三疣梭子蟹Spook基因克隆及其表達分析
2016-06-23 13:49:50周彥琦朱冬發(fā)
生物技術通報 2016年5期
關鍵詞:分析
周彥琦 朱冬發(fā)
(寧波大學海洋學院,寧波 315211)
三疣梭子蟹Spook基因克隆及其表達分析
周彥琦 朱冬發(fā)
(寧波大學海洋學院,寧波 315211)
甲殼動物蛻皮激素(Ecdysteroids)主要在Y器官(YO)中合成與分泌,參與調控蛻皮、繁殖等多項生理活動。Spook基因編碼的細胞色素P450(CYP)307a1是蛻皮激素合成通路早期的關鍵酶。為了研究其在三疣梭子蟹蛻皮過程中的調控作用,采用反轉錄PCR(RT-PCR)和cDNA末端快速擴增(RACE)技術,克隆得到了三疣梭子蟹Spook基因的全長cDNA序列(Gen-Bank 登錄號:KM030021)。該序列全長為2 200 bp,包含一個長度為1 563 bp的開放閱讀框,編碼520個氨基酸;比對分析顯示該氨基酸序列含helix-C、helix-K、helix-I、PERF、heme-binding 5個P450特征保守區(qū)域;系統(tǒng)進化樹分析發(fā)現推導的三疣梭子蟹Spook蛋白與其他物種Spook聚為一支,而其他Halloween基因則分別聚為一支,表明推導的氨基酸確實是三疣梭子蟹Spook的蛋白序列。采用實時熒光定量PCR(qPCR)技術分析了其在不同組織中的表達情況,結果顯示Spook基因主要在三疣梭子蟹的YO中表達。在三疣梭子蟹的蛻皮周期中,Spook基因的表達水平自蛻皮后期(A、B期)逐漸上升,并在蛻皮間期(C期)上升至最大,隨后在蛻皮前期逐漸下降至D3、D4亞期最低。研究結果表明Spook基因可能參與調控三疣梭子蟹的蛻皮過程。
三疣梭子蟹;Spook;蛻皮周期;基因克隆;表達水平
蛻皮激素(Ecdysteroids)在節(jié)肢動物的生長、繁殖和形態(tài)發(fā)生方面起著重要的作用,包括個體發(fā)育、變態(tài)、蛻皮、肌肉再生和能量儲備等多種生物過程[1]。在周期性蛻皮過程中蛻皮激素刺激表皮發(fā)生分離并啟動一系列與蛻皮相關的重要生理過程[2]。蛻皮激素是一種類固醇激素,最早從蛾蛹中分離得到[3]。隨后Huber[4]確定其化學結構并發(fā)現分泌組織為昆蟲前胸腺(Prothoracic gland,PG)。
在甲殼動物中,蛻皮激素由位于其頭胸部前端的Y器(Y-organ,YO)所分泌,并在外周組織中轉化為其活性形式,一般為20-羥基蛻皮酮(20-Hydroxyecdysone,20E)[5]。YO中蛻皮激素的生物合成的途徑包括兩個階段:第一階段是由含有Riseke功能結構域的Neverland(Nvd)蛋白催化膽固醇向7-脫氫膽固醇(7-dehydrocholesterol,7DC)的轉化[6],然后7DC被轉運至線粒體中轉化為5β-diketol(3D2,22,25,dE)。7DC轉化為5β-diketol的機制尚不清楚,被稱為“黑匣子(black box)”。目前普遍認為Spook(CYP307a1)、Spookier(CYP307a2)和Spookiest(CYP307B1)參與該階段的調控[7]。