一次離心運(yùn)動(dòng)結(jié)合針刺對大鼠骨骼肌線粒體動(dòng)力學(xué)蛋白的影響
2016-07-18 11:58:22于瀅
哈爾濱體育學(xué)院學(xué)報(bào) 2016年4期
于 瀅
?
一次離心運(yùn)動(dòng)結(jié)合針刺對大鼠骨骼肌線粒體動(dòng)力學(xué)蛋白的影響
于 瀅
目的:觀察一次離心運(yùn)動(dòng)對骨骼肌線粒體動(dòng)力學(xué)相關(guān)蛋白的變化,并探究針刺干預(yù)對其影響。方法:將SD大鼠分為安靜對照、針刺、一次離心運(yùn)動(dòng)和運(yùn)動(dòng)針刺四大組。運(yùn)動(dòng)方式采取下坡跑,坡度-16°,速度16m/min,時(shí)間90min;針刺方法小腿三頭肌縱向從遠(yuǎn)端斜刺,進(jìn)針角度約 30°,進(jìn)針止于小腿三頭肌肌腹處,留針2min。各組又按運(yùn)動(dòng)后時(shí)相點(diǎn)分為 0h、6h、12h、24h、48h和 72h 小組,大鼠在對應(yīng)時(shí)間點(diǎn)取比目魚肌進(jìn)行檢測。Western Blot法檢測針刺干預(yù)及一次運(yùn)動(dòng)后不同時(shí)程后線粒體動(dòng)力學(xué)相關(guān)蛋白Mfn2、Drp1的蛋白表達(dá)。結(jié)果:單純針刺后Mfn2蛋白表達(dá)上調(diào),Drp1在12h出現(xiàn)顯著上調(diào)。單純運(yùn)動(dòng)組Drp1在0h出現(xiàn)顯著降低。與運(yùn)動(dòng)組相比較,運(yùn)動(dòng)針刺后Mfn2在0h、24h有顯著變化。Drp1蛋白表達(dá)在0h、12h有顯著變化。結(jié)論:一次離心運(yùn)動(dòng)早期使骨骼肌線粒體出現(xiàn)分裂抑制。針刺可以改善一次離心運(yùn)動(dòng)導(dǎo)致的骨骼肌線粒體動(dòng)力學(xué)狀態(tài),進(jìn)而影響線粒體分布與功能。
離心運(yùn)動(dòng);骨骼肌;線粒體;針刺
線粒體是一種始終處于高度運(yùn)動(dòng)狀態(tài)的細(xì)胞器,頻繁地出現(xiàn)分裂與融合的動(dòng)態(tài)過程。線粒體動(dòng)力學(xué)是指細(xì)胞中的線粒體不斷進(jìn)行分裂與融合的一種動(dòng)態(tài)變化,包括分裂與融合兩個(gè)過程,受線粒體分裂和融合蛋白調(diào)控。只有線粒體分裂與融合達(dá)到一定程度上的平衡,才能維持正常的線粒體形態(tài)結(jié)構(gòu)與功能[1]。正常情況下,線粒體融合與分裂的速度相當(dāng),其數(shù)目與形態(tài)較為穩(wěn)定,保持在一個(gè)動(dòng)態(tài)的平衡之中。但是,如果線粒體的分裂增多,就會(huì)引起線粒體的片段化,與此同時(shí)線粒體會(huì)出現(xiàn)損傷,可能表現(xiàn)出線粒體膜電位的降低。而如果線粒體的融合增多,其在細(xì)胞內(nèi)的網(wǎng)絡(luò)化程度就會(huì)加強(qiáng),同時(shí)可以改善線粒體的功能,對細(xì)胞可能會(huì)起到一定保護(hù)作用。線粒體動(dòng)力學(xué)變化是在一組進(jìn)化保守的線粒體融合和分裂蛋白的控制下完成的[2]。線粒體融合的核心裝置是融合蛋白1和2(Mitofusin1/2,Mfn1/Mfn2)以及視神經(jīng)萎縮蛋白1(Optic atrophy 1,OPA1)。其中Mfn1/Mfn2位于線粒體外膜和中膜,而OPA1位于線粒體內(nèi)膜,在促進(jìn)線粒體內(nèi)膜融合的同時(shí)也使線粒體的嵴進(jìn)行了重構(gòu)。線粒體分裂的核心裝置由發(fā)動(dòng)蛋白相關(guān)GTP酶1(Dynamin-related protein 1,Drp1)、分裂蛋白1(Fission1,F(xiàn)is1)、線粒體分裂因子(mitochondrial fission factor,MFF)等蛋白組成[3]。
