一株鐵還原菌的分離及其碳源利用特性研究
2016-06-17 09:43:07李湘凌吳紀(jì)南周濤發(fā)
童 磊,李湘凌,吳紀(jì)南,袁 峰,周濤發(fā)
(合肥工業(yè)大學(xué) 資源與環(huán)境工程學(xué)院,安徽 合肥 230009)
?
一株鐵還原菌的分離及其碳源利用特性研究
童磊,李湘凌,吳紀(jì)南,袁峰,周濤發(fā)
(合肥工業(yè)大學(xué) 資源與環(huán)境工程學(xué)院,安徽 合肥230009)
摘要:文章探究從污水處理廠(chǎng)污泥中篩選的一株鐵還原菌的生長(zhǎng)特性,研究不同碳源對(duì)Fe(Ⅲ)還原的影響。結(jié)果表明:此株鐵還原菌株適宜的生長(zhǎng)條件為pH=7、溫度為35℃、黑暗條件;碳源類(lèi)型和濃度顯著影響菌株的鐵還原能力,其最適碳源濃度為1.5 mol/L,以蔗糖、葡萄糖、丙酮酸鈉、乙酸鈉、乳酸鈉為碳源時(shí),其Fe(Ⅲ)還原率依次降低,以蔗糖和葡萄糖為碳源時(shí)Fe(Ⅲ)還原率分別為81.0%和57.2%;Fe(Ⅲ)還原過(guò)程中脫氫酶活性與Fe(Ⅲ)還原率顯著正相關(guān),脫氫酶活性能在一定程度上反映Fe(Ⅲ)還原程度。
關(guān)鍵詞:鐵還原菌;碳源;脫氫酶活性
異化鐵還原是鐵還原菌介導(dǎo)進(jìn)行的Fe(Ⅲ)還原,該過(guò)程廣泛地存在于自然界的厭氧環(huán)境[1]。在異化鐵還原過(guò)程中鐵還原菌不僅以簡(jiǎn)單有機(jī)物等為電子供體,而且可以偶聯(lián)難降解有機(jī)污染物氧化分解及重金屬還原等,因而鐵異化還原對(duì)環(huán)境污染修復(fù)具有重要的意義[2-3]。在異化鐵還原相關(guān)研究中,高效鐵還原菌的分離純化工作是研究異化鐵還原過(guò)程的基礎(chǔ),也是利用鐵還原菌開(kāi)展污染修復(fù)的基礎(chǔ)。迄今,研究人員從各種厭氧環(huán)境中分離得到了不同的異化Fe(Ⅲ)還原菌,這些Fe(Ⅲ)還原菌可利用的電子供體包括了纖維素、糖類(lèi)、氨基酸、氫氣和有機(jī)酸等[3],如其中研究最為廣泛和系統(tǒng)的Geobacter和Shewanella分別以乙酸鹽和乳酸鹽、丙酮酸鹽為電子供體[4-5]。而微生物對(duì)碳源要求的單一性會(huì)極大地限制微生物在環(huán)境污染修復(fù)中應(yīng)用。因此,研究并探明不同鐵還原菌對(duì)不同碳源的利用特征,得到可利用多種碳源的鐵還原菌對(duì)其在污染修復(fù)中應(yīng)用具有重要意義。
本文從污水處理廠(chǎng)的污泥中分離純化鐵還原菌,以蔗糖、葡萄糖、丙酮酸鈉、乳酸鈉和乙酸鈉分別作為碳源(電子供體),研究該鐵還原菌厭氧條件下Fe(Ⅲ)的還原能力,并探索不同碳源條件下脫氫酶在Fe(Ⅲ)還原過(guò)程中的意義。
1材料與方法
1.1儀器
采用722E型分光光度計(jì)、無(wú)菌操作臺(tái)、HC-2064高速離心機(jī)。
1.2培養(yǎng)基
50% LB液體培養(yǎng)基:蛋白胨5 g/L,酵母膏5 g/L,NaCl 2.5 g/L,調(diào)節(jié)培養(yǎng)基pH=7。
