骨骼肌發育調控相關lncRNAs研究進展
2016-07-11 07:42:43占思遠王林杰張紅平
畜牧獸醫學報 2016年4期
占思遠,李 利,王林杰,仲 濤,張紅平
(四川農業大學畜禽遺傳資源發掘與創新利用四川省重點實驗室,成都 611130)
?
骨骼肌發育調控相關lncRNAs研究進展
占思遠,李利,王林杰,仲濤,張紅平*
(四川農業大學畜禽遺傳資源發掘與創新利用四川省重點實驗室,成都 611130)
摘要:長鏈非編碼RNA(Long non-coding RNA,lncRNA)是一類長度大于200 個核苷酸、無蛋白質編碼功能的RNA。相對于研究較多的短鏈非編碼RNA,lncRNA的種類繁多,數量占哺乳動物基因組的絕大部分,功能目前尚不完全清楚。近年來研究發現,lncRNA的功能涉及表觀遺傳、轉錄調控、細胞分化、胚胎發育以及疾病發生等諸多方面。本文總結了lncRNA的分類、作用機制及其在骨骼肌發育調控和家養動物中的研究進展。
關鍵詞:長鏈非編碼RNA;作用機制;骨骼肌發育;調控
骨骼肌的發育是由多種因素參與并受嚴格調控的復雜生物學過程,對哺乳動物的生長發育有著重要的影響。骨骼肌細胞的增殖與分化是骨骼肌發育研究的核心問題,以往的研究主要集中于各類成肌調控因子的時序表達及其對骨骼肌細胞增殖分化的調控機理,包括成肌調節因子家族(Myf5、MyoD、MyoG 和 Myf6)、肌細胞增強因子-2(Myocyte enhancer factor-2,MEF-2)家族、含有配對結構域的轉錄調控因子Pax家族成員Pax3和Pax7等。隨著對哺乳動物中RNA轉錄本日漸深入的研究,哺乳動物基因組上大部分的非編碼RNA被證實具有重要的基礎代謝以及調控功能,例如rRNA、tRNA、miRNA、siRNA以及Piwi蛋白互作RNA(piRNA)等。其中短鏈ncRNA(miRNA、siRNA以及piRNA)被重點關注并被證實在哺乳動物生長、發育和繁殖等生理代謝過程中具有廣泛的調控作用[1]。然而長鏈非編碼RNA(Long non-coding RNA,lncRNA)是一類在真核生物體內主要由RNA聚合酶Ⅱ指導轉錄而成并且長度大于200 nt的ncRNA[2],由于已知功能甚少,其對應的基因組DNA序列甚至被認為是“垃圾”DNA。近年的研究表明,lncRNA能在表觀遺傳、轉錄及轉錄后水平上等多個層次調控基因的表達,并參與了X染色體沉默、基因組印記、胚胎發育、脂肪代謝調控、骨骼肌生長發育以及染色質修飾等重要的調控過程。本文將概述lncRNA的分類、作用機制及其在骨骼肌發育調控方面的研究進展。
1lncRNA的分類與作用機制
lncRNA是一類長度大于200個核苷酸,無蛋白質編碼潛能的RNA。lncRNA通常較長,具有mRNA樣結構和啟動子結構,有些具有poly(A)尾巴,有些沒有poly(A)尾巴,與編碼基因相比,lncRNA表達量更低。許多lncRNA都具有保守的二級結構,一定的剪切形式以及亞細胞定位;同時lncRNA的表達也具有時空特異性。這種保守性和特異性表明它們是具有功能的。根據它們在基因組上相對于蛋白質編碼基因的位置,將其分為5類,包括反義型lncRNA、增強子型lncRNA、發散型lncRNA、內含子型lncRNA和基因間型lncRNA(lincRNA)[3]。依據lncRNA的功能,可以將其劃分為信號分子(Signal molecule)、誘騙分子(Decoy molecule)、引導分子(Guide molecule)以及骨架分子(Scaffold molecule)等4類分子類型[4]。目前相關科研人員已經建立了多個lncRNA的數據庫(表1)。這些數據庫所收錄的lncRNA數據來自于NCBI和已發表的論文,包括人、小鼠、豬、雞和牛等物種的lncRNA信息。