骨骼肌細(xì)胞的功能維持極度依賴線粒體正常分布與功能,而運(yùn)動(dòng)可能通過干擾線粒體動(dòng)力學(xué)平衡引發(fā)骨骼肌能量代謝障礙。線粒體動(dòng)力學(xué)失衡可能是運(yùn)動(dòng)導(dǎo)致骨骼肌運(yùn)動(dòng)能力下降亞細(xì)胞水平的原因之一。施冰等[4]觀察到大鼠急性心肌梗死后,梗死周邊區(qū)域心肌組織中Mfn2和Drp1mRNA下降,并在心肌梗死后2周內(nèi)表達(dá)呈逐漸下降的趨勢,認(rèn)為線粒體融合和分裂異常導(dǎo)致的能量代謝障礙,可能是心肌梗死后心室重塑及心力衰竭的重要機(jī)制之一。本研究擬通過實(shí)驗(yàn)分析針刺和一次離心運(yùn)動(dòng)后骨骼肌線粒體動(dòng)力學(xué)蛋白Mfn2、Drp1的變化特點(diǎn),探討針刺和一次離心運(yùn)動(dòng)對大鼠骨骼肌線粒體動(dòng)力學(xué)的影響。
1 實(shí)驗(yàn)材料與方法
1.1 動(dòng)物分組
本實(shí)驗(yàn)的實(shí)驗(yàn)動(dòng)物為雄性SPF級Spraque-Dawley(SD)大鼠114只,體重為215.79±9.38g,購于北京維通利華實(shí)驗(yàn)動(dòng)物技術(shù)有限公司,許可證編號(hào):SCXK(京)2012-0001。實(shí)驗(yàn)大鼠飼養(yǎng)和訓(xùn)練均在北京體育大學(xué)科學(xué)研究中心的動(dòng)物房內(nèi)進(jìn)行。大鼠分籠飼養(yǎng),自由飲食飲水,室溫22±2℃,相對濕度30%~60%,12h光照/12h熄燈模擬日晝交替。大鼠適應(yīng)環(huán)境3天,之后進(jìn)行后續(xù)實(shí)驗(yàn)(見表1)。
表1 實(shí)驗(yàn)動(dòng)物分組情況一覽表
1.2 實(shí)驗(yàn)動(dòng)物運(yùn)動(dòng)和針刺方案
運(yùn)動(dòng)形式采用大負(fù)荷損傷運(yùn)動(dòng)模型。具體運(yùn)動(dòng)方案:跑臺(tái)坡度-16°,跑速為16m/min,運(yùn)動(dòng)持續(xù)時(shí)間為90min。針刺方案:沿大鼠的小腿三頭肌縱向,從遠(yuǎn)端斜刺進(jìn)入到小腿三頭肌肌腹,留針時(shí)間為2min。
1.3 測試樣本的收集與保存
大鼠稱重后,用5%的水合氯醛(0.8ml/100g)進(jìn)行腹腔麻醉。將血抽取干凈,迅速分離比目魚肌。取出來的比目魚肌立即用錫紙包好,做好標(biāo)記,立刻投入到液氮罐中暫時(shí)保存,待取材完成后將比目魚肌標(biāo)本轉(zhuǎn)移至-80℃冰箱保存,備用。
1.4 蛋白質(zhì)免疫印跡方法測定比目魚肌蛋白含量