分離純化培養(yǎng)基:C6H12O6·H2O 1.0 g/L,FeCl3·6H2O 9.6 g/L,NH4Cl 0.53 g/L,KH2PO40.272 g/L,CaCl20.056 g/L,NaHCO30.252 g/L(使用量根據(jù)富集期間pH=7±0.2而定),微量元素溶液1 mL。
鐵還原培養(yǎng)基:檸檬酸鐵3.3 g/L,NH4Cl 1 g/L,CaCl2·2H2O 0.07 g/L,MgSO4·7H2O 0.6 g/L,K2HPO4·3H2O 0.722 g/L,KH2PO40.25 g/L,葡萄糖10 g/L,調(diào)節(jié)培養(yǎng)基pH=7。將該培養(yǎng)基在0.1 MPa下滅菌30 min,加瓊脂粉10 g/L即為固體培養(yǎng)基。
1.3菌種來(lái)源及培養(yǎng)
菌種來(lái)源于合肥市王小郢污水處理廠(chǎng)的消化池污泥。
富集培養(yǎng):將20 mL污泥和180 mL分離純化培養(yǎng)基加入250 mL鹽水瓶,氮?dú)怛?qū)氧10 min后密封,30℃恒溫暗條件下培養(yǎng)。待溶液顏色由棕黃色變成無(wú)色時(shí),取鹽水瓶中菌懸液繼續(xù)富集培養(yǎng)。連續(xù)富集培養(yǎng)3次后,從鹽水瓶中吸取2 mL富集培養(yǎng)液加入25 mL血清瓶中,同時(shí)添加18 mL檸檬酸鐵培養(yǎng)基,氮?dú)怛?qū)氧10 min后密封,于30℃恒溫培養(yǎng)箱中培養(yǎng),每隔一天搖勻,當(dāng)菌液由黃色變成無(wú)色后,再連續(xù)富集3次。
分離純化:取上述最終富集所得菌液在檸檬酸鐵固體培養(yǎng)基上進(jìn)行平板畫(huà)線(xiàn)和涂布,待平板中長(zhǎng)出細(xì)菌后,挑取形態(tài)一致、生長(zhǎng)快的菌落接種到液體培養(yǎng)基中培養(yǎng);將培養(yǎng)所得菌液劃線(xiàn)和涂布,進(jìn)一步分離純化菌種;重復(fù)該工作5次后,根據(jù)菌落形態(tài)和細(xì)胞的一致性判斷菌株的純化狀態(tài),選取最優(yōu)菌株。制得菌液,轉(zhuǎn)入1.5mL離心管中,4 000 r/min離心10 min,棄上清液,分離得到的菌液于4℃保存?zhèn)溆谩?/p>
1.4實(shí)驗(yàn)設(shè)計(jì)
1.4.1菌株基本理化性質(zhì)分析
觀(guān)察菌株在檸檬酸鐵固體培養(yǎng)基上的菌落特征,并進(jìn)行菌株革蘭氏染色試驗(yàn)[6]。
1.4.2菌株生長(zhǎng)特性
(1)pH對(duì)菌株生長(zhǎng)的影響。將50% LB液體培養(yǎng)基的pH值用0.1 mol/L 的NaOH和HCl調(diào)至4、5、6、7、8、9,每個(gè)梯度設(shè)置3個(gè)水平,滅菌后接入2 mL菌懸液,置于30℃培養(yǎng)箱中避光培養(yǎng)。然后分別在第2、4、6、8、10、12、16、20、26、32、40 h用注射器在錐形瓶中取出2 mL,置于5 mL離心管中測(cè)定菌液OD值。
(2)溫度對(duì)菌株生長(zhǎng)的影響。將配置好的培養(yǎng)基pH值調(diào)節(jié)為7,滅菌后接入2 mL菌懸液,置于溫度分別為4、15、20、30、35、40℃的培養(yǎng)箱中避光培養(yǎng),每種溫度下設(shè)置3個(gè)水平。然后按上述(1)的操作測(cè)定OD值。