第二階段包括5β-diketol經過Phantom、Disembodied和Shadow分別編碼的CYP306a1、CYP-302a1和CYP315a1的催化作用,其C25、C22和C5發(fā)生羥基化進而合成YO最終的分泌產物[8,9],產物種類因物種及蛻皮時期的差異而有所不同。此外,Shade編碼的CYP314a1參與催化YO分泌的無活性蛻皮激素的活化過程[10,11]。上述Neverland(Nvd)、Spook(spo)、Spookier(spok)、Spookiest(spokest)、Phantom(phm)、Disembodied(Dib)、Shadow(Sad)和Shade被統(tǒng)稱為“Halloween家族基因”。
Halloween家族基因最早發(fā)現自一系列果蠅(Drosophila melanogaster)胚胎發(fā)育突變體的性狀研究[12-14]。果蠅Phm、Dib、Sad和Shd突變體表型十分相似,都在胚胎發(fā)育后期出現表皮不能分化、頭部內翻、背部閉合失敗和中腸缺失等異常表型。這些突變體大多在胚胎發(fā)育末期死亡,且表現出來的性狀與蛻皮激素缺失有關[10,15-17],因此Halloween家族基因在果蠅蛻皮激素合成過程中有著重要意義。
目前,甲殼動物蛻皮激素的研究主要集中在其活性產物的分離鑒定及生理作用上[18-21]。對于Halloween基因及調控機制的研究則少有報道。本研究以三疣梭子蟹(Portunus trituberculatus)為研究對象,通過RT-PCR和RACE技術克隆獲得了三疣梭子蟹Spook基因(PtSpook)的全長cDNA,并運用實時熒光定量PCR(qRT-PCR)分析了其組織分布情況及在蛻皮周期YO中的表達變化,旨在為進一步理解Halloween家族基因在蛻皮激素合成過程中的作用,為深入探討三疣梭子蟹蛻皮的調控機制奠定基礎。
1 材料與方法
1.1 材料
選取健康的野生三疣梭子蟹,暫養(yǎng)于寧波市寧海縣得水育苗場。根據形態(tài)學特征變化研究,將三疣梭子蟹蛻皮周期分為蛻皮后期(A、B期)、蛻皮間期(C期)、蛻皮前期(D0、D1、D2、D3和D4亞期)和蛻皮期(E)5個階段[22]。采集C期蟹(頭胸甲寬(CW)為 15.0-16.5 cm、體質量(BW)為170-265 g)的表皮、卵巢、精巢、肝胰腺、大顎器、眼柄、胸神經節(jié)、Y器和心臟等9個組織用于分析PtSpook在三疣梭子蟹組織分布情況;采集蛻皮周期各期(E期除外)蟹(CW:8-12 cm、BW:42-83 g)的Y器用于分析Spook在蛻皮周期中的表達水平變化。每期取5只蟹用于平行實驗。各組織在冰上解剖后被轉移至RNA保護液(上海生工)-20℃保存?zhèn)溆谩?/p>
1.2 方法
1.2.1 總RNA提取 將上述組織樣品按照Trizol(上海生工)說明進行總RNA提取。總RNA用DNase I(TaKaRa)去除基因組DNA后,用瓊脂糖凝膠電泳檢測其完整性,并用超微量分光光度計Nanodrop 2000(Thermo scientific)進行純度分析。