目前的研究表明,lncRNA發揮生物學功能的主要作用機制有基因印記、染色質重塑、組蛋白修飾、轉錄和轉錄后水平調控、細胞凋亡和細胞周期調控等。已有研究證實,H19[5]、X染色體特異性失活轉錄本[6](X inactivation-specific transcription,Xist)等多種lncRNAs參與了基因組印記。lncRNA調控機制中很重要的一部分就是導致染色質重塑,有作者研究了干細胞中lncRNA與組蛋白甲基化的調控關系。研究人員利用定制的lncRNA芯片篩選不同類型樣本(小鼠胚胎干細胞、譜系限制性神經前體細胞和終端分化成纖維細胞)中lncRNA表達差異,分析其表達變化及啟動子區域H3K4和H3K27甲基化程度,發現在不同類型細胞中lncRNA表達模式存在差異。在ES細胞中,lncRNA表達下調導致啟動子區域H3K4甲基化程度變高,從而提出Ezh2介導H3K27甲基化的lncRNA沉默機制[7]。另外,lncRNA還可以在轉錄水平上直接與靶基因結合,促進或抑制靶基因的表達。已經有研究表明,lncRNA不僅可以招募轉錄抑制復合物來沉默基因的表達,還可以作為輔助因子招募轉錄因子,參與基因表達的調控[8]。例如細胞周期蛋白D1(cyclin D1)在人類細胞系中的轉錄抑制機制[9],DNA損傷信號可以誘導cyclin D1基因啟動子區域的lncRNA表達,這些lncRNA可以將TLS(RNA 結合蛋白)招募到cyclin D1基因啟動子區域。隨后,TLS通過抑制CREB結合蛋白和p300的組蛋白乙酰轉移酶活性,進而抑制cyclin D1基因的表達。此外,還有研究表明,lncRNA也具有調控細胞生長的作用,這主要是通過調控細胞凋亡和細胞周期來實現的。生長阻滯特異轉錄物5(Growth arrest-specific transcript 5,Gas5)是哺乳動物細胞生長和凋亡的關鍵性調控因子,它通過模擬糖皮質激素應答元件來結合糖皮質激素受體的 DNA結合域,阻止糖皮質激素受體與其他糖皮質激素應答元件之間的相互作用,從而抑制下游基因的表達,促進細胞凋亡的發生[10]。
2lncRNA調控骨骼肌發育的作用機制
近年來,越來越多的研究證實非編碼RNA(ncRNAs)也是肌肉調控網絡中重要的成員。到目前為止,miRNAs是研究最廣泛的一種非編碼RNA,許多肌肉特異性和非肌肉特異性表達的miRNAs被發現,提示了骨骼肌發育調控的復雜性。而作為非編碼RNA中另一個重要成員,lncRNA廣泛存在于多種生物體內,其功能涉及到細胞生命活動和個體發育與疾病發生的各個方面。目前關于lncRNA的研究主要集中在篩選、鑒定存在于不同物種的不同組織或調控機體生命活動過程的lncRNAs[28-36]。隨著lncRNAs研究的深入,在人和小鼠等模式動物上,lncRNAs的功能機制研究日益增多。因此,越來越多的研究發現骨骼肌的發育過程也會受到各種lncRNAs的調控(表2)。
2.1lncRNA通過競爭性結合miRNA調控骨骼肌的生長發育
M.Cesana等[37]的研究證實了一種在成肌細胞分化過程中特異性表達的長鏈非編碼RNA-linc-MD1可發揮ceRNA(Competing endogenous RNA,競爭性內源RNA)活性,通過miR-133 和miR-135實現對肌肉特異性轉錄調控因子MAML1和MEF2C的表達調控。在未分化的肌細胞中,miR-133和miR-135可分別結合MAML-1和MEF2C基因的mRNA,并抑制其表達。linc-MD-1含有這兩個miRNA的結合位點,分化過程中高表達的linc-MD-1競爭性結合細胞中的miR-133和miR-135,降低了這兩種miRNA的濃度,從而解除了對肌肉分化相關蛋白質因子的抑制作用,促進了細胞的分化。另外,linc-MD1的表達抑制可導致成肌細胞分化延遲,與杜氏肌營養不良癥的發病密切相關[37]。近年來,關于肺腺癌轉移相關轉錄本Malat1(Metastasis-associated lung adenocarcinoma transcript 1)在腫瘤方面的研究已經很清楚,但最近的研究發現Malat1還能調節肌細胞的生長。X.Han等[38]在C2C12細胞中研究發現,Malat1通過競爭性的結合miR-133,調控其靶基因肌細胞分化關鍵因子SRF(Serum response factor)的表達,從而影響肌細胞的分化。研究表明,Malat1被沉默時,與Malat1結合的miR-133被釋放出來作用于SRF的3′UTR區,導致SRF的表達抑制從而抑制了肌細胞的分化。
表1lncRNA相關數據庫
Table 1lncRNA databases