(3)光照對(duì)菌株生長(zhǎng)的影響。將配置好的培養(yǎng)基pH調(diào)節(jié)為7。滅菌后接入2 mL菌懸液,置于溫度為35℃的培養(yǎng)箱中,分別采取連續(xù)光照和避光暗處理,每種處理設(shè)置3個(gè)水平。然后按上述(1)的操作測(cè)定OD值。
1.4.3不同碳源條件下的Fe(Ⅲ)還原
將純化菌液擴(kuò)繁32 h后,測(cè)定菌液濃度后備用。選用碳源為蔗糖(Sue)、葡萄糖(Glu)、丙酮酸鈉(Pyr)、乙酸鈉(Ace)和乳酸鈉(Lac),以單一變量的控制方式分別添加,見(jiàn)表1所列。
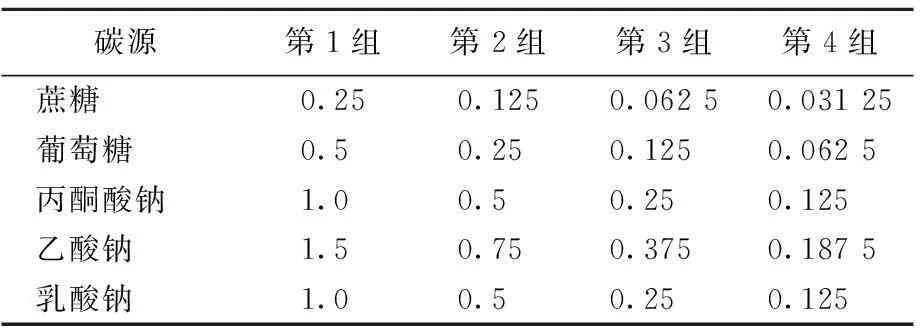
表1 碳源添加量 mol/L
注:第1組的碳濃度為3.0 mol/L,第2~4組依次減1/2。
采用厭氧混合培養(yǎng)。分別取2 mL碳源、2 mL NH4Cl(5 g/L)和2 mL的檸檬酸鐵溶液(13 g/L)于10 mL的血清瓶中,高壓滅菌后,接種液2 mL擴(kuò)繁菌液,并用滅菌后的磷酸緩沖溶液調(diào)節(jié)pH=7±0.2,氮?dú)怛?qū)氧5 min,用膠塞和鋁蓋密封[7]后置于35℃恒溫箱中培養(yǎng)。每組實(shí)驗(yàn)中不添加碳源為平行樣。
1.5分析方法
(1)菌懸液濃度。比濁法測(cè)定,由于細(xì)菌懸液的濃度與光密度(OD值)成正比,因此可利用分光光度計(jì)測(cè)定菌懸液的光密度來(lái)推知菌液的濃度,選用600 nm波長(zhǎng)分光光度計(jì),以同樣培養(yǎng)基作為空白對(duì)照,并對(duì)培養(yǎng)液從0 h起依次進(jìn)行測(cè)定,對(duì)濃度大的菌懸液用未接種的液體培養(yǎng)基適當(dāng)稀釋后測(cè)定,使其OD值在0.10~0.65以?xún)?nèi)。
(2)Fe(Ⅱ)測(cè)定。采用鄰菲啰啉比色法,在培養(yǎng)的1個(gè)周期內(nèi)每天于同一時(shí)刻進(jìn)行采樣,每次采樣時(shí)取待測(cè)樣品1瓶,搖勻,用注射器平行取樣品液2.0 mL 置于含2 mL的1+3 HCl(HCl與H2O的體積比為1∶3)的50 mL比色皿中,重復(fù)3組,分別加入5 mL鐵緩沖溶液與鄰菲羅啉溶液2 mL,顯色5~10 min,在510 μm處以水為參比測(cè)量吸光度。
(3)脫氫酶活性測(cè)定。脫氫酶活性(dehydrogenase activity,DHA)測(cè)定采用TTC分光光度法[8],將吸取的1 mL樣品溶液置于含有2 mL Tris-HC緩沖液(pH值為8.4)、0.5 mL 3.6 g/L Na2SO3溶液和0.5 mL 20 g/L TTC溶液的離心管中,重復(fù)3組,置37℃恒溫水浴中避光反應(yīng)2 h后加入0.5 mL 甲醛終止酶反應(yīng)。再加5 mL丙酮作為萃取劑[9],37℃200 r/min振蕩萃取10 min,于3 500 r/min離心5 min,取上清液于492 nm下測(cè)定吸光度。通過(guò)標(biāo)準(zhǔn)曲線(xiàn)計(jì)算生成的TF含量,定義1 h產(chǎn)生1 mg/L為1個(gè)脫氫酶活力單位(U)[10]。