1.2.2 三疣梭子蟹Spook基因的克隆 采用RT-PCR進行PtSpook核心片段的擴增。模板cDNA由YO總RNA通過 PrimeScriptRTreagent Kit試劑盒(TaKaRa)反轉錄獲得,擴增引物設計自NCBI已知物種Spook序列的保守區(qū)域(表1)。根據得到核心序列設計基因特異性引物,利用RACE技術分別對PtSpook的3'和5'端進行巢式PCR擴增(表1)。其中,3'RACE-cDNA將接頭引物AP替換上述試劑盒中的OligodT primer和Random 6 mers后用相同的方法進行制備,5'RACE-cDNA按照 SMARTerTMRACE cDNA Amplification Kit試劑盒(Clontech)說明書進行合成(表1)。以上PCR均在25 μL體系下進行反應,反應條件如下:94℃預變性5 min;94℃ 50 s,57℃ 50 s,72℃ 1 min 50 s,34個循環(huán);72℃延伸10 min。所有PCR產物經瓊脂糖電泳后,用DNA回收試劑盒(上海生工)進行回收并純化,再與pMD18-T載體連接并轉化至E. coli DH5α感受態(tài)細胞進行培養(yǎng),之后選取陽性克隆菌落交由上海生工生物工程有限公司進行測序。

表1 PCR引物序列
1.2.3 序列分析 利用VectorNTI 10.0對測序結果進行分析和拼接,獲得三疣梭子蟹Spook的cDNA全長。ORF Finder確定開放閱讀框(Open reading frame,ORF),并翻譯成氨基酸序列。BLAST(http://blast. ncbi.nlm.nih.gov/Blast.cgi)分析與其他物種相似性。ExPASy Proteomics Server(http://ca.expasy.org/)對氨基酸序列進行蛋白預測分析 ;SMART(http://www. cbs.dtu.dk/services/TMHMM/)分 析氨基酸結構;用 Signal 4.1 Server(http:// www.cbs.dtu.dk/services/ SignalP)預測氨基酸序列信號肽;利用在線TMHMM工具(http://www.cbs.dtu.dk/services/TMHMM/)預測氨基酸跨膜區(qū)域;利用ClustalX軟件將該氨基酸序列與已公布的Spook氨基酸序列進行同源性比對分析,并用MEGA 6.0軟件的鄰位法(Neighbor-Joining)構建系統(tǒng)進化樹。
1.2.4 實時熒光定量PCR(qPCR)分析 將取好的組織抽取總RNA,取1.0 μg的RNA用Perfect Real Time PrimeScript?RT reagent Kit(TaKaRa)進行反轉錄獲得cDNA。根據PtSpook cDNA全長設計一對熒光定量引物spQF1和spQR1(表1)檢測PtSpook表達水平,Actin-F和Antin-R(表1)擴增三疣梭子蟹β-actin基因(FJ641977.1)作為內參。標準曲線用于驗證PtSpook和β-actin基因引物的效率。按照SYBR?Premix Ex TaqTMII kit(TaKaRa)使用說明進行qPCR反應。