取100mg凍存的骨骼肌組織,使用液氮在研缽中將組織研磨成粉末,收集粉末,將其加入到1ml含蛋白酶抑制劑和PMSF的裂解液中。冰水混合物中靜置30min,4℃,12 000g離心10min,取上清液,分裝,-80℃保存。電泳前以4:1比例加入5×上樣緩沖液,98℃,煮沸10min。蛋白含量測定采用BCA 測試方法,按試劑盒說明進(jìn)行。配制Tris-甘氨酸SDS聚丙烯酰胺凝膠,分離膠10%、,積層膠5%。電泳條件:積層膠70V,40min;分離膠110V,通過預(yù)染蛋白marker來確定電泳停止時(shí)間。采用濕法進(jìn)行蛋白質(zhì)轉(zhuǎn)膜,根據(jù)PVDF膜上預(yù)染Marker轉(zhuǎn)移情況,判斷蛋白的轉(zhuǎn)移情況。將目的條帶剪下,放入自封袋中,倒入適量5%BSA封閉液,室溫,平緩搖動(dòng),孵育2h;封閉PVDF膜上存在的非特異性的結(jié)合位點(diǎn)。封閉結(jié)束后,用濾紙吸干目的條帶上的封閉液,將目的條帶放入已經(jīng)加了一抗(Mfn2為1:2 000,Drp1為1:4000,abcum)和封閉液的潔凈自封袋中,放入冰箱4℃,過夜;二抗稀釋比例為1:4 000(中山金橋)。化學(xué)發(fā)光成像在Image Lab 4.1軟件凝膠成像系統(tǒng)中進(jìn)行。經(jīng)Image Lab拍攝出的照片,導(dǎo)出后使用Gelpro軟件對目的蛋白質(zhì)進(jìn)行光密度相對定量分析,以GAPDH為內(nèi)參。
1.5 數(shù)據(jù)統(tǒng)計(jì)方法
所有數(shù)據(jù)用SPSS17.0統(tǒng)計(jì)軟件包處理,數(shù)據(jù)均以平均值±標(biāo)準(zhǔn)差(X±SD)的形式表示。實(shí)驗(yàn)數(shù)據(jù)采用雙因素方差分析(Two-way ANOVA),對處理因素(安靜對照、單純針刺、一次離心運(yùn)動(dòng)和運(yùn)動(dòng)針刺)和不同的取材時(shí)間因素(運(yùn)動(dòng)后即刻、6h、12h、24h、48h和72h)的主效應(yīng)以及兩者的交互作用進(jìn)行分析。各組之間的單獨(dú)效應(yīng)統(tǒng)計(jì)方法使用ANOVA單因素方差分析,方差齊性時(shí)用LSD方法,方差不齊性時(shí)用Tamhane′s方法。顯著性水平定為P<0.05,非常顯著性水平定為P<0.01。
2 實(shí)驗(yàn)結(jié)果
2.1 一次離心運(yùn)動(dòng)和針刺后不同時(shí)相Mfn2蛋白表達(dá)的變化
雙因素分析結(jié)果顯示(見表2),不同的干預(yù)方法及干預(yù)后時(shí)間兩因素以及二者的交互作用顯著(P<0.05),即不同干預(yù)手段對不同時(shí)間點(diǎn)的Mfn2的影響不同。
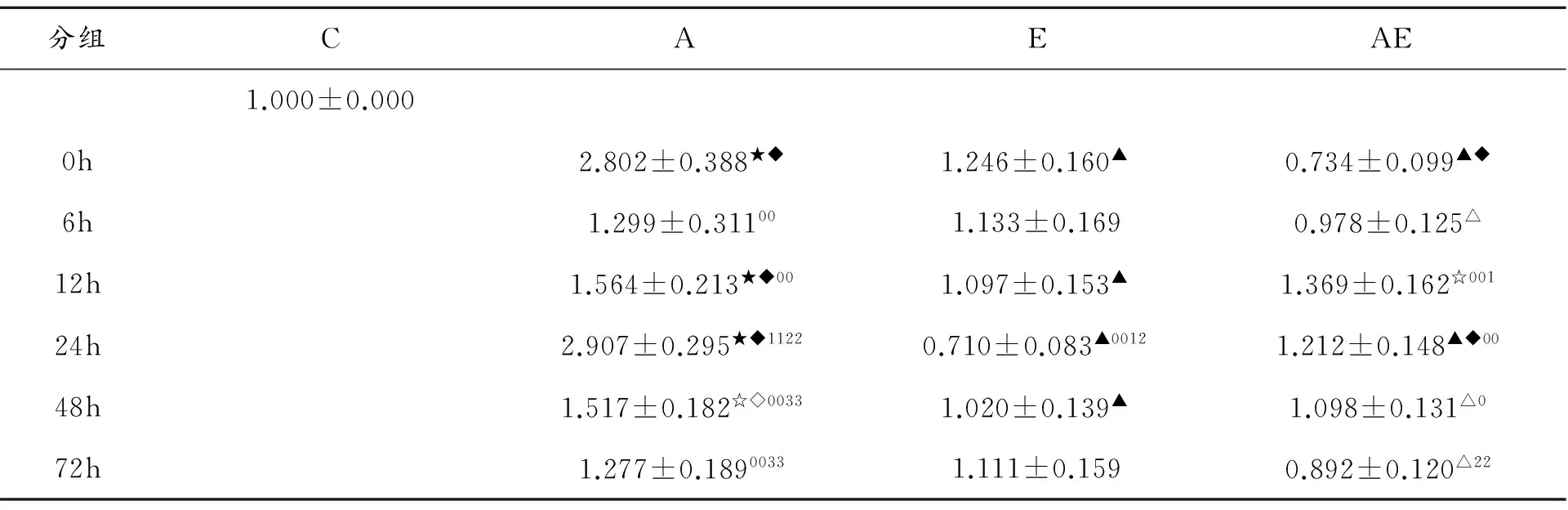
表2 一次離心運(yùn)動(dòng)和針刺骨骼肌Mfn2蛋白表達(dá)變化 (% C group)
注:組內(nèi),與C相比,☆P<0.05;組內(nèi),與C相比,★P<0.01;同一時(shí)間點(diǎn),與A相比,△P<0.05;同一時(shí)間點(diǎn),與A相比,▲P<0.01;同一時(shí)間點(diǎn),與E相比,◇P<0.05;同一時(shí)間點(diǎn),與E相比,◆P<0.01;組內(nèi),與0h相比,0P<0.05;組內(nèi),與0h相比,00P<0.01;組內(nèi),與6h相比,1P<0.05;組內(nèi),與6h相比,11P<0.01;組內(nèi),與12h相比,2P<0.05;組內(nèi),與12h相比,22P<0.01;組內(nèi),與24h相比,3P<0.05;組內(nèi),與24h相比,33P<0.01;組內(nèi),與48h相比,4P<0.05;組內(nèi),與48h相比,44P<0.05