1.6動(dòng)力學(xué)模型
本文采用Logistic方程描述微生物介導(dǎo)的異化Fe(Ⅲ)還原過(guò)程[11],Logistic方程為:
其中,t為培養(yǎng)時(shí)間;ct表示培養(yǎng)時(shí)間t時(shí)體系中Fe(Ⅱ)的濃度;a為體系中Fe(Ⅲ)還原的最大潛勢(shì),即Fe(Ⅱ)的最大累積量;b為模型參數(shù);k為反應(yīng)的速率常數(shù),即 Fe(Ⅱ)的累積速率常數(shù)。Fe(Ⅲ)還原反應(yīng)的最大反應(yīng)速率Vmax=0.25ak;最大反應(yīng)速率對(duì)應(yīng)的時(shí)間tVmax=ln(b/k)。
數(shù)據(jù)處理采用Curve Expert1.4。
2實(shí)驗(yàn)結(jié)果和討論
2.1菌株基本生長(zhǎng)特性
通過(guò)富集及分離純化等過(guò)程,獲得一株具有良好Fe(Ⅲ)還原能力的菌株。該菌株在檸檬酸鐵固體培養(yǎng)基上的菌落較小,呈圓形或散狀生長(zhǎng),菌落顏色為淺黃色,參考文獻(xiàn)[12]初步判別該菌為革蘭氏陰性菌,短桿狀,長(zhǎng)2 μm、寬0.8 μm,如圖1所示。
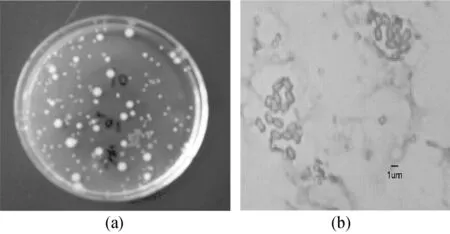
圖1 菌株在檸檬酸鐵固體培養(yǎng)基及顯微鏡下形態(tài)圖
pH值、溫度、培養(yǎng)時(shí)間及光照條件對(duì)菌株生長(zhǎng)[13]的影響如圖2所示。
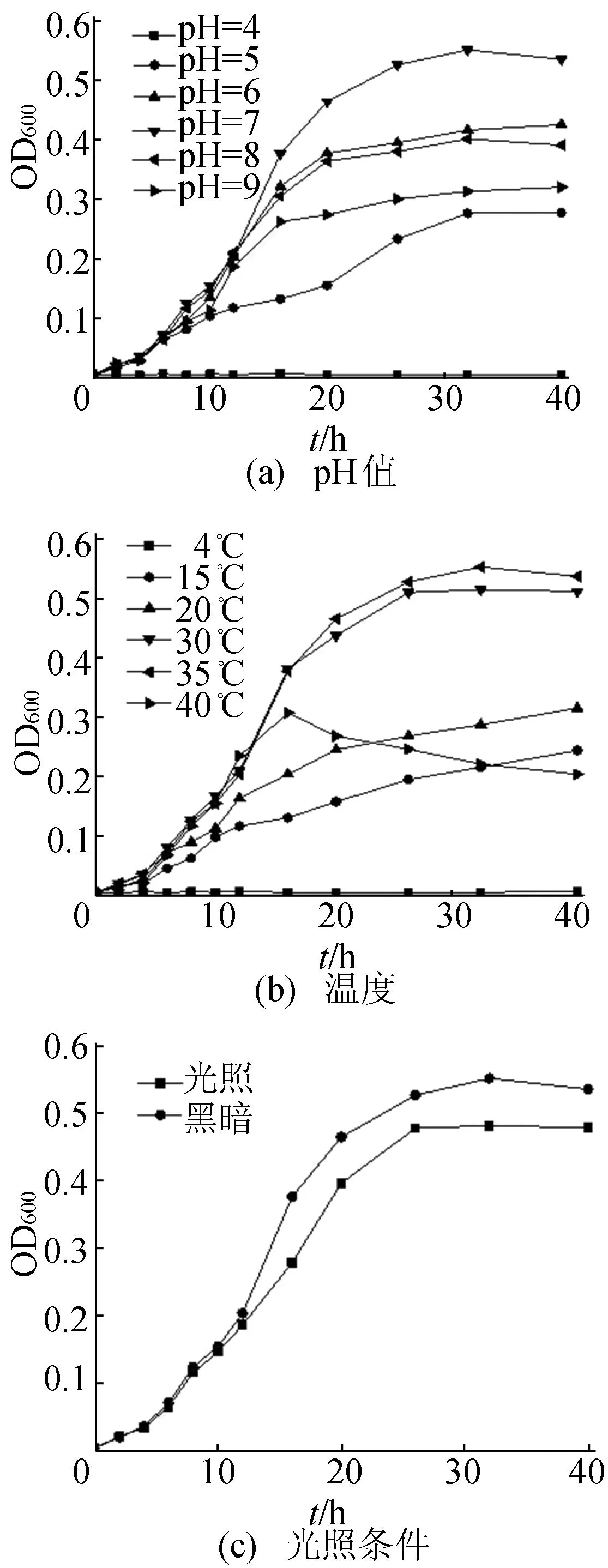
圖2 鐵還原菌在不同條件下菌懸液的光密度值(OD600)變化
圖2a顯示,在30℃、暗培養(yǎng)條件下,pH值分別為4、5、6、7、8、9時(shí),菌液濃度差異較大,菌液濃度在pH=7是菌株生長(zhǎng)最佳條件,其次為pH=6和pH=8,pH=9和pH=5再次之,當(dāng)培養(yǎng)體系pH<4時(shí)不能存活。此株鐵還原菌耐酸堿性能力較強(qiáng)。
在pH=7、暗培養(yǎng)條件下,溫度對(duì)菌株生長(zhǎng)的影響如圖2b所示。由圖2b可見(jiàn),30℃為菌株生長(zhǎng)最適溫度,其次為30℃,而當(dāng)溫度為20、15℃時(shí),菌株生長(zhǎng)速度顯著下降,溫度降至4℃時(shí)菌株停止生長(zhǎng)。溫度為40℃時(shí),培養(yǎng)時(shí)間超過(guò)16 h后,隨著培養(yǎng)時(shí)間的增加,菌濃度顯著下降,菌株表現(xiàn)出較差的耐熱性。
在pH=7、溫度為35℃條件下,暗培養(yǎng)條件下的菌量比光照條件下量多,因此暗培養(yǎng)有利于菌株的生長(zhǎng)。
因此,該株鐵還原菌最佳的生長(zhǎng)條件是pH=7、溫度為35℃、暗培養(yǎng)。
從培養(yǎng)時(shí)間上來(lái)看,在pH=7、溫度為35℃、暗培養(yǎng)條件下,菌株在LB培養(yǎng)基中停滯期比較短,培養(yǎng)12 h左右生長(zhǎng)曲線(xiàn)就進(jìn)入對(duì)數(shù)期,26 h后進(jìn)入穩(wěn)定生長(zhǎng)期,到32 h時(shí)的生物量最大,故在后續(xù)的碳源利用特征試驗(yàn)中,采用擴(kuò)繁32 h處于菌株穩(wěn)定期的菌液進(jìn)行鐵還原實(shí)驗(yàn)。