反應條件如下:95℃ 2 min;95℃ 5 s,58℃ 20 s,68℃ 30 s,共40個循環(huán)。設定熔解曲線用于保證產物的特異性,條件如下:55-95℃,每秒上升0.2℃。每個cDNA重復3個平行。數據用±SEM方法表示。采用 2-△△Ct方法計算目的基因相對表達量[23]。選重復組中的一個組作為1。其他組的數值以重復組的倍數來表示。用SPSS Statistics軟件進行單因素方差(ANOVA,Duncan’s test)分析,利用Excel對統(tǒng)計結果進行作圖,P<0.05表示顯著性差異。
2 結果
2.1 三疣梭子蟹Spook基因cDNA序列分析
將PCR產物的測序結果進行拼接,得到全長為2 200 bp的三疣梭子蟹Spook的cDNA序列。該序列已登錄GenBank(登錄號:KM030021),包含524 bp 的5'端非編碼區(qū),113 bp的3'端非編碼區(qū)和1 563 b p的開放閱讀框(ORF)(圖1)。ORF編碼520個氨基酸。ExPASy ProtParam Tool預測的蛋白分子式為C6472H10746N2200O2624S691,分子量大小約18.3 kD,等電點為4.84。Signal 4.1 Server和THMMHM分析發(fā)現其氨基酸序列第1位至第21位為氨基酸信號肽序列,并且有跨膜結構域。Blast比對發(fā)現,推導的氨基酸序列與果蠅(Drosophila melanogaster)、家蠶(Bombyx mori)、煙草天蛾(Manduca sexta)的相似性分別為36%、33%和37%。ClustalX軟件比對分析顯示,該氨基酸序包含經典的P450特征保守區(qū)域:helix-C、helix-I、helix-K、PERF和Hemebinding,且在其N末端有一段膜疏水序列(圖2)。通過不同物種Halloween基因構建的系統(tǒng)進化樹(圖3)顯示,推導的氨基酸序列與Spook聚為一支,親緣關系較近。以上結果表明推導的氨基酸序列即為三疣梭子蟹的Spook的蛋白序列。

圖1 三疣梭子蟹Spook的cDNA核苷酸序列和編碼區(qū)氨基酸序列

圖2 三疣梭子蟹Spook氨基酸序列與其他物種Spook氨基酸序列比對
2.2 組織表達差異分析
運用qPCR檢測了Pt Spook在三疣梭子蟹表皮、卵巢、精巢、肝胰腺、大顎器、眼柄、胸神經節(jié)、Y器和心臟等9個組織中的表達情況,結果(圖4)表明PtSpook表達量在Y器(YO)中最高,極顯著高于眼柄、心臟、表皮等其他組織(P<0.05)。
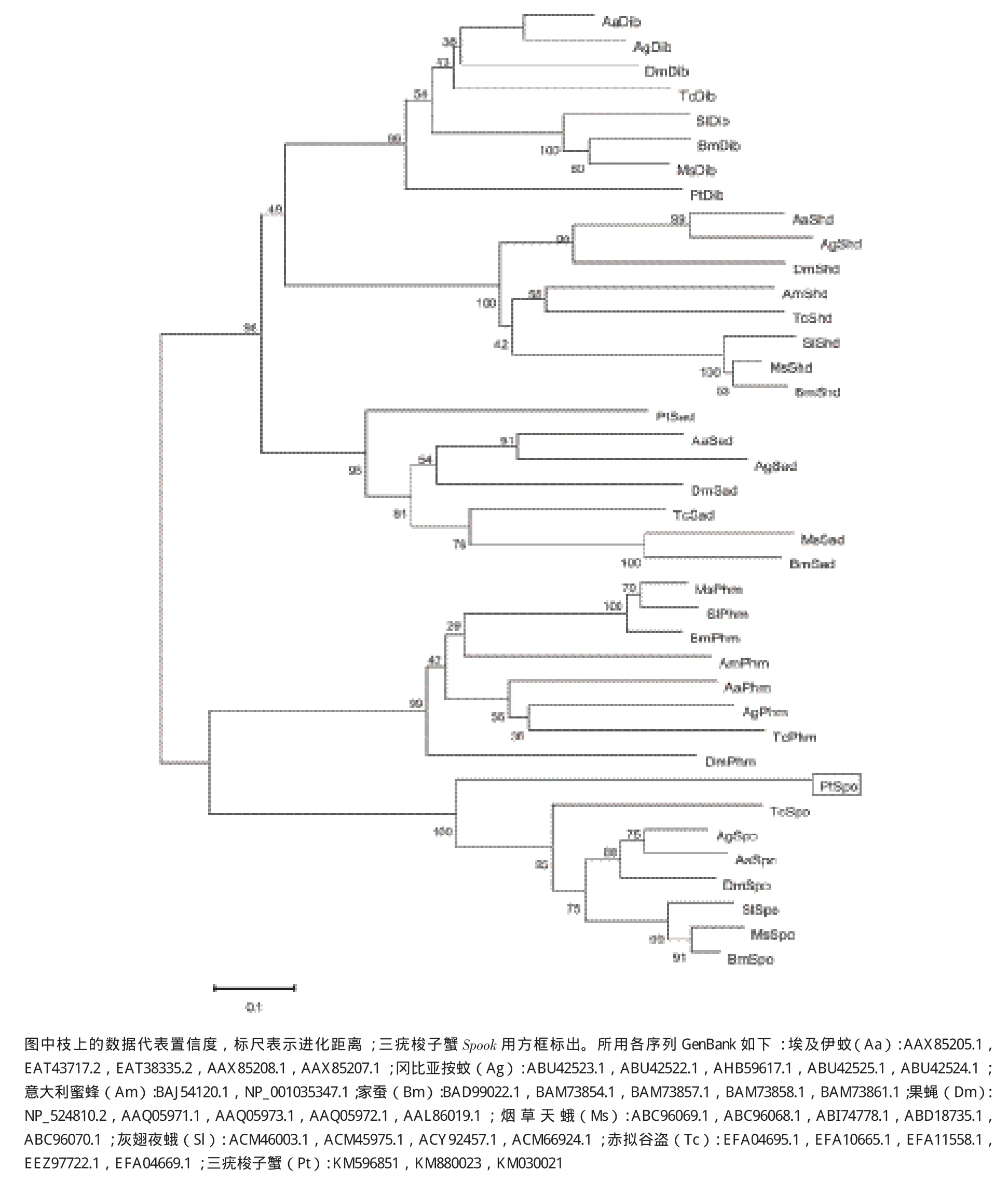
圖3 Spook基因系統(tǒng)進化樹
2.3 三疣梭子蟹Spook基因在蛻皮周期中的表達水平變化
根據組織表達結果,選取YO研究PtSpook在蛻皮周期中的表達變化。結果(圖5)顯示,PtSpook表達量在蛻皮后期(A期和B期)逐漸上升(P< 0.05),并在C期升至最大,之后在蛻皮前期逐漸下降,至D3、D4亞期降至最低。
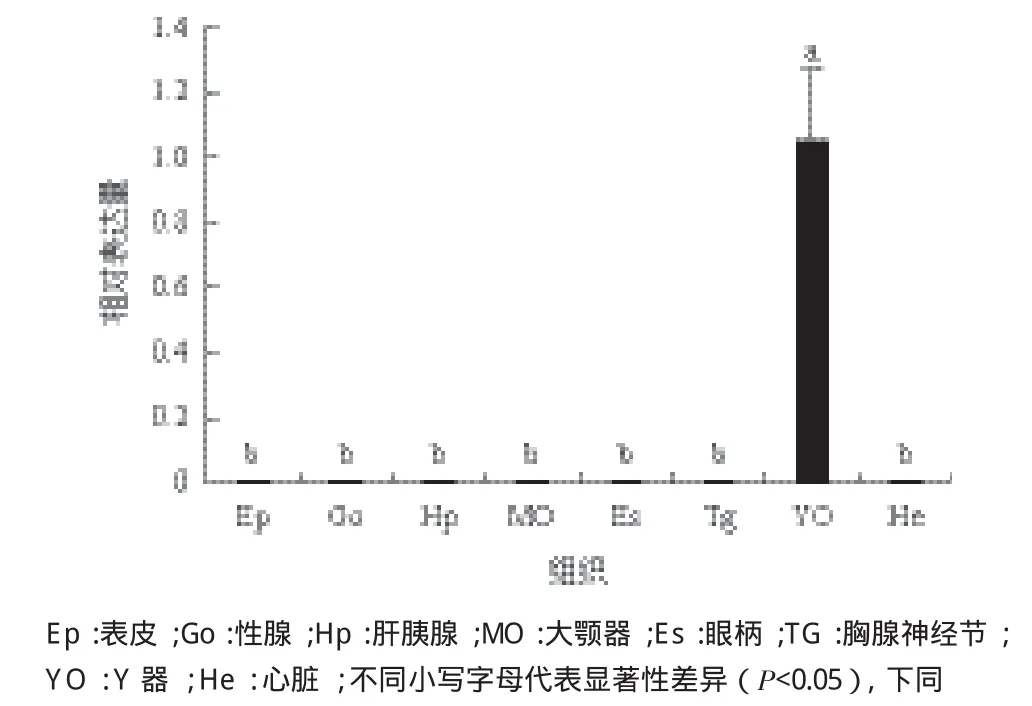