采用單因素方差分析各組的單獨(dú)效應(yīng)。實(shí)驗(yàn)結(jié)果顯示(表1、圖1),單純針刺組在干預(yù)后Mfn2蛋白的表達(dá)呈現(xiàn)升高趨勢,與安靜對照組相比,0h、12h、24h和48h組Mfn2蛋白的表達(dá)差異具有顯著性。6h、12h、48h和72h組與0h組相比,Mfn2蛋白的表達(dá)差異具有顯著性;24h與6h組Mfn2蛋白的表達(dá)相比較,差異具有顯著性;24h組與12h組相比較差異顯著,48h和72h與24h差異顯著。單純運(yùn)動(dòng)組在干預(yù)后Mfn2蛋白的表達(dá)呈現(xiàn)圍繞對照組的水平上下波動(dòng)的趨勢。24h與0h、6h、12h組Mfn2蛋白的表達(dá)相比較,差異具有顯著性。
運(yùn)動(dòng)針刺組Mfn2蛋白的表達(dá)呈現(xiàn)先升高后降低的趨勢,與安靜組相比較, 12h組Mfn2蛋白的表達(dá)明顯升高,差異具有顯著性。12h和48h組Mfn2蛋白與0h組相比較差異具有顯著性;12h、24h、48h與0h組相比較差異具有顯著性;12h與6h組Mfn2蛋白的表達(dá)差異具有顯著性;72h與12h組Mfn2蛋白的表達(dá)差異具有顯著性。
圖1 一次離心運(yùn)動(dòng)和針刺骨骼肌Mfn2蛋白表達(dá)變化圖
與針刺組相比,運(yùn)動(dòng)組在0h、12h、24h和48h組Mfn2蛋白表達(dá)均顯著降低降低,差異具有顯著性。運(yùn)動(dòng)針刺組在0h、6h、24h、48h、72h組骨骼肌Mfn2蛋白表達(dá)顯著低于針刺組。
與運(yùn)動(dòng)組相比,針刺組在0h、12h、24h和48h組Mfn2蛋白表達(dá)差異具有顯著性。運(yùn)動(dòng)針刺組在0h和24h組Mfn2蛋白表達(dá)差異具有顯著性。
2.2 一次離心運(yùn)動(dòng)和針刺后不同時(shí)相Drp1蛋白表達(dá)的變化
雙因素分析結(jié)果顯示,不同的干預(yù)方法對蛋白表達(dá)影響明顯,而時(shí)間因素不明顯,二者的交互作用顯著(P<0.05),即不同干預(yù)手段對不同時(shí)間點(diǎn)的Drp1的影響不同。采用單因素方差分析各組的單獨(dú)效應(yīng)。實(shí)驗(yàn)結(jié)果顯示(表3、圖2)單純針刺組在干預(yù)后Drp1蛋白的表達(dá)呈現(xiàn)先降低后升高趨勢,與安靜對照組相比,12h組Drp1蛋白的表達(dá)升高明顯,差異具有顯著性。12h、48h和72h組與0h組相比,Drp1蛋白的表達(dá)差異具有顯著性;12h與6h組Drp1蛋白的表達(dá)相比較,差異具有顯著性。單純運(yùn)動(dòng)組在干預(yù)后Drp1蛋白的表達(dá)呈現(xiàn)波動(dòng)的變化的趨勢。運(yùn)動(dòng)后即刻Drp1蛋白的表達(dá)即出現(xiàn)降低,與安靜對照組相比,Drp1蛋白的表達(dá)降低明顯,差異具有顯著性;12h和72h組與0h組相比,Drp1蛋白的表達(dá)差異具有顯著性;12h、72h組與6h組Drp1蛋白的表達(dá)相比較,差異具有顯著性。運(yùn)動(dòng)針刺后即刻Drp1蛋白的表達(dá)沒有顯著變化,0h與12h組相比,Drp1蛋白的表達(dá)差異具有顯著性。
與針刺組相比,運(yùn)動(dòng)組48h組Drp1蛋白表達(dá)差異具有顯著性。運(yùn)動(dòng)針刺組12h和48h組Drp1蛋白表達(dá)均降低,差異具有顯著性。與運(yùn)動(dòng)組相比,針刺組48h組Drp1蛋白表達(dá)升高,具有顯著性差異;運(yùn)動(dòng)針刺組在0h、12h組差異顯著。

表3 一次離心運(yùn)動(dòng)和針刺骨骼肌Drp1蛋白表達(dá)變化 (% C group)