2.2碳源對(duì)鐵還原菌Fe(Ⅲ)還原能力的影響
2.2.1碳源濃度對(duì)Fe(Ⅲ)還原的影響
碳源濃度對(duì)Fe(Ⅲ)還原的影響如圖3所示(圖注數(shù)值的單位為mol/L)。
圖3中碳濃度為1.5 mol/L時(shí)分別對(duì)應(yīng)蔗糖、葡萄糖、乙酸鈉、乳酸鈉和丙酮酸鈉濃度為0.125、0.25、0.75、0.5、0.5 mol/L,以此類(lèi)推。結(jié)果表明,未添加碳源時(shí),培養(yǎng)體系中幾乎未檢測(cè)到Fe(Ⅱ),說(shuō)明微生物Fe(Ⅲ)還原過(guò)程需要有電子供體才能進(jìn)行。添加不同碳源時(shí),反應(yīng)體系中均可以檢測(cè)到Fe(Ⅱ)的累積。當(dāng)添加碳濃度為1.5 mol/L時(shí),Fe(Ⅱ)的累積量明顯大于0.75、0.375 mol/L碳濃度條件下的累積量。以蔗糖為例,碳濃度添加量為3.0、1.5、0.75、0.375 mol/L時(shí),反應(yīng)體系中Fe(Ⅱ)累積量分別為17.52、17.38、9.24、7.66 mg/L,1.5 mol/L碳濃度條件下Fe(Ⅱ)累積量分別是0.75、0.375 mol/L碳源濃度時(shí)Fe(Ⅱ)累積量的1.9和2.3倍,而3.0 mol/L的碳源濃度下Fe(Ⅱ)累積量并無(wú)顯著的增加。由此可見(jiàn),較低的碳源濃度因不能提供足夠的電子供體,不利于Fe(Ⅲ)還原過(guò)程進(jìn)行;過(guò)高的碳源濃度也未顯著增加Fe(Ⅱ)的累積量,多余的碳源不能有效被菌株利用。因此,選擇1.5 mol/L的碳源濃度是異化鐵還原高效碳源濃度。
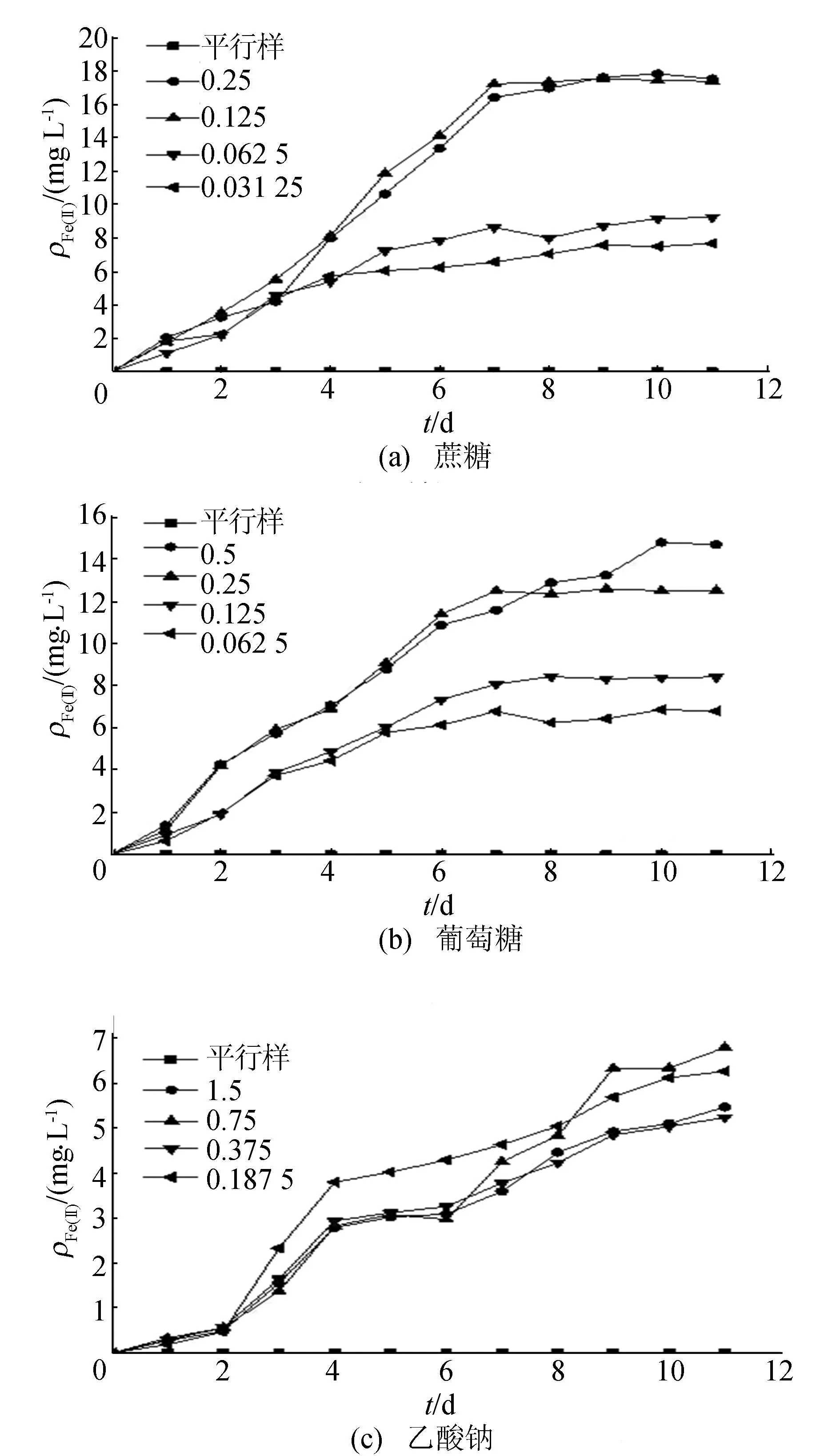
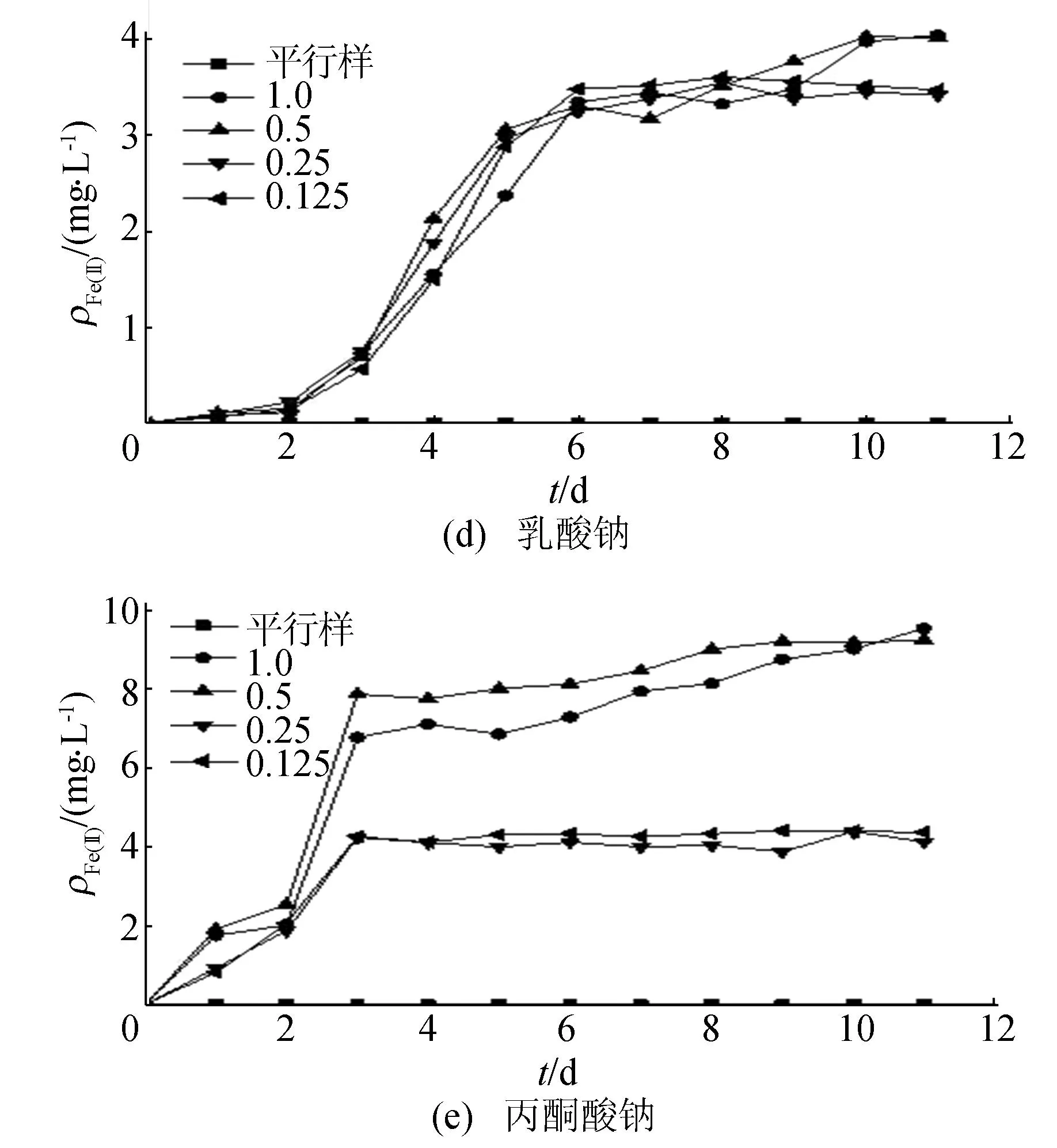
圖3 鐵還原菌利用不同碳源作為電子供體的Fe(Ⅱ)質(zhì)量濃度變化
2.2.2碳源類(lèi)型對(duì)Fe(Ⅲ)還原的影響
在添加不同碳源條件下,反應(yīng)體系中Fe(Ⅱ)的積累量存在明顯的差異,以蔗糖為碳源的體系中Fe(Ⅱ)累積量最多,其后依次為葡萄糖、丙酮酸鈉、乙乳酸鈉和乳酸鈉。在1.5 mol/L最佳碳濃度條件下,蔗糖、葡萄糖、丙酮酸鈉、乙乳酸鈉和乳酸鈉為碳源時(shí),體系中Fe(Ⅲ)還原的比例依次為81.0%、57.2%、42%、30.8%、18.3%。可見(jiàn)該菌以蔗糖為碳源時(shí)的Fe(Ⅲ)還原能力最強(qiáng),而以蔗糖無(wú)氧分解終產(chǎn)物乳酸鈉為碳源時(shí)的Fe(Ⅲ)還原能力最弱。與文獻(xiàn)[14]分離得到的一株鐵/硝酸鹽還原菌38.9%的Fe(Ⅲ)還原率(碳源為葡萄糖)相比,該菌表現(xiàn)出較強(qiáng)的Fe(Ⅲ)還原,而與文獻(xiàn)[15]以葡萄糖為碳源的6株菌24.52%~92.11%的Fe(Ⅲ)還原能力相比,仍具有較好的Fe(Ⅲ)還原能力。