數據庫Database網址Website參考文獻ReferenceLNCipediahttp://www.lncipedia.org[11]ncRNAdatabasehttp://biobases.ibch.poznan.pl/ncRNA[12]NONCODEv4.0http://www.noncode.org[13]lncRNAdbhttp://www.lncrnadb.org[14]NREDhttp://jsm-research.imb.uq.edu.au/NRED[15]ncFANshttp://www.noncode.org/ncFANs[16]fRNAdbhttp://www.ncrna.org/frnadb[17]LncRNADiseasehttp://cmbi.bjmu.edu.cn/lncrnadisease[18]ChIPBasehttp://deepbase.sysu.edu.cn/chipbase[19]DIANA-LncBasehttp://www.microrna.gr/LncBase[20]starBasev2.0http://starbase.sysu.edu.cn[21]FunctionallncRNADatabasehttp://www.valadkhanlab.org/database[22]lnCeDBhttp://gyanxet-beta.com/lncedb[23]ALDBhttp://res.xaut.edu.cn/aldb/index.jsp[24]LncRNA2Targethttp://www.lncrna2target.org[25]LncRNASNPhttp://bioinfo.life.hust.edu.cn/lncRNASNP[26]LncRNAWikihttp://lncrna.big.ac.cn/index.php/Main_Page[27]
2.2lncRNA通過調節靶mRNA的表達和降解影響骨骼肌的生長發育
Malat1是一種高度保守的約8.7 kb的非編碼轉錄本,在癌細胞中大量表達,同時也是癌細胞轉移的重要信號分子[39]。有試驗研究發現重組了肌肉抑制素(Myostatin)的小鼠腓腸肌中,Malat1的表達量顯著下降[40];同時也發現在小鼠成肌細胞向肌管分化的過程中,Malat1表達量上升,敲除了Malat1的成肌細胞的增殖被抑制。這些結果表明,Malat1作為一種新的myostatin的下游靶標,在肌細胞的生成過程中起著重要的調控作用。J.Wang等[41]研究表明,鼠類含有SINE(Short interspersed elements)的非編碼長鏈RNA能夠控制肌肉生成。比如1/2-sbsRNA通過與RNA結合蛋白STAU1或STAU2的結合引起SMD(Staufen1-mediated mRNA decay)對mRNA的降解從而正向調控小鼠C2C12細胞的成肌分化過程。
2.3lncRNA通過順式作用影響骨骼肌的成肌分化
L.Lu等[42]研究發現了一種肌肉相關lincRNA—Yam-1,它能夠被轉錄因子Yin Yang 1(YY1)正向調節,同時Yam-1也是肌細胞生成的抑制因子,它通過順式調控miR-715,影響其靶基因Wnt7b的表達,從而達到抑制成肌細胞分化的目的。另外有研究也發現了lncRNA的非順式調控作用模式,例如MUNC(MyoD Upstream Non-Coding),通常被認為是一種DRR(Distal Regulatory Region)增強子RNA(DRReRNA),在骨骼肌組織中特異性地表達[43]。試驗結果表明,MUNC的作用機制并不像傳統的eRNA那樣,通過刺激鄰近位點的MyoD基因的表達來達到順式調控的目的,而更像一種前肌源lncRNA,通過直接或間接地作用于多個啟動子區域增加生肌調節因子基因的表達,從而促進骨骼肌細胞的成肌分化過程[43]。
2.4lncRNA作為印記基因在骨骼肌生長發育中發揮重要作用
B.K.Dey等[44]研究發現,作為參與基因組印記的典型lncRNA—H19,它在胚胎組織中大量轉錄,呈母系表達,但出生后表達被顯著抑制,唯獨在骨骼肌中仍維持顯著的轉錄水平,進一步研究發現,H19敲低的小鼠成肌細胞和H19敲除小鼠的骨骼肌衛星細胞中,成肌分化顯著降低。H19的外顯子1編碼兩個保守miRNAs前體,分別為miR-675-3p和miR-675-5p,H19缺陷小鼠在受傷后表現為畸形的骨骼肌再生,當外源引入miR-675-3p和miR-675-5p后可以被調整,它們能直接靶定和下調抗分化轉錄因子Smad和DNA復制起始因子Cdc6的表達,從而促進骨骼肌的成肌分化和肌肉再生。另有研究結果表明,let-7的過度表達會引起過早的肌肉分化,而H19可以充當分子海綿來調節let-7的活性,進而阻止肌肉分化的過早發育[5]。