注:組內(nèi),與C相比,☆P<0.05;組內(nèi),與C相比,★P<0.01;同一時(shí)間點(diǎn),與A相比,△P<0.05;同一時(shí)間點(diǎn),與A相比,▲P<0.01;同一時(shí)間點(diǎn),與E相比,◇P<0.05;同一時(shí)間點(diǎn),與E相比, ◆P<0.01;組內(nèi),與0h相比,0P<0.05;組內(nèi),與0h相比,00P<0.01;組內(nèi),與6h相比,1P<0.05;組內(nèi),與6h相比,11P<0.01
圖2 一次離心運(yùn)動(dòng)和針刺骨骼肌Drp1蛋白表達(dá)變化圖
3 分析與討論
3.1 一次離心運(yùn)動(dòng)對骨骼肌線粒體動(dòng)力學(xué)相關(guān)蛋白的影響
在運(yùn)動(dòng)干預(yù)條件下,線粒體動(dòng)力學(xué)相關(guān)蛋白會(huì)受到影響,線粒體的功能也會(huì)發(fā)生變化,大部分研究結(jié)果認(rèn)為,線粒體融合分裂參與了細(xì)胞對能量需求的快速應(yīng)答。通過融合,線粒體體積增大,是線粒體運(yùn)動(dòng)適應(yīng)的一種早期機(jī)制。尤其是耐力訓(xùn)練,可以增加線粒體的大小、體積,增強(qiáng)其磷酸化能力[5]。
運(yùn)動(dòng)對骨骼肌線粒體動(dòng)力學(xué)會(huì)產(chǎn)生影響。劉慧君[6]研究表明,小鼠一次中等強(qiáng)度負(fù)荷跑臺(tái)運(yùn)動(dòng)過程中,骨骼肌線粒體Mfn2蛋白運(yùn)動(dòng)60min、90min、120min組基因及蛋白表達(dá)較安靜組顯著降低,線粒體分裂蛋白Drp1運(yùn)動(dòng)60min、90min組mRNA及蛋白表達(dá)較安靜組顯著升高。且急性運(yùn)動(dòng)過程中,Mfn2基因蛋白表達(dá)顯著下調(diào),Drp1基因蛋白表達(dá)上調(diào)。這提示小鼠一次120min中等強(qiáng)度負(fù)荷跑臺(tái)運(yùn)動(dòng)過程中,骨骼肌線粒體融合受抑制趨于分裂。
漆正堂[7]報(bào)道,骨骼肌線粒體在對不同訓(xùn)練方式(后肢懸吊組、耐力訓(xùn)練組、間歇性沖刺訓(xùn)練組)的適應(yīng)過程中,線粒體相關(guān)指標(biāo)會(huì)發(fā)生變化。耐力訓(xùn)練可上調(diào)Mfn1 mRNA表達(dá);間歇性沖刺訓(xùn)練后Mfn2 mRNA轉(zhuǎn)錄下調(diào),蛋白表達(dá)上調(diào);懸吊組Mfn1 mRNA表達(dá)顯著高于安靜組。間歇性沖刺訓(xùn)練后,Drpl mRNA轉(zhuǎn)錄上調(diào),蛋白表達(dá)下調(diào)。懸吊組Drp1蛋白表達(dá)顯著低于安靜組。懸吊組和沖刺組線粒體Mfn2/Drpl比值顯著高于安靜組,其中沖刺組還顯著高于耐力組。間歇性沖刺訓(xùn)練能誘導(dǎo)線粒體融合加強(qiáng)而抑制分裂,可能協(xié)同促進(jìn)線粒體的融合,間歇性沖刺訓(xùn)練誘導(dǎo)線粒體融合活性顯著高于耐力訓(xùn)練。間歇性沖刺訓(xùn)練對線粒體系統(tǒng)的影響比耐力訓(xùn)練大得多。從提高線粒體機(jī)能和防止線粒體疾病的角度出發(fā),認(rèn)為間歇性沖刺訓(xùn)練是一種省時(shí)的運(yùn)動(dòng)形式。