不同碳源條件下,隨著時(shí)間的變化,體系中Fe(Ⅲ)還原變化趨勢(shì)有較明顯的差異。以葡萄糖和蔗糖作為碳源時(shí),前7 d,體系中Fe(Ⅱ)迅速累積,之后體系中Fe(Ⅱ)累積量緩慢增加,到第9天時(shí)趨于穩(wěn)定。以丙酮酸鈉為碳源時(shí),體系中的Fe(Ⅱ)累積量迅速增加,在第3天時(shí)Fe(Ⅱ)累積量已經(jīng)達(dá)到最大累積量(以第11天計(jì))的85.2%,表明鐵還原菌以丙酮酸鈉為碳源時(shí)能在較短時(shí)間內(nèi)達(dá)到較高的Fe(Ⅲ)還原水平。乙酸鈉和乳酸鈉為碳源時(shí),體系存在1~2 d的緩沖期,此時(shí)體系中Fe(Ⅱ)累積水平低,之后Fe(Ⅱ)累計(jì)量迅速增加,乳酸鈉碳源體系中在第6天時(shí)Fe(Ⅱ)累積量已經(jīng)達(dá)到最大累積量的82.3%,之后增幅降低,直到第10天左右Fe(Ⅱ)累計(jì)量趨于穩(wěn)定。綜上可見(jiàn),從Fe(Ⅲ)還原率角度來(lái)看,蔗糖、葡萄糖是最佳碳源;從還原所需時(shí)間角度來(lái)看,丙酮酸鈉是較好的碳源選擇。
2.3Fe(Ⅲ)還原動(dòng)力學(xué)
采用Logistic方程擬合不同碳源對(duì)Fe(Ⅲ)還原的影響,結(jié)果見(jiàn)表2所列。
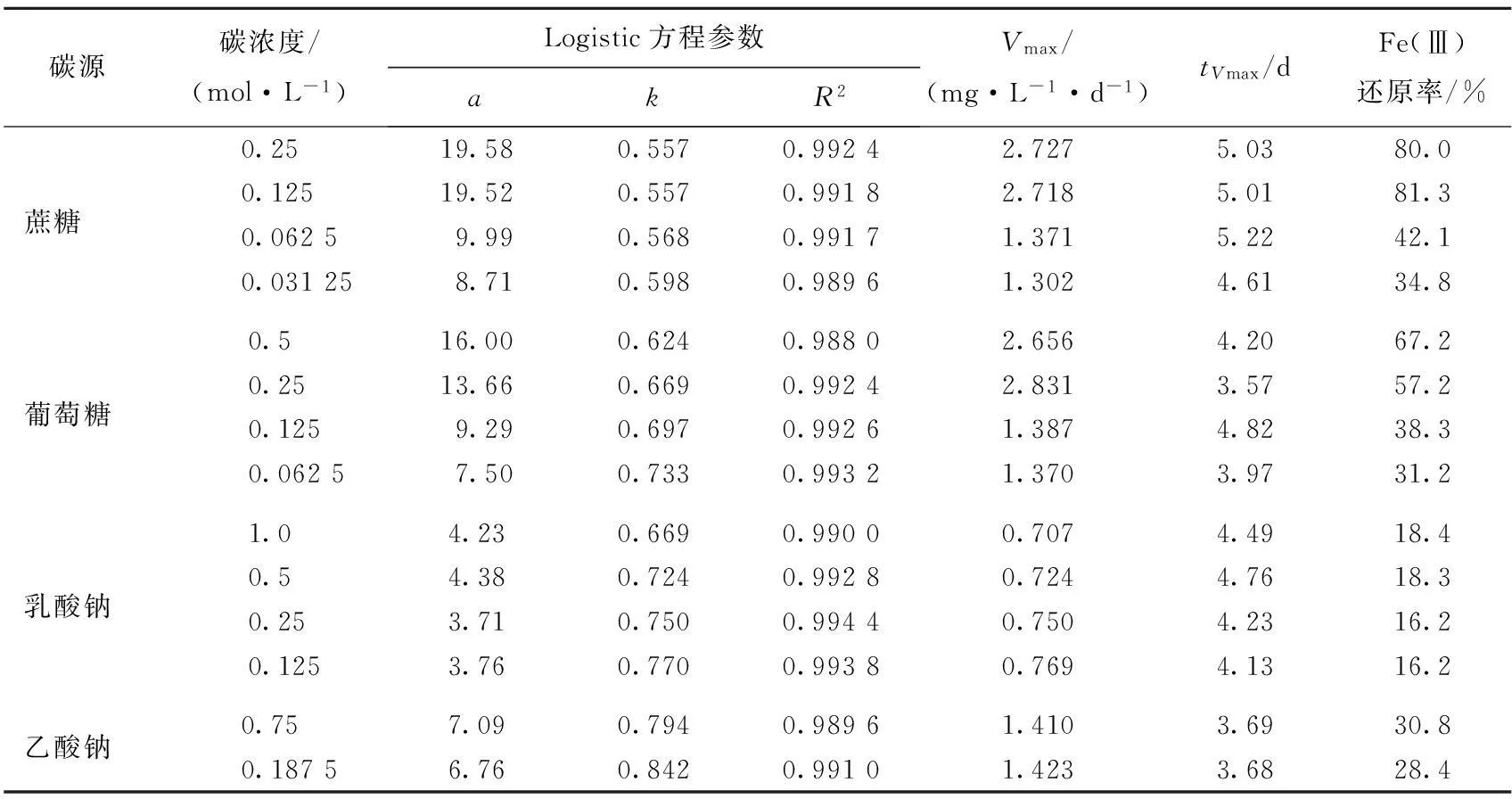
表2 不同碳濃度下鐵還原菌介導(dǎo)的Fe(Ⅲ)還原的動(dòng)力學(xué)擬合
結(jié)果表明,丙酮酸鈉為碳源時(shí),乙酸鈉濃度為1.5、0.375 mol/L時(shí),不能用Logistic方程進(jìn)行Fe(Ⅲ)還原動(dòng)力學(xué)擬合,其他不同碳源條件下均能用Logistic方程擬合Fe(Ⅲ)還原動(dòng)力學(xué)過(guò)程。從還原最大潛勢(shì)a看,蔗糖和葡萄糖作為電子供體時(shí)Fe(Ⅲ)還原的最大潛勢(shì)a隨著碳源濃度增大而增大,而以乳酸鈉和乙酸鈉為電子供體時(shí)Fe(Ⅲ)還原的最大潛勢(shì)與碳源濃度無(wú)明顯的關(guān)聯(lián)。從反應(yīng)速率常數(shù)k看,Fe(Ⅱ)累積速率常數(shù)隨添加的碳源濃度的降低而增加。高濃度的蔗糖(0.25、0.125 mol/L)和高濃度葡萄糖(0.5、0.25 mol/L)的最大反應(yīng)速率Vmax比同系低濃度的蔗糖(0.062 5、0.031 25 mol/L)和葡萄糖(0.125、0.062 5 mol/L)的最大反應(yīng)速率Vmax高出1倍,而高、低濃度乳酸鈉和乙酸鈉體系中最大反應(yīng)速率無(wú)明顯差別。最大反應(yīng)速率對(duì)應(yīng)時(shí)間(tVmax)均在第3~6天,表明鐵還原菌要經(jīng)過(guò)2~3 d生長(zhǎng)期后大量增殖,參與Fe(Ⅲ)還原。Fe(Ⅲ)還原率均是高濃度比低濃度的高,其中蔗糖和葡萄糖隨著濃度的加倍,還原率有較大的變化,而乳酸鈉和乙酸鈉的還原率很接近,與濃度的大小沒(méi)有關(guān)系。這表明蔗糖和葡萄糖對(duì)此株鐵還原菌有更大的還原潛勢(shì)。
2.4不同碳源的脫氫酶活性變化
碳濃度為1.5 mol/L時(shí),研究不同碳源條件下脫氫酶活性,結(jié)果如圖4所示。
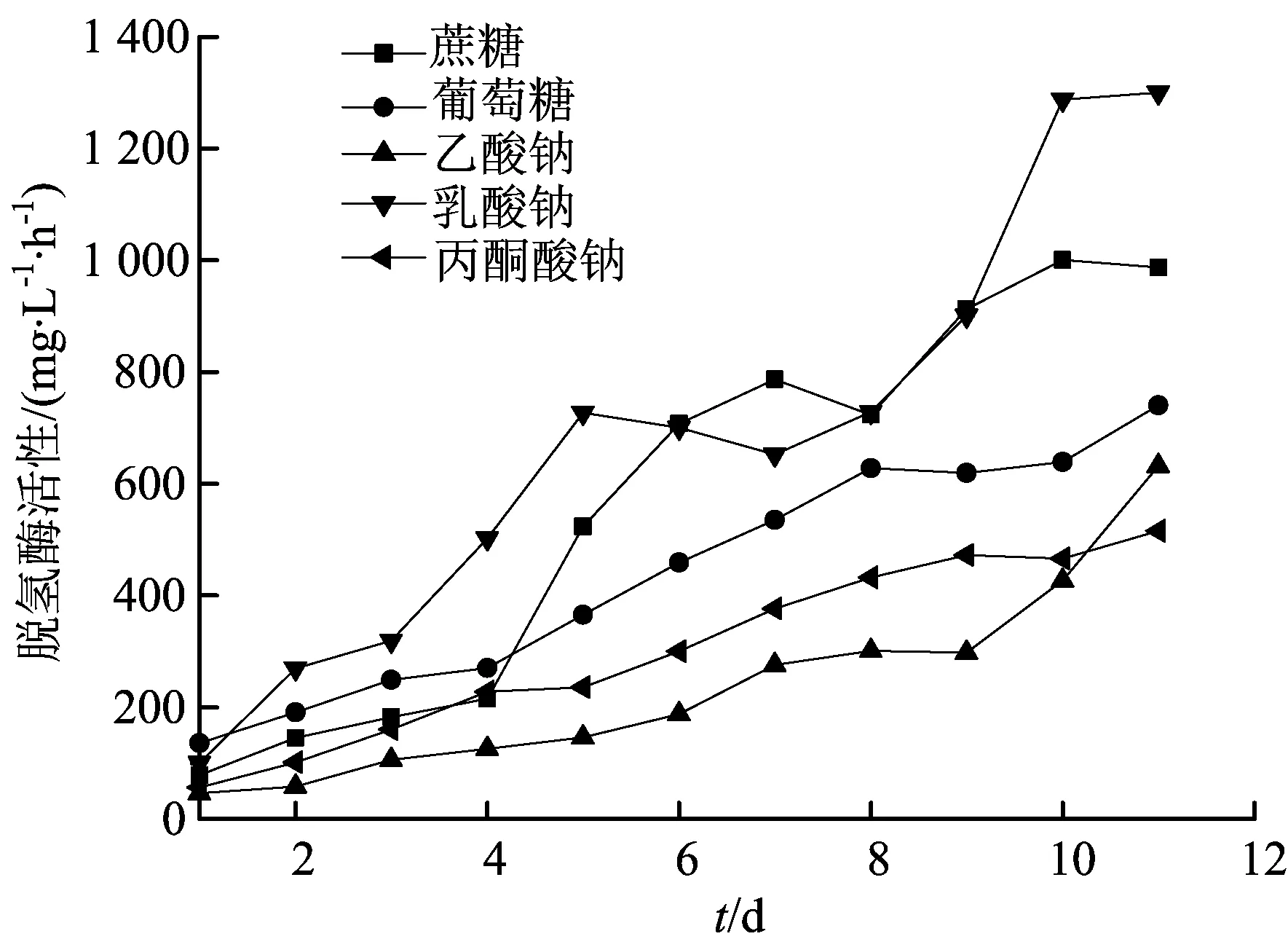