2.5lncRNA調控肌源性前體細胞的分化決定
M.Ballarino等[45]研究新發現了一種調控肌肉生成的長鏈非編碼RNA—lnc-31,它和miR-31來自相同的核內miR-31前體,但產生途徑卻獨立于miR-31。小鼠lnc-31與它的人類同源序列hsa-lnc-31被證實在維持細胞增殖和抑制分化中扮演了重要的作用。同樣地,小鼠lnc-31和hsa-lnc-31被發現在mdx(DMD動物模型)小鼠肌肉和Duchenne型肌營養不良(DMD)的病人成肌細胞中呈高表達,伴隨著大量的肌源性前體細胞的激活。這些結果表明,lnc-31在調控肌源性前體細胞的分化決定方面起著關鍵性的作用。
2.6lncRNA編碼微肽調控骨骼肌細胞的生理學過程
最近,有研究者發現了一個保守微肽,命名為肌調素(Myoregulin,MLN),它由骨骼肌特異性RNA編碼,這個RNA被注釋為假定的長鏈非編碼RNA[46]。MLN與肌漿網受磷蛋白(Phospholamban,PLN)和肌脂蛋白(Sarcolipin,SLN)結構和功能相似,并抑制心肌肌漿網Ca2+-ATP酶(SERCA),SERCA通過調節鈣離子攝取到肌質網(SR)控制肌肉松弛。MLN直接與SERCA相互作用并阻礙鈣吸收到SR。PLN和SLN在小鼠心和慢速骨骼肌中表達,MLN積極的表達于所有骨骼肌。在小鼠中的MLN基因缺失,增強鈣離子在骨骼肌中的處置能力并提高運動能力[46]。這些發現表明了MLN是骨骼肌生理的一個重要調節因子,最大的亮點是在許多被注釋為非編碼的RNA可能編碼“微肽(Micropeptides)”。
2.7lncRNA通過調節DNA甲基化影響骨骼肌成肌分化
L.Wang等[47]在骨骼肌成肌細胞中鑒定出一個lncRNA—Dum(多能發育相關基因2(Dppa2)上游結合肌肉lncRNA),體內外研究發現,Dum的表達量在肌細胞生成過程中呈動態變化,它能被上游的MyoD轉錄誘導從而影響成肌細胞分化;進一步研究表明,Dum能夠促進成肌細胞的分化和受損肌肉的再生,作用機制在于在染色體上Dum基因座與Dppa2基因的啟動子區域是并排的,Dum通過招募多種DNA甲基化轉移酶—Dnmts(Dnmt1,Dnmt3a和Dnmt3b)到Dppa2的啟動子區域,從而導致CpG位點的高甲基化和Dppa2基因的沉默。這些研究結果表明,MyoD-Dum-Dppa2調節軸在成肌分化和肌肉再生中發揮了重要的功能。
2.8lncRNA調控骨骼肌發育的其他機制
目前雖然有很多lncRNAs被發現,但是大部分的功能尚未清楚,比如人類的ε/β lncRNAs(Men ε/β lncRNAs),它有ε和β兩種亞型,在患有I型多發性內分泌瘤的病人多個組織中表達,包括肌肉組織[48],在C2C12細胞分化過程中,Men ε/β lncRNAs的表達量顯著上升,但是具體的生物學機制仍不清楚;另有研究發現,類固醇受體RNA激活因子(Steroid receptor RNA activator,SRA)RNA具有兩種特異的存在形式,分別是非編碼和編碼的RNA(Yielding SRA ncRNA and protein SRAP),其中SRA ncRNA作為MyoD的共同激活因子,在肌細胞分化過程中高度表達[49-50]。最近有研究還發現了一種lncRNA—細胞核因子非編碼抑制因子(Non-coding repressor of NFAT,NRON),它本身具有組織特異性的表達模式,在胎盤、肌肉和淋巴組織中高度表達[51]。以上的這些研究都表明,在骨骼肌的生長發育過程中,長鏈非編碼RNA起著重要的調控作用,并且作用機制具有多元化特點,需要更深入的功能機制研究。
表2骨骼肌發育調控相關lncRNAs
Table 2lncRNAs in the regulation of skeletal muscle development
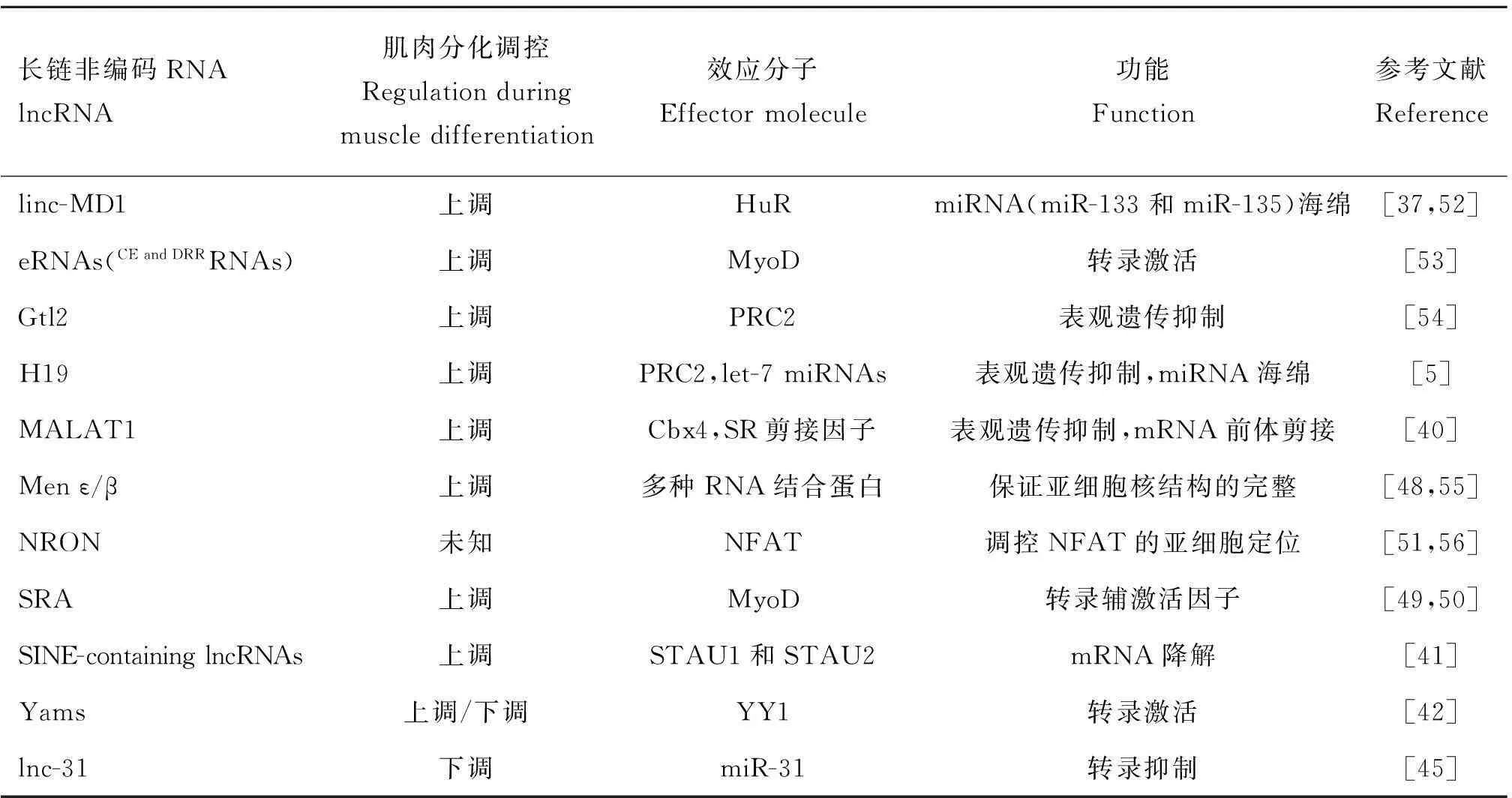
