學報, 2011, 32(6): 1 042-1 045.
[14] 李再峰, 歐陽樂軍,李莉梅. 雜環脲類細胞分裂素誘導荔枝單性結實的試驗研究[J]. 湛江師范學院學報, 2014, 35(6): 79-84.
[15] 李冬波, 黎曉峰,黃鳳珠. 荔枝胚胎發育不同品種誘導焦核的研究[J]. 中國農學通報, 2013, 29(16): 115-118.
[16] 鄧九生, 黃在猛. 龍眼荔枝的果實發育與調控[J]. 廣西熱作科技, 1998(2): 1-5.
[17] 劉成明, 胡桂兵, 黃穗生, 等. 晚熟優質荔枝新品種‘廟種糯[J]. 園藝學報, 2014, 41(3): 595-596.
[18] 陳潔珍, 歐良喜, 王麗敏, 等. 荔枝釀酒優良單株“皇醉”和“妃醉”初選[J]. 南方農業學報, 2013, 44(1): 121-125.
[19] 劉成明, 胡又厘, 傅嘉欣, 等. 荔枝育種研究進展——技術、成就及努力方向[J]. 中國南方果樹, 2004, 33(2): 36-40.
[20] 馬 錁, 胡銳清, 尹金華, 等. “紅蜜荔”荔枝的生長特性及栽培技術要點[J]. 中國南方果樹, 2011, 40(5): 89-90.
[21] 葉耀雄, 朱劍云, 葉永昌, 等. 4個優質荔枝品種生長規律及果實性狀比較分析[J]. 中國園藝文摘, 2010(11): 22-23, 25.
[22]林旗華,楊 凌. 中國晚熟荔枝品種介紹[J]. 現代園藝, 2010(11): 16-17.
[23] 范 妍, 尹金華, 劉成明,等. 晚熟荔枝新品種——嶺豐糯的選育[J]. 果樹學報, 2010, 27(5): 852-853.
[24] 王澤槐, 劉秀榮, 陳襯喜, 等. 荔枝新品種‘觀音綠[J]. 園藝學報, 2012, 39(8): 1 615-1 616.
[25] 邱燕萍, 歐良喜, 李志強,等. 荔枝新品種紅繡球的選育[J]. 中國果樹, 2008(5): 8-9.
[26] 陳贊盛, 趙 軍, 羅秀英. 增大荔枝果實并提高座果率研究[J]. 仲愷農業技術學院學報, 1992, 5(1): 64-68.
[27] 歐良喜, 陳潔珍,蔡長河等. 優質荔枝新品種——鳳山紅燈籠的選育[J]. 果樹學報, 2012, 29(1): 314-315.
[28] 葉自行, 胡桂兵, 陳杰忠,等. 晚熟抗裂果荔枝新品種‘井崗紅糯[J]. 園藝學報, 2011, 38(1): 189-190.
[29] 潘建平, 林志雄, 古河洋, 等. 豐產、優質荔枝仙婆果的選育研究[J]. 廣州大學學報(自然科學版), 2007, 6(2): 26-29.
[30] 王澤槐, 梁立峰. 廣東荔枝10年研究進展概述[J]. 廣東農業科學, 1994(4): 22-24.
[31] 孫清明, 李永忠, 向 旭, 等. 利用SNP和EST-SSR分子標記鑒定荔枝新種質御金球[J]. 分子植物育種, 2013, 11(3): 403-414.
[32] 彭宏祥, 朱建華, 劉業強, 等. 焦核優質荔枝新品種‘草莓荔[J]. 園藝學報, 2007, 34(6): 1 592.
[33] 錢開勝. 荔枝實生變異新品種“英山紅”通過審定[J]. 中國果業信息, 2010, 27(11): 43.
[34] 黎展文, 林起森, 戴國權,等. 晚熟荔枝品種——“江口荔”的選育和栽培特性[J]. 中國南方果樹, 2010, 39(1): 51-52.
[35] 朱建華, 彭宏祥, 蘇偉強, 等. 廣西八個荔枝新品種(株系)果實品質分析[J]. 中國南方果樹, 2005, 34(6): 31-32.
[36] 梁逸飛. 廣西荔枝[J]. 廣西農業科學, 1982(2): 27-30.
[37] 朱建華, 彭宏祥, 潘介春,等. 廣西荔枝種質資源研究及品種選育[J]. 福建果樹, 2005(134): 11-13.