圖4 Spook在三疣梭子蟹不同組織中的相對表達量
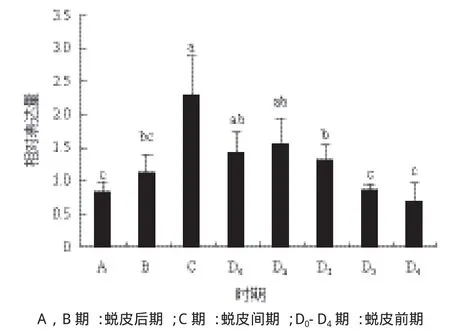
圖5 YO中Spook在三疣梭子蟹中蛻皮周期的相對表達量
3 討論
本研究克隆獲得了三疣梭子蟹Spook基因的全長cDNA序列。Spook基因參與編碼Spook,屬于細胞色素P450超基因家族,該家族是由數量眾多、功能復雜的一類血紅蛋白酶基因組成,廣泛存在于細菌、真菌和動植物生物中。其具有多樣化的氧化酶作用,參與生物體中眾多重要的生理過程[24]。很多其他Halloween基因諸如Spooki er、Spookiest、Phantom、Disembodied、Shadow也都各自編碼不同的細胞色素P450。細胞色素P450家族成員雖然在氨基酸序列變異性很高,但仍存在一定的保守結構域。昆蟲中報道的保守結構域包括helix-C、helix-I、helix-K、PERF和 heme-binding。Heme-binding(FXXGXXXCXG)為細胞色素P450的標志性基序。血紅素鐵接受電子被還原后再與CO結合,會在450 nm附近產生特征吸收峰,heme-binding中絕對保守的半胱氨酸作為血紅素鐵的第5配體參與此過程[25]。helix-C(WXXR)中的精氨酸能夠與血紅素的丙酸酯相結合。helix-I(AGXXT)基序能與氧原子結合,推測與血紅素的遠端側的質子傳遞溝(proton transfer groove)有關。helix-K(EXXR)推測能與氫原子結合,與helix-K形成鹽橋而參與穩(wěn)定血紅素核心結構的作用[26]。保守域PERF(PXXF)屬于芳香族。P450還保有跨膜信號序列,是由甲硫氨酸開始15-20個疏水性的氨基酸構成,信號序列后 富含脯氨酸(P/G-rich),主要功能是指導P450 進入膜內或內質網腔中[27]。本研究推導的氨基酸序列經多重比對后發(fā)現,包含這5個保守結構域(圖2)和信號肽序列(圖1),證明其屬于P450家族基因。此外,系統(tǒng)進化樹顯示該氨基酸序列與其他物種Spook一致性較高,共同聚為一支,鱗翅目(Lepidoptera)家蠶和煙草天蛾等聚為一支,再與雙翅目(Diptera)昆蟲果蠅等組成的分支聚為另一大支,最后與甲殼動物三疣梭子蟹聚為泛甲殼動物大類(圖3)。以上結果與Rewitz等[28]和Zhou等[29]的研究結果一致,由此推斷該序列為三疣梭子蟹Spook基因。
qRT-PCR結果表明,PtSpook在三疣梭子蟹的多個組織中均有不同程度的表達,其中YO中的表達極顯著(P<0.05),該結果與三疣梭子蟹Disembodied基因的組織表達的特異性一致[30]。這進一步證實了YO是甲殼動物合成蛻皮激素的主要器官[31]。另外,PtSpook與昆蟲Spook基因的組織表達特性也極其相似,昆蟲Spook基因主要在其前胸腺(PG)組織中表達[29,32],這說明甲殼動物的YO和昆蟲PG在功能上具有相似性。