長鏈非編碼RNAlncRNA肌肉分化調控Regulationduringmuscledifferentiation效應分子Effectormolecule功能Function參考文獻Referencelinc-MD1上調HuRmiRNA(miR-133和miR-135)海綿[37,52]eRNAs(CEandDRRRNAs)上調MyoD轉錄激活[53]Gtl2上調PRC2表觀遺傳抑制[54]H19上調PRC2,let-7miRNAs表觀遺傳抑制,miRNA海綿[5]MALAT1上調Cbx4,SR剪接因子表觀遺傳抑制,mRNA前體剪接[40]Menε/β上調多種RNA結合蛋白保證亞細胞核結構的完整[48,55]NRON未知NFAT調控NFAT的亞細胞定位[51,56]SRA上調MyoD轉錄輔激活因子[49,50]SINE-containinglncRNAs上調STAU1和STAU2mRNA降解[41]Yams上調/下調YY1轉錄激活[42]lnc-31下調miR-31轉錄抑制[45]
3家養動物骨骼肌相關lncRNAs的研究進展
目前,國內外lncRNA的研究還處于初步階段,lncRNA的鑒定以及功能研究主要集中于人以及模式動物中(如小鼠、果蠅和線蟲等),其他物種的lncRNA研究相對滯后,特別是在家養動物領域。至今只有較少的文章對雞、牛和豬的lncRNA進行了研究報道,其他家養動物中lncRNA的相關研究仍然是空白。
T.Li等[36]在2012年報道了雞骨骼肌中lncRNA的鑒定研究結果。該研究組利用RNA-seq 技術對3個發育階段的雞骨骼肌進行了轉錄組測序,并利用自行開發的生物信息學流程鑒定出281個雞的lncRNAs。并在進一步分析中發現,相比蛋白編碼基因,雞lncRNA的保守性更低。其后,J.Luo[57]對馬萊克氏病抵抗型和易感型的雞CD4 細胞系進行了轉錄組測序和比對,共鑒定出2 626個雞的lncRNAs,其中1 177個特異性表達于馬萊克氏病抵抗型細胞,另有1 048個特異性表達于馬萊克氏病易感型細胞中。
W.Huang等[58]對公共的牛特異性表達序列標簽(Expressed sequence tags,EST)數據進行了重新組裝,并利用支持向量機算法對RNA轉錄本進行了編碼蛋白質潛能篩選,共鑒定出405個基因間lncRNAs(lincRNAs)。這些牛lncRNAs普遍具有組織特異性表達特征,它們的GC含量比隨機選擇的基因間序列的要高,但比蛋白質編碼基因的低,并且它們在哺乳動物之間中度保守。C.Billerey等[35]利用RNA-seq技術對9頭利木贊牛的胸腰最長肌進行了轉錄組測序,共鑒定出了418個lincRNAs,并對其基因組特征進行了分析。此后2014年12月,J.Caballero等[59]利用轉錄組芯片技術對奶牛早期胚胎發育不同階段細胞進行測序分析,發現了3個從卵母細胞到8細胞期差異表達的lncRNAs,進一步研究表明,這些lncRNAs是通過調節靶基因mRNA的翻譯水平,從而調控奶牛早期胚胎的發育過程。
Z.Y.Zhou等[60]利用已知的豬RNA-seq信息和ESTs數據,鑒定出6 621個lincRNAs,發現其中一些lincRNA具有同線性和序列保守性,如linc-sscg2561,在基因組上與其鄰近的是與情緒行為相關聯的Dnmt3a(DNA methyltransferase 3a)基因。研究還發現linc-sscg2561和Dnmt3a在家豬和野豬的額葉皮質中表達差異顯著,提示linc-sscg2561在家豬的馴化過程中可能發揮了重要的作用。
4展望
盡管長度小于50個堿基的非編碼 RNA(如 miRNA 和 piRNA 等)的研究已取得突破性進展,但長度大于200個堿基的非編碼RNA還有很多的功能和作用機制尚未闡明。lncRNA不僅數量巨大,且其作用機制具有多樣性和復雜性,而隨著lncRNA研究技術和手段的不斷發展,lncRNA的功能機制研究也會越來越多元化。通過對骨骼肌發育相關lncRNAs的研究,將進一步深化對哺乳動物肌肉生成分子機制的認識,并可能為肌肉發育相關疾病的治療提供新的思路和應對策略。
參考文獻(References):
[1]STEFANI G,SLACK F J.Small non-coding RNAs in animal development[J].NatRevMolCellBiol,2008,9(3):219-230.