Cartoni等[8]報(bào)道,運(yùn)動(dòng)訓(xùn)練24h后,人體骨骼肌細(xì)胞內(nèi)Mfn1、Mfn2的mRNA表達(dá)顯著上升,并推測Mfn2基因表達(dá)可能受雌激素相關(guān)受體(ERRα)的調(diào)控。另一項(xiàng)研究表明,骨骼肌中hFisl mRNA表達(dá)在急性運(yùn)動(dòng)中明顯增加,而運(yùn)動(dòng)后骨骼肌hFisl mRNA逐漸減少,且在運(yùn)動(dòng)后24h末未恢復(fù)到安靜水平,認(rèn)為hFisl基因的動(dòng)態(tài)表達(dá)是線粒體對細(xì)胞能量需求急劇增長的一種快速應(yīng)答反應(yīng)[9]。
此外,線粒體在年齡增加導(dǎo)致的肌肉質(zhì)量和力量下降過程中發(fā)揮了重要作用[10]。劉子泉發(fā)現(xiàn),增齡過程中骨骼肌線粒體融合蛋白Mfn1、Mfn2 mRNA和蛋白表達(dá)減少,而經(jīng)12周中等強(qiáng)度耐力訓(xùn)練后,Mfn1、Mfn2的mRNA和蛋白表達(dá)均增加;增齡和耐力訓(xùn)練后,分裂蛋白Drp1、Fis1mRNA和蛋白表達(dá)增加。這提示線粒體經(jīng)訓(xùn)練后建立了適度水平的動(dòng)力學(xué)平衡,使融合分裂更加活躍;有氧耐力訓(xùn)練能提高線粒體融合分裂的動(dòng)態(tài)平衡。骨骼肌是重要的運(yùn)動(dòng)器官,通過適當(dāng)?shù)倪\(yùn)動(dòng)干預(yù)可以改善其細(xì)胞內(nèi)的線粒體融合分裂。
本研究發(fā)現(xiàn),一次離心運(yùn)動(dòng)后即刻骨骼肌Mfn2和Drp1蛋白的表達(dá)變化呈現(xiàn)波動(dòng)變化,Mfn2與安靜組無顯著性差異。在運(yùn)動(dòng)后即刻Mfn2蛋白變化不明顯,但Drp1蛋白表達(dá)出現(xiàn)顯著降低,線粒體的動(dòng)力學(xué)表現(xiàn)為分裂受抑制,這可能是骨骼肌內(nèi)線粒體動(dòng)力學(xué)對一次離心運(yùn)動(dòng)的一種應(yīng)激性變化,線粒體分裂相關(guān)蛋白表達(dá)降低,導(dǎo)致線粒體的分裂受到抑制,鏡下可觀察到巨大線粒體。說明一次離心運(yùn)動(dòng)即刻對線粒體動(dòng)力學(xué)的影響是向抑制分裂變化的,這種變化導(dǎo)致線粒體不能被及時(shí)的分裂成適于移動(dòng)的大小,因此不利于線粒體在細(xì)胞中的移動(dòng),尤其是在經(jīng)過一次離心的運(yùn)動(dòng),損傷部位的能量需求較多,而線粒體不能通過分裂,快速的移動(dòng)到需要部位,不利于損傷的恢復(fù),并且可能加劇損傷,因此線粒體動(dòng)力學(xué)改變可能是導(dǎo)致骨骼肌延遲性損傷的原因之一。
3.2 針刺對骨骼肌線粒體動(dòng)力學(xué)相關(guān)蛋白的影響
針刺作為一種治療方法,已有很多研究證明其對一些疾病的治療具有一定的療效,而在治療機(jī)理上也有一些針對于對線粒體線粒體動(dòng)力學(xué)方面的研究報(bào)道。
羅磊研究發(fā)現(xiàn),在AD發(fā)病過程中,大鼠海馬神經(jīng)元線粒體融合相關(guān)蛋白Opa1的水平明顯下降,而線粒體分裂相關(guān)蛋白Fis1的水平明顯升高。而“益腎調(diào)督”針灸法能夠明顯的上調(diào)Opa1,下調(diào)Fis1的水平,認(rèn)為針灸法能夠有效的改善海馬神經(jīng)元線粒體分裂融合的失衡狀態(tài)[11]。
何勛通過研究AD海馬神經(jīng)元線粒體Mfn1、Mfn2的明顯減少,經(jīng)針灸治療后,其Mfn1、Mfn2明顯提高,認(rèn)為針灸治療能提高海馬神經(jīng)元線粒體Mfn1、Mfn2的表達(dá),改善AD 發(fā)病過程中海馬神經(jīng)元線粒體動(dòng)力學(xué)的異常,從而達(dá)到防治AD的目的[12]。