作為Halloween家族基因成員之一,Spook通過參與蛻皮激素的合成,進而調控昆蟲的多項生理功能。Chávez等[15]用酶免疫分析(EIA)方法檢測到spook突變體中蛻皮酮(ecdysone)和20E含量極低,也無法檢測到IMP-E1和L1等蛻皮激素后期響應基因的表達,表明果蠅的Spook基因參與了蛻皮激素合成。Namiki等[33]應用熒光標記差異顯示技術得到了家蠶(Bombyx mori)的Spook基因,并發(fā)現其基因表達水平與血淋巴中蛻皮激素滴度呈正相關,同時還發(fā)現Spook突變后胚胎中蛻皮激素濃度降低并導致胚胎發(fā)育停滯。Ono等[32]在微粒體中檢測到Spook編碼產物,Spook突變體中添加20E、蛻皮酮或者三脫氧蛻皮酮都能挽救胚胎和幼蟲發(fā)育停滯。本實驗同樣表明Spook基因可能通過參與三疣梭子蟹蛻皮激素的合成,進而實現對蛻皮的調控。在三疣梭子蟹的蛻皮周期中,Spook基因的表達水平與之前報道的三疣梭子蟹血淋巴中蛻皮激素濃度[34]和YO中Disembodied基因的表達水平[30]有著良好的協(xié)同關系,且其表達水平峰值出現較早。該結果表明三疣梭子蟹Spook作為蛻皮激素早期合成酶,其表達水平與下游合成酶和蛻皮激素合成均有著密切的聯(lián)系。
4 結論
本研究首次克隆獲得了三疣梭子蟹Spook基因的全長cDNA序列,其氨基酸序列與其他物種Spook一致性較高,共同聚為一支。組織差異表達分析顯示三疣梭子蟹Spook在YO中最高,顯著高于其他所檢組織。在三疣梭子蟹蛻皮周期過程中,PtSpook在YO的表達水平自蛻皮后期(A、B期)逐漸上升,并在蛻皮間期(C期)上升至最大,隨后在蛻皮前期逐漸下降至D3、D4亞期最低。該結果表明PtSpook可能參與調控三疣梭子蟹的蛻皮過程。
[1]Kuballa AV, Holton TA, Paterson B, et al. Moult cycle specific differential gene expression profiling of the crab Portunus pelagicus[J]. BMC Genomics, 2011, 12(1):147.
[2]Skinner DM. Molting and regeneration[J]. The Biology of Crustacea, 1985, 9:143-146.
[3]Butenandt A, Karlson P. über die isolierung eines metamorphosehormons der insekten in kristallisierter form[J]. Zeitschrift für Naturforschung B, 1954, 9(6):389-391.
[4]Huber R, Hoppe W. Zur Chemie des Ecdysons, VII:Die Kristallund Molekülstrukturanalyse des Insektenverpuppungshormons Ecdyson mit der automatisierten Faltmolekülmethode[J]. Chemische Berichte, 1965, 98(7):2403-2424.