[2]WILUSZ J E,SUNWOO H,SPECTOR D L.Long noncoding RNAs:functional surprises from the RNA world[J].GenesDev,2009,23(13):1494-1504.
[3]KNOLL M,LODISH H F,SUN L.Long non-coding RNAs as regulators of the endocrine system[J].NatRevEndocrinol,2015,11(3):151-160.
[4]WANG K C,CHANG H Y.Molecular mechanisms of long noncoding RNAs[J].MolCell,2011,43(6):904-914.
[5]KALLEN A N,ZHOU X B,XU J,et al.The imprinted H19 lncRNA antagonizes let-7 microRNAs[J].MolCell,2013,52(1):101-112.
[6]ENGREITZ J M,PANDYA-JONES A,MCDONEL P,et al.The Xist lncRNA exploits three-dimensional genome architecture to spread across the X chromosome[J].Science,2013,341(6147):1237973.
[7]WU S C,KALLIN E M,ZHANG Y.Role of H3K27 methylation in the regulation of lncRNA expression[J].CellRes,2010,20(10):1109-1116.
[8]CHEN L L,CARMICHAEL G G.Decoding the function of nuclear long non-coding RNAs[J].CurrOpinCellBiol,2010,22(3):357-364.
[9]WANG X,ARAI S,SONG X,et al.Induced ncRNAs allosterically modify RNA-binding proteins in cis to inhibit transcription[J].Nature,2008,454(7200):126-130.
[10]KINO T,HURT D E,ICHIJO T,et al.Noncoding RNA gas5 is a growth arrest-and starvation-associated repressor of the glucocorticoid receptor[J].SciSignal,2010,3(107):ra8.
[11]VOLDERS P J,HELSENS K,WANG X,et al.LNCipedia:a database for annotated human lncRNA transcript sequences and structures[J].NucleicAcidsRes,2013,41(Database issue):D246-D251.
[12]ERDMANN V A,SZYMANSKI M,HOCHBERG A,et al.Non-coding,mRNA-like RNAs database Y2K[J].NucleicAcidsRes,2000,28(1):197-200.
[13]XIE C,YUAN J,LI H,et al.NONCODEv4:exploring the world of long non-coding RNA genes[J].NucleicAcidsRes,2014,42(Database issue):D98-D103.
[14]AMARAL P P,CLARK M B,GASCOIGNE D K,et al.lncRNAdb:a reference database for long noncoding RNAs[J].NucleicAcidsRes,2011,39(Database issue):D146-D151.
[15]DINGER M E,PANG K C,MERCER T R,et al.NRED:a database of long noncoding RNA expression[J].NucleicAcidsRes,2009,37(Database issue):D122-D126.
[16]LIAO Q,XIAO H,BU D,et al.ncFANs:a web server for functional annotation of long non-coding RNAs[J].NucleicAcidsRes,2011,39(Web Server issue):W118-W124.
[17]MITUYAMA T,YAMADA K,HATTORI E,et al.The functional RNA database 3.0:databases to support mining and annotation of functional RNAs[J].NucleicAcidsRes,2009,37(Database issue):D89-D92.
[18]CHEN G,WANG Z,WANG D,et al.LncRNADisease:a database for long-non-coding RNA-associated diseases[J].NucleicAcidsRes,2013,41(Database issue):D983-D986.
[19]YANG J H,LI J H,JIANG S,et al.ChIPBase:a database for decoding the transcriptional regulation of long non-coding RNA and microRNA genes from ChIP-Seq data[J].NucleicAcidsRes,2013,41(Database issue):D177-D187.
[20]PARASKEVOPOULOU M D,GEORGAKILAS G,KOSTOULAS N,et al.DIANA-LncBase:experimentally verified and computationally predicted microRNA targets on long non-coding RNAs[J].NucleicAcidsRes,2013,41(Database issue):D239-D245.
[21]LI J H,LIU S,ZHOU H,et al.StarBase v2.0:decoding miRNA-ceRNA,miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data[J].NucleicAcidsRes,2014,42(Database issue):D92-D97.
[22]NIAZI F,VALADKHAN S.Computational analysis of functional long noncoding RNAs reveals lack of peptide-coding capacity and parallels with 3′UTRs[J].RNA,2012,18(4):825-843.
[23]DAS S,GHOSAL S,SEN R,et al.lnCeDB:database of human long noncoding RNA acting as competing endogenous RNA[J].PLoSOne,2014,9(6):e98965.
[24]LI A,ZHANG J,ZHOU Z,et al.ALDB:a domestic-animal long noncoding RNA database[J].PLoSOne,2015,10(4):e0124003.
[25]JIANG Q,WANG J,WU X,et al.LncRNA2Target:a database for differentially expressed genes after lncRNA knockdown or overexpression[J].NucleicAcidsRes,2015,43(Database issue):D193-D196.