圖4 鐵還原菌利用不同碳源(c=1.5 mol/L)作為電子供體的脫氫酶活性變化
由圖4可知隨著培養(yǎng)時(shí)間增加,各碳源體系中脫氫酶活性增加,然而增長(zhǎng)的幅度和時(shí)間有很大不同。前3 d中,不同碳源體系中脫氫酶活性差異較小,第4天開(kāi)始,不同碳源條件下的脫氫酶活性差異顯著,到第11天時(shí),丙酮酸鈉、蔗糖、葡萄糖、乙酸鈉和乳酸鈉條件下的脫氫酶活性依次為120.41、90.23、67.75、47.20、20.47 mg/(L·h),丙酮酸鈉體系中的脫氫酶活性為乳酸鈉體系中的脫氫酶活性的6倍。
每種碳源體系中的脫氫酶活性與Fe(Ⅱ)濃度的關(guān)系,見(jiàn)表3所列。由表3可知,蔗糖、葡萄糖、乙酸鈉和乳酸鈉碳源體系中Fe(Ⅱ)積累量與脫氫酶活性均為顯著正相關(guān)關(guān)系,而丙酮酸鈉碳源體系中Fe(Ⅱ)積累量與脫氫酶活性的相關(guān)系數(shù)明顯低于其他4種碳源體系,但兩者間仍為正相關(guān)。
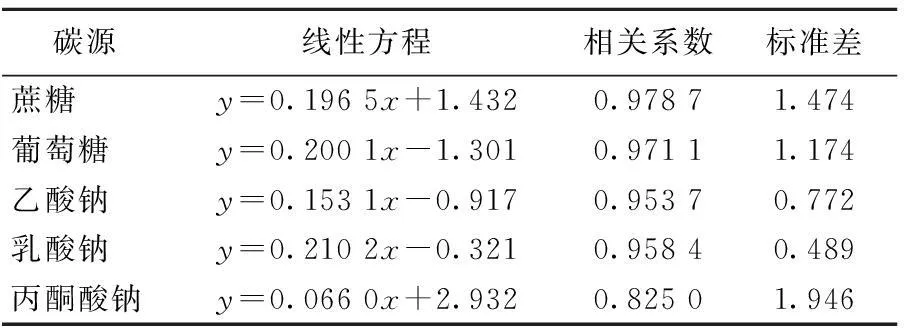
表3 Fe(Ⅱ)濃度(y)與脫氫酶活性(x)的相關(guān)性
注:當(dāng)α=0.05,r0.05=0.602 1;當(dāng)α=0.01,r0.01=0.734 8;當(dāng)α=0.001,r0.001=0.847 1。
對(duì)比不同碳源間的脫氫酶活性與Fe(Ⅱ)濃度可以發(fā)現(xiàn),雖然丙酮酸鈉碳源體系中脫氫酶活性顯著大于其余碳源體系,但其Fe(Ⅱ)濃度小于蔗糖和葡萄糖碳源體系。這可能是由于蔗糖、葡萄糖在無(wú)氧條件下酵解為丙酮酸,提供給鐵還原菌更多的能量,而丙酮酸還原生成乳酸的過(guò)程產(chǎn)能低,提供給鐵還原菌可利用的能量低,從而導(dǎo)致丙酮酸鈉碳源體系中雖然有高的脫氫酶活性,但體系中Fe(Ⅱ)積累較少。因此,脫氫酶活性可以在某種程度上反映碳源的利用程度,也能間接反映異化鐵還原程度[16]。
綜合不同碳源條件下的脫氫酶活性及Fe(Ⅱ)累積濃度分析,該菌具有利用多種碳源為電子供體異化還原Fe(Ⅲ)的能力,在環(huán)境污染物控制方面具有較好的應(yīng)用前景,在污水處理中具有較好的應(yīng)用前景。
3結(jié)論
(1)分離獲得的鐵還原菌可耐受pH值為5~9的酸堿度變化,其最佳生長(zhǎng)條件為pH=7、溫度為35℃、黑暗條件。
(2)在不同碳源條件下,異化鐵還原最佳碳濃度均為1.5mol/L。蔗糖和葡萄糖是該菌株的最佳碳源,其后依次是丙酮酸鈉、乙酸鈉和乳酸鈉。蔗糖和葡萄糖為碳源時(shí),Fe(Ⅲ)還原率分別為81.0%和57.2%。
(3)碳源明顯影響著異化鐵還原過(guò)程中的脫氫酶活性,各碳源體系脫氫酶活性依次為丙酮酸鈉>蔗糖>葡萄糖>乙酸鈉>乳酸鈉。碳源體系中,除去丙酮酸鈉,脫氫酶活性與Fe(Ⅱ)累積量呈顯著正相關(guān),脫氫酶活性在某種程度上能反應(yīng)異化鐵還原程度。
[參考文獻(xiàn)]
[1]WeberK A,AchenbachL A,Coates J D.Microorganisms pumping iron:anaerobic microbial iron oxidation and reduction[J].Nature Review of Microbiology,2006,45(6):752-764.