針刺對骨骼肌線粒體的動(dòng)力學(xué)蛋白研究較少。宋福[13]從線粒體融合方面研究針刺對骨骼肌萎縮治療的分子機(jī)制,針刺小鼠雙側(cè)下肢的環(huán)跳、足三里和承山3個(gè)穴位,結(jié)果發(fā)現(xiàn)針刺組小鼠的脛骨前肌中Mfn1和Opa1含量顯著升高,腓腸肌中Mfn1和Opa1含量沒有顯著性變化,認(rèn)為單純的針刺治療很可能通過提高線粒體的融合使線粒體功能得到提高,進(jìn)而為機(jī)體提供更多的能量。
本實(shí)驗(yàn)觀察了針刺對正常骨骼肌Mfn2蛋白和Drp1蛋白的表達(dá),發(fā)現(xiàn)針刺后即刻、12h、24h和48h組,Mfn2蛋白顯著升高;Drp1蛋白表達(dá)僅在12h出現(xiàn)顯著升高。說明針刺可以提高線粒體的融合,對線粒體的動(dòng)力學(xué)平衡產(chǎn)生了一定的影響,通過提高M(jìn)fn2蛋白和Drp1蛋白的表達(dá),使線粒體的融合與分裂建立一個(gè)新的平衡點(diǎn),有利于線粒體功能的提高。
3.3 一次離心運(yùn)動(dòng)后針刺干預(yù)對骨骼肌線粒體動(dòng)力學(xué)相關(guān)蛋白的影響
有關(guān)針刺和運(yùn)動(dòng)結(jié)合對骨骼肌線粒體動(dòng)力學(xué)蛋白的報(bào)道尚不多見,本研究試圖在這方面進(jìn)行一些嘗試性的研究和探索。通過結(jié)果分析發(fā)現(xiàn),與運(yùn)動(dòng)組相比較,運(yùn)動(dòng)針刺后即刻Drp1蛋白的表達(dá)在0h組顯著升高,同時(shí)發(fā)現(xiàn)Mfn2蛋白的表達(dá)有顯著性降低,此時(shí),線粒體的融合與分裂相比較于運(yùn)動(dòng)組,建立起一個(gè)新的動(dòng)態(tài)平衡,分裂表現(xiàn)活躍,有利于運(yùn)動(dòng)后能量的轉(zhuǎn)運(yùn)。與運(yùn)動(dòng)組相比較,運(yùn)動(dòng)針刺后即刻Drp1蛋白的表達(dá)在12h組顯著下調(diào),且在12h和24hMfn2蛋白表達(dá)顯著升高,線粒體的動(dòng)力學(xué)表現(xiàn)以融合占優(yōu)勢,這可能是針刺對一次離心運(yùn)動(dòng)大鼠骨骼肌內(nèi)線粒體產(chǎn)生刺激效應(yīng),且這種刺激效應(yīng)在不同時(shí)間點(diǎn)表現(xiàn)不同,主要通過提高M(jìn)fn2蛋白表達(dá),有效的改善了由于一次離心運(yùn)動(dòng)導(dǎo)致的線粒體動(dòng)力學(xué)失衡狀態(tài)。一次離心運(yùn)動(dòng)后即刻給予針刺,可使線粒體的融合減少,而12h和24h又可以提高融合過程,有利于線粒體的網(wǎng)絡(luò)化分布,從以上分析可以看出,針刺對線粒體的動(dòng)力學(xué)的影響也是動(dòng)態(tài)變化的,這種變化與運(yùn)動(dòng)組相比較,使線粒體的分裂與融合活躍,能夠更好地滿足機(jī)體在運(yùn)動(dòng)后恢復(fù)的能量需求,而這種作用可能主要是通過提高M(jìn)fn2表達(dá)來實(shí)現(xiàn)的。
4 結(jié) 論
(1)一次離心運(yùn)動(dòng)后即刻骨骼肌線粒體出現(xiàn)分裂抑制。
(2)單純針刺可以通過上調(diào)Mfn2蛋白表達(dá)使線粒體趨于融合。
(3)針刺可以使一次離心運(yùn)動(dòng)后的線粒體動(dòng)力學(xué)狀態(tài)得到改善,有利于機(jī)體在恢復(fù)過程的能量供應(yīng)。