[5]Lachaise F, Le Roux A, Hubert M, et al. The molting gland of crustaceans:localization, activity, and endocrine control(a review)[J]. J Crust Biol, 1993, 13(2):198-234.
[6]Yoshiyama T, Namiki T, Mita K, et al. Neverland is an evolutionally conserved Rieske-domain protein that is essential for ecdysone synthesis and insect growth[J]. Development, 2006, 133(13):2565-2574.
[7]Thummel CS, Chory J. Steroid signaling in plants and insects—common themes, different pathways[J]. Genes & Development, 2002, 16(24):3113-3129.
[8]B?cking D, Dauphin-Villemant C, Sedlmeier D, et al. Ecdysteroid biosynthesis in moulting glands of the crayfish Orconectes limosus:evidence for the synthesis of 3-dehydroecdysone by in vitro synthesis and conversion studies[J]. Insect Biochemistry and Molecular Biology, 1993, 23(1):57-63.
[9]Mykles DL. Ecdysteroid metabolism in crustaceans[J]. The Journal of Steroid Biochemistry and Molecular Biology, 2011, 127(3):196-203.
[10]Petryk A, Warren JT, Marqués G, et al. Shade is the Drosophila P450 enzyme that mediates the hydroxylation of ecdysone to the steroid insect molting hormone 20-hydroxyecdysone[J]. Proceedings of the National Academy of Sciences, 2003, 100(24):13773- 13778.
[11]Rewitz KF, Rybczynski R, Warren JT, et al. Developmental expression of Manduca shade, the P450 mediating the final step in molting hormone synthesis[J]. Molecular and Cellular Endocrinology, 2006, 247(1):166-174.
[12]Nüsslein-Volhard C, Wieschaus E, Kluding H. Mutations affecting the pattern of the larval cuticle in Drosophila melanogaster[J]. Wilhelm Roux’s Archives of Developmental Biology, 1984, 193(5):267-282.
[13]Nüsslein-Volhard C, Wieschaus E. Mutations affecting segment number and polarity in Drosophila[J]. Nature, 1980, 287(5785):795-801.
[14]Wieschaus E, Nüsslein-Volhard C, Jürgens G. Mutations affecting the pattern of the larval cuticle in Drosophila melanogaster[J]. Wilhelm Roux’s Archives of Developmental Biology, 1984, 193(5):296-307.
[15]Chávez VM, Marqués G, Delbecque JP, et al. The Drosophila disembodied gene controls late embryonic morphogenesis and codes for a cytochrome P450 enzyme that regulates embryonic ecdysone levels[J]. Development, 2000, 127(19):4115-4126.
[16]Warren JT, Petryk A, Marqués G, et al. Molecular and biochemical characterization of two P450 enzymes in the ecdysteroidogenic pathway of Drosophila melanogaster[J]. Proceedings of the National Academy of Sciences, 2002, 99(17):11043-11048.
[17]Warren JT, Petryk A, Marqués G, et al. Phantom encodes the 25-hydroxylase of Drosophila melanogaster and Bombyx mori:a P450 enzyme critical in ecdysone biosynthesis[J]. Insect Biochemistry and Molecular Biology, 2004, 34(9):991-1010.
[18]Hampshire F, Horn D. Structure of crustecdysone, a crustacean moulting hormone[J]. Chemical Communications(London), 1966(2):37-38.
[19]Soumoff C, Skinner DM. Ecdysteroid titers during the molt cycle of the blue crab resemble those of other Crustacea[J]. The Biological Bulletin, 1983, 165(1):321-329.
[20]Lachaise F, Lafont R. Ecdysteroid metabolism in a crab:Carcinus maenas L.[J]. Steroids, 1984, 43(3):243-259.
[21]Watson RD, Spaziani E. Biosynthesis of ecdysteroids from cholesterol by crab Y-organs, and eyestalk suppression of cholesterol uptake and secretory activity, in vitro[J]. General and Comparative Endocrinology, 1985, 59(1):140-148.
[22]沈潔, 朱冬發(fā), 胡則輝, 等. 三疣梭子蟹蛻皮周期的分期[J].水產學報, 2011(10):1481-1487.
[23]Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparative C(T)method[J]. Nature Protocol, 2008, 3(6):1101-1108.
[24]Coon MJ. Cytochrome P450:nature’s most versatile biological catalyst[J]. Annu Rev Pharmacol Toxicol, 2005, 45:1-25.