[26]GONG J,LIU W,ZHANG J,et al.lncRNASNP:a database of SNPs in lncRNAs and their potential functions in human and mouse[J].NucleicAcidsRes,2015,43(Database issue):D181-D186.
[27]MA L,LI A,ZOU D,et al.LncRNAWiki:harnessing community knowledge in collaborative curation of human long non-coding RNAs[J].NucleicAcidsRes,2015,43(Database issue):D187-D192.
[28]ZHU J G,SHEN Y H,LIU H L,et al.Long noncoding RNAs expression profile of the developing mouse heart[J].JCellBiochem,2014,115(5):910-918.
[29]MATKOVICH S J,EDWARDS J R,GROSSENHEIDER T C,et al.Epigenetic coordination of embryonic heart transcription by dynamically regulated long noncoding RNAs[J].ProcNatlAcadSciUSA,2014,111(33):12264-12269.
[30]MALOUF G G,ZHANG J,YUAN Y,et al.Characterization of long non-coding RNA transcriptome in clear-cell renal cell carcinoma by next-generation deep sequencing[J].MolOncol,2015,9(1):32-43.
[31]LV J,HUANG Z,LIU H,et al.Identification and characterization of long intergenic non-coding RNAs related to mouse liver development[J].MolGenetGenomics,2014,289(6):1225-1235.
[32]LIPOVICH L,TARCA A L,CAI J,et al.Developmental changes in the transcriptome of human cerebral cortex tissue:long noncoding RNA transcripts[J].CerebCortex,2014,24(6):1451-1459.
[33]LIANG M,LI W,TIAN H,et al.Sequential expression of long noncoding RNA as mRNA gene expression in specific stages of mouse spermatogenesis[J].SciRep,2014,4:5966.
[34]LV J,CUI W,LIU H,et al.Identification and characterization of long non-coding RNAs related to mouse embryonic brain development from available transcriptomic data[J].PLoSOne,2013,8(8):e71152.
[35]BILLEREY C,BOUSSAHA M,ESQUERRé D,et al.Identification of large intergenic non-coding RNAs in bovine muscle using next-generation transcriptomic sequencing[J].BMCGenomics,2014,15:499.
[36]LI T,WANG S,WU R,et al.Identification of long non-protein coding RNAs in chicken skeletal muscle using next generation sequencing[J].Genomics,2012,99(5):292-298.
[37]CESANA M,CACCHIARELLI D,LEGNINI I,et al.A long noncoding RNA controls muscle differentiation by functioning as a competing endogenous RNA[J].Cell,2011,147(2):358-369.
[38]HAN X,YANG F,CAO H,et al.Malat1 regulates serum response factor through miR-133 as a competing endogenous RNA in myogenesis[J].FASEBJ,2015,29(7):3054-3064.
[39]SCHMIDT L H,SPIEKER T,KOSCHMIEDER S,et al.The long noncoding MALAT-1 RNA indicates a poor prognosis in non-small cell lung cancer and induces migration and tumor growth[J].JThoracOncol,2011,6(12):1984-1992.
[40]WATTS R,JOHNSEN V L,SHEARER J,et al.Myostatin-induced inhibition of the long noncoding RNA Malat1 is associated with decreased myogenesis[J].AmJPhysiolCellPhysiol,2013,304(10):C995-C1001.
[41]WANG J,GONG C,MAQUAT L E.Control of myogenesis by rodent SINE-containing lncRNAs[J].GenesDev,2013,27(7):793-804.
[42]LU L,SUN K,CHEN X,et al.Genome-wide survey by ChIP-seq reveals YY1 regulation of lincRNAs in skeletal myogenesis[J].EMBOJ,2013,32(19):2575-2588.
[43]MUELLER A C,CICHEWICZ M A,DEY B K,et al.MUNC,a long noncoding RNA that facilitates the function of MyoD in skeletal myogenesis[J].MolCellBiol,2015,35(3):498-513.
[44]DEY B K,PFEIFER K,DUTTA A.The H19 long noncoding RNA gives rise to microRNAs miR-675-3p and miR-675-5p to promote skeletal muscle differentiation and regeneration[J].GenesDev,2014,28(5):491-501.
[45]BALLARINO M,CAZZELLA V,D’ANDREA D,et al.Novel long noncoding RNAs (lncRNAs) in myogenesis:a miR-31 overlapping lncRNA transcript controls myoblast differentiation[J].MolCellBiol,2015,35(4):728-736.
[46]ANDERSON D M,ANDERSON K M,CHANG C L,et al.A micropeptide encoded by a putative long noncoding RNA regulates muscle performance[J].Cell,2015,160(4):595-606.
[47]WANG L,ZHAO Y,BAO X,et al.LncRNA Dum interacts with Dnmts to regulate Dppa2 expression during myogenic differentiation and muscle regeneration[J].CellRes,2015,25(3):335-350.
[48]SUNWOO H,DINGER M E,WILUSZ J E,et al.MEN epsilon/beta nuclear-retained non-coding RNAs are up-regulated upon muscle differentiation and are essential components of paraspeckles[J].GenomeRes,2009,19(3):347-359.