[1] Knott A B, Bossy-Wetzel E. Impairing the mitochondrial fission and fusion balance: a new mechanism of neurodegeneration[J].Annals of the New York Academy of Sciences,2008, 147(1): 283-292.
[2] Sang-Bing O, Andrew R. H, Derek J ,et al. Mitochondrial Dynamics in Cardiovascular Health and Disease[J].Antioxidants redox signaling,2013,19(4):400-414.
[3] José MG .Mitochondrial Dynamics in Health and Disease[J]. Mitochondria and Their Role in Cardiovascular Disease,2013:329-342.
[4] 施 冰,冬 蘭,尹秋生,等.大鼠心肌梗死后心肌組織中線粒體融合蛋白2基因和線粒體分裂蛋白mRNA表達(dá)水平的變化[J].中國臨床保健雜志,2012,15(4):392-394.
[5] Mark W. Molecular Signals and Skeletal Muscle Adaptation to Exercise[J].International Journal of Applied Exercise Physiology,2013,2(1).
[6] 劉慧君.過氧化氫、Ca2+對骨骼肌線粒體移動(dòng)和動(dòng)態(tài)變化的調(diào)節(jié)[D].北京:中國人民解放軍軍事醫(yī)學(xué)科學(xué)院,2010.
[7] 漆正堂.骨骼肌線粒體對不同訓(xùn)練方式的適應(yīng)及其基因應(yīng)答機(jī)制的研究[D].上海:華東師范大學(xué),2009.
[8] Cartoni R,Leger B,Hock MB,et al. Mitofusins 1/2 and ERR alpha expression are increased in human skeletal muscle after physical exercise[J].J Physiol-London,2005,567(1):349-358.
[9] 孫衛(wèi)東,丁 虎,劉曉然,等. 骨骼肌線粒體對細(xì)胞能量需求的快速應(yīng)答:mfn1/2與fis1基因在急性運(yùn)動(dòng)中的動(dòng)態(tài)表達(dá)[J].中國運(yùn)動(dòng)醫(yī)學(xué)雜志,2008,27(5):544-550.
[10] Courtney M P,Darcy L J,Eric R .Skeletal Muscle Mitochondria and Aging:A Review[J].Journal of Aging Research,2012:1-20.
[11] 羅 磊. “益腎調(diào)督” 法對 AD 模型大鼠神經(jīng)元軸突線粒體損傷的影響及針灸作用機(jī)制研究[D].武漢:湖北中醫(yī)藥大學(xué),2014.
[12] 何 勛. “益腎調(diào)督” 法針灸對阿爾茨海默病模型大鼠海馬神經(jīng)元線粒體分裂/融合蛋白的影響[D].武漢:湖北中醫(yī)藥大學(xué),2014.
[13] 宋 福. 針刺對治療脛骨前肌和腓腸肌肌萎縮分子機(jī)制的初步研究[D].天津:天津體育學(xué)院,2014.
An Eccentric