[25]Werck-Reichhart D, Feyereisen R. Cytochromes P450:a success story[J]. Genome Biol, 2000, 1(6):3001-3009.
[26]艾均文, 董元凌, 孔衛(wèi)青, 等. 家蠶細胞色素 P450 基因CYP6AE2 的分子克隆與序列分析[J]. 蠶業(yè)科學, 2007, 33(3):380-386.
[27]Gonzalez F. The molecular biology of cytochrome P450s[J]. Pharmacological Reviews, 1988, 40(4):243-288.
[28]Rewitz KF, Gilbert LI. Daphnia Halloween genes that encode cytochrome P450s mediating the synthesis of the arthropod molting hormone:evolutionary implications[J]. BMC Evolutionary Biology, 2008, 8(1):60.
[29]Zhou J, Zhang H, Li J, et al. Molecular cloning and expression profile of a Halloween gene encoding CYP307A1 from the seabuckthorn carpenterworm, Holcocerus hippophaecolus[J]. Journal of Insect Science, 2013, 13(1):56.
[30]柳志業(yè), 朱冬發(fā), 謝熙, 等. 三疣梭子蟹CYP302a1基因克隆及其表達分析[J]. 水產學報, 2015(5):628-637.
[31]Chang ES, O’Connor JD. Secretion of alpha-ecdysone by crab Y-organs in vitro[J]. Proceedings of the National Academy of Sciences, 1977, 74(2):615-618.
[32]Ono H, Rewitz KF, Shinoda T, et al. Spook and Spookier code for stage-specific components of the ecdysone biosynthetic pathway in Diptera[J]. Developmental Biology, 2006, 298(2):555-570.
[33]Namiki T, Niwa R, Sakudoh T, et al. Cytochrome P450 CYP307A1/ Spook:a regulator for ecdysone synthesis in insects[J]. Biochemical and Biophysical Research Communications, 2005, 337(1):367-374.
[34]汪春建, 朱冬發(fā), 亓一舟, 等. 三疣梭子蟹蛻皮周期中 MIH基因 mRNA 水平與蛻皮激素濃度變化[J]. 水生生物學報, 2013, 37(1):22-28.
(責任編輯 李楠)
Cloning and Expression Analysis of Spook Gene in Portunus trituberculatus
ZHOU Yanqi ZHU Dong-fa
(The School of Marine Science,Ningbo University,Ningbo 315211)
Ecdysteroids in crustaceans are mainly synthesized and secreted in the Y-organs(YO),and involve in the many essential physiological activities such as molting,reproduction,etc. The cytochrome P450(CYP)307a1 encoded by Spook is the key enzyme in early steps of ecdysteroids biosynthesis. To study the regulatory role of Spook in molting process,the full-length cDNA of gene was cloned and characterized from Portunus trituberculatus(GenBank accession number:KM030021)by reverse transcription PCR(RT-PCR)and RACE,designated as Pt-Spook. The cDNA of Pt-Spook was 2 200 bp in length and contained an opening reading frame(ORF)of 1 563 bp encoding 520 amino acid residues. Multiple alignment revealed that the Pt-Spook contained 5 conserved domains of helix-C,helix-K,helix-I,PERF and heme-binding. The analysis by phylogenetic tree discovered that the Pt-Spook clustered in the same branch with other species,however,Halloween genes were in another branch,indicating that the deduced amino acid sequence was truly the protein sequence of P. trituberculatus Spook. The tissue distribution of Pt-Spook was detected by qPCR,and the results showed that Pt-spook expression was highly specific to YO. During the molt cycle,the expression of Pt-spook gradually increased from post-molt(stage A and B),and reached the maximum in intermolt(stage C),then gradually decreased during pre-molt to the minimum in sub-stage D3and D4. The results suggested that the Pt-spook involved in the regulation of molt cycle in P. trituberculatus.
Portunus trituberculatus;Spook;molt cycle;gene clone;expression level
10.13560/j.cnki.biotech.bull.1985.2016.05.017
2015-12-05
國家自然科學基金項目(41376152,40976098)
周彥琦,女,碩士,研究方向:甲殼動物內分泌學;E-mail:zenlight@foxmail.com
朱冬發(fā),男,教授,研究方向:甲殼動物遺傳育種;E-mail:zhudongfa@nbu.edu.cn
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