[49]HUBé F,VELASCO G,ROLLIN J,et al.Steroid receptor RNA activator protein binds to and counteracts SRA RNA-mediated activation of MyoD and muscle differentiation[J].NucleicAcidsRes,2011,39(2):513-525.
[50]CARETTI G,SCHITZ R L,DILWORTH F J,et al.The RNA helicases p68/p72 and the noncoding RNA SRA are coregulators of MyoD and skeletal muscle differentiation[J].DevCell,2006,11(4):547-560.
[51]WILLINGHAM A T,ORTH A P,BATALOV S,et al.A strategy for probing the function of noncoding RNAs finds a repressor of NFAT[J].Science,2005,309(5740):1570-1573.
[52]LEGNINI I,MORLANDO M,MANGIAVACCHI A,et al.A feedforward regulatory loop between HuR and the long noncoding RNA linc-MD1 controls early phases of myogenesis[J].MolCell,2014,53(3):506-514.
[53]MOUSAVI K,ZARE H,DELL’ORSO S,et al.eRNAs promote transcription by establishing chromatin accessibility at defined genomic loci[J].MolCell,2013,51(5):606-617.
[54]SCHUSTER-GOSSLER K,BILINSKI P,SADO T,et al.The mouse Gtl2 gene is differentially expressed during embryonic development,encodes multiple alternatively spliced transcripts,and may act as an RNA[J].DevDyn,1998,212(2):214-228.
[55]CLEMSON C M,HUTCHINSON J N,SARA S A,et al.An architectural role for a nuclear noncoding RNA:NEAT1 RNA is essential for the structure of paraspeckles[J].MolCell,2009,33(6):717-726.
[56]SHARMA S,FINDLAY G M,BANDUKWALA H S,et al.Dephosphorylation of the nuclear factor of activated T cells (NFAT) transcription factor is regulated by an RNA-protein scaffold complex[J].ProcNatlAcadSciUSA,2011,108(28):11381-11386.
[57]LUO J.Identification of specific long noncoding RNA profiles in chicken with different susceptibility to Marek’s disease[C].in Plant and Animal Genome XXI Conference.2013.Plant and Animal Genome.
[58]HUANG W,LONG N,KHATIB H.Genome-wide identification and initial characterization of bovine long non-coding RNAs from EST data[J].AnimGenet,2012,43(6):674-682.
[59]CABALLERO J,GILBERT I,FOURNIER E,et al.Exploring the function of long non-coding RNA in the development of bovine early embryos[J].ReprodFertilDev,2014,27(1):40-52.
[60]ZHOU Z Y,LI A M,ADEOLA A C,et al.Genome-wide identification of long intergenic noncoding RNA genes and their potential association with domestication in pigs[J].GenomeBiolEvol,2014,6(6):1387-1392.
(編輯郭云雁)
Research Progress of Long Noncoding RNAs in the Regulation of Skeletal Muscle Development
ZHAN Si-yuan,LI Li,WANG Lin-jie,ZHONG Tao,ZHANG Hong-ping*
(KeyLaboratoryofFarmAnimalGeneticResourcesExplorationandInnovationofSichuanProvince,SichuanAgriculturalUniversity,Chengdu611130,China)
Abstract:Long non-coding RNAs(lncRNAs)are a novel class of RNAs,which are longer than 200 nucleotides and have no protein-coding potential.In comparison with short noncoding RNAs which was studied extensively,a variety of long noncoding RNAs account for the majority of the transcripts in mammalian genome and the functions of lncRNAs are little understood.The recent studies have indicated that lncRNAs’ function involved in epigenetic modification,transcriptional regulation,cell differentiation,embryonic development and disease occurrence.In this review,we summarized the classification and molecular mechanisms of lncRNA,and research advances in the regulation of skeletal muscle development and domestic animal.
Key words:long non-coding RNAs;mechanism of action;skeletal muscle development;regulation
doi:10.11843/j.issn.0366-6964.2016.04.001
收稿日期:2015-06-24
基金項目:四川省科技支撐計劃(2014NZ0077;2015NZ0112)
作者簡介:占思遠(1987-),男,湖北紅安人,博士生,主要從事動物遺傳育種與繁殖研究,E-mail:siyuan_zhan@163.com *通信作者:張紅平,博士,教授,主要從事動物遺傳育種與繁殖研究,E-mail:zhp@sicau.edu.cn
中圖分類號:S827.2
文獻標志碼:A
文章編號:0366-6964(2016)04-0637-08
猜你喜歡
中國醫藥導報(2017年21期)2017-09-04 04:22:04
中國醫藥導報(2017年8期)2017-05-11 21:45:43
科技創新導報(2016年28期)2017-03-14 11:32:29
中國醫藥導報(2016年31期)2017-02-28 14:47:45
商業經濟研究(2016年22期)2016-12-27 10:51:08
中國實用醫藥(2016年29期)2016-12-26 08:53:55
農家科技下旬刊(2016年9期)2016-12-15 08:33:09
中國中藥雜志(2016年20期)2016-11-19 12:24:52
人間(2016年26期)2016-11-03 17:51:57
中國實用醫藥(2016年24期)2016-10-17 04:45:48