[2]曾洪學(xué),楊玉泉.異化Fe(Ⅲ)還原菌及其還原機(jī)制的環(huán)境意義[J].安徽農(nóng)業(yè)科學(xué),2012(18):9843-9846.
[3]賀江舟,曲東,張莉莉.Fe(Ⅲ)的微生物異化還原[J].微生物學(xué)通報(bào),2006,33(5):158-164.
[4]Lovley D R,Phillips E J P.Organic matter mineralization with reduction of ferric iron in anaerobic sediments[J].Applied and Environmental Microbiology,1986;51(4):683-689.
[5]Noguera D R,Brusseau G A,Rittmann B E,et al.A unified model describing the role of hydrogen in the growth of Desulfovibrio vulgaris under different environmental conditions[J].Biotechnology and Bioengineering,1998,59(6):732-746.
[6]葉明.微生物學(xué)實(shí)驗(yàn)技術(shù)[M].合肥:合肥工業(yè)大學(xué)出版社,2009:24-26.
[7]孫麗蓉,曲東,易維杰.多糖對(duì)水稻土中異化Fe(Ⅲ)還原過(guò)程的影響[J].河南農(nóng)業(yè)科學(xué),2010(6):62-66.
[8]Tian Q,Chen J,Zhang H,et al.Study on the modified triphenyl tetrazolium chloride(TTC-DHA)method in determination of bioactivity in the up-flow aerated bio-activated carbon filter[J].African Journal of Biotechnology,2006,5(2):181-188.
[9]尹軍,譚學(xué)軍,張立國(guó),等.測(cè)定脫氫酶活性的萃取劑選擇[J].中國(guó)給水排水,2004,20(7):96-98.
[10]易維潔,曲東,賈蓉.不同淹水時(shí)間水稻土微生物群落的還原能力及其與脫氫酶活性的關(guān)系[J].應(yīng)用基礎(chǔ)與工程科學(xué)學(xué)報(bào),2012,20(5):768-776.
[11]劉碩,曲東.苯系物作為唯一碳源對(duì)異化鐵還原過(guò)程的影響[J].西北農(nóng)林科技大學(xué)學(xué)報(bào):自然科學(xué)版,2006,34(10):101-106.
[12]布坎南.伯杰細(xì)菌鑒定手冊(cè)[M].第8版.北京:科學(xué)出版社,1984:535-587.
[13]葛曉光,楊柳,彭申華,等.一株煤礦地下水硫酸鹽還原細(xì)菌的分離、鑒定及性質(zhì)研究[J].合肥工業(yè)大學(xué)學(xué)報(bào):自然科學(xué)版,2011,34(3):420-423.
[14]汪國(guó)威.一株鐵/硝酸鹽還原菌分離、特性鑒定及其與鐵氧化物作用[D].合肥:合肥工業(yè)大學(xué),2013.
[15]王偉民.水稻土中鐵還原菌的分離純化及鐵還原能力鑒定[D].西安:西北農(nóng)林科技大學(xué),2007.
[16]賈蓉,曲東,喬莎莎.發(fā)酵脫氫產(chǎn)氫過(guò)程對(duì)微生物鐵還原的影響[J].農(nóng)業(yè)環(huán)境科學(xué)學(xué)報(bào),2013,32(12):2395-2402.
(責(zé)任編輯張淑艷)
Isolation of an iron-reducing bacteria strain and its carbon source utilization
TONG Lei,LI Xiang-ling,WU Ji-nan,YUAN Feng,ZHOU Tao-fa
(School of Resources and Environmental Engineering,Hefei University of Technology,Hefei 230009,China)
Abstract:An iron-reducing bacteria strain was isolated from municipal sewage sludge.The characteristics of the bacteria strain and the influence of carbon sources on the Fe(Ⅲ)reduction were studied.The results showed that the strain grew best when pH value was 7 and the temperature was 35℃ under the darkness condition.The iron reduction ability of the bacteria strain was significantly influenced by the type and the concentration of carbon source.The optimum concentration of the carbon source was 1.5 mol/L.When sucrose,glucose,sodium pyruvate,sodium acetate,sodium lactate were serviced as carbon sources respectively,Fe(Ⅲ)reduction rate of the strain reduced in turn.When sucrose and glucose were used as carbon sources,Fe(Ⅲ)reduction rate of the strain was 81.0% and 57.2% respectively.There was a significant positive correlation between the dehydrogenase activity of the strain and Fe(Ⅲ)reduction rate.Dehydrogenase activity could reflect the Fe(Ⅲ)reduction degree.
Key words:iron-reducing bacteria;carbon source;dehydrogenase activity
收稿日期:2015-01-04;修回日期:2015-03-04
基金項(xiàng)目:環(huán)保部公益性行業(yè)科研專(zhuān)項(xiàng)經(jīng)費(fèi)資助項(xiàng)目(201009041-03)
作者簡(jiǎn)介:童磊(1988-),男,安徽蕪湖人,合肥工業(yè)大學(xué)碩士生;周濤發(fā)(1964-),男,安徽廬江人,博士,合肥工業(yè)大學(xué)教授,博士生導(dǎo)師.
doi:10.3969/j.issn.1003-5060.2016.04.021
中圖分類(lèi)號(hào):X172
文獻(xiàn)標(biāo)識(shí)碼:A
文章編號(hào):1003-5060(2016)04-0536-07
袁峰(1971-),男,廣西桂林人,博士,合肥工業(yè)大學(xué)教授,博士生導(dǎo)師;