[38] 尤有利, 王江波, 潘東明,等. 晚熟焦核優質荔枝新株系——岵山焦核晚荔[J]. 中國南方果樹, 2012, 41(2): 99-100.
[39]譚全偉. cDNA-AFLP技術分離和分析荔枝胚胎發育相關基因[D]. 福州: 福建農林大學碩士學位論文, 2010.
[40] 陳業淵, 高愛平. 海南適栽荔枝優良品種選擇初探[J]. 熱帶作物研究, 1998(2): 39-43.
[41] 王家保, 鄧穗生,劉志媛,等. 海南荔枝(Litchi chinensis Sonn.)主要栽培品種的RAPD分析[J]. 農業生物技術學報,2006, 14(3): 391-396.
[42]向 旭, 顏昌瑞, 柯立祥, 等. 臺灣省荔枝育種概況與新品種選育進展[J]. 現代農業科技, 2012(7): 141-143.
[43] 徐遲默. 世界荔枝品種[J]. 世界熱帶農業信息, 2007(5): 20-23.
[44] 黃旭明, 胡桂兵, 陳維信, 等. 澳大利亞荔枝產業考察報告[J]. 中國園藝文摘, 2009(5): 1-5.
[45] 孫清明, 歐良喜, 向 旭, 等. 荔枝品種選育進展[J]. 果樹學報, 2010, 27(5): 790-796.
[46] 葉秀粦, 王伏雄, 錢南芬. 荔枝的胚胎學研究[J]. 云南植物研究, 1992, 14(1): 59-65.
[47] 呂柳新, 余小玲, 葉明志,等. 荔枝胚胎發育機制的探討[J]. 福建農學院學報, 1989, 18(2): 149-155.
[48] 張 健, 呂柳新, 葉明志. 胚胎敗育型荔枝胚胎發育異常的顯微及亞顯微觀察[J]. 河南農業大學學報, 2006, 40(2): 194-197.
[49] 張 健, 呂柳新, 葉明志. 荔枝胚胎發育研究進展[J]. 福建農業學報, 2001, 16(1): 57-60.
[50] 李明芳,鄭學勤. 荔枝開花結果調控機制及種質分析研究進展[J]. 熱帶農業科學, 2004, 24(2): 41-47.
[51] 葉明志, 肖振通, 呂柳新,等. 荔枝幼果內源生長調節物的消長與胚胎發育的關系[J]. 福建農學院學報, 1990, 19(3): 268-272.
[52] 陳 偉, 呂柳新, 葉陳亮,等. 荔枝胚胎敗育與胚珠內源激素關系的研究[J]. 熱帶作物學報, 2000, 21(3): 34-38.
[53] 陳 偉,呂柳新. 蘭竹荔枝胚珠內源激素含量變化與胚胎發育的關系[J]. 應用與環境生物學報, 2000, 6(5): 419-422.
[54] 張以順, 向 旭, 黃上志,等. 荔枝胚敗育過程中內源激素與蛋白質含量的變化[J]. 植物生理與分子生物學學報, 2003, 29(3): 233-238.
[55] 陳 偉,呂柳新. 荔枝胚珠中多胺含量變化與胚胎發育的關系[J]. 熱帶亞熱帶植物學報, 2000, 8(3): 229-234.
[56] CHEN Wei, L LiuXin. Relationship Between Litchi Embryo Abortion and Phenolic Inhibitors[J]. Acta Botanica Sinica,2002, 44(2): 168-172.
[57] 劉成明, 梅曼彤. 利用RAPD分析鑒別荔枝的焦核突變體[J]. 園藝學報, 2002, 29(1): 57-59.
[58] 張以順, 向 旭, 傅家瑞,等. 荔枝胚敗育差異表達基因 cDNA片段的克隆及序列分析[J]. 園藝學報, 2004, 31(1): 25-28.
[59] 張以順, 向 旭, 傅家瑞,等. 荔枝敗育胚S-腺苷甲硫氨酸合成酶基因的全長擴增和序列分析[J].園藝學報, 2004, 31(2):160-164.
[60] 向 旭, 歐良喜, 邱燕萍,等. 荔枝大核品種誘導焦核結果研究[J]. 果樹學報, 2002, 19(2): 111-114.
[61] 刁森標,官雪芳. 赤霉素和青鮮素誘導荔枝果實無核或細核試驗[J]. 中國南方果樹, 1998, 27(4): 25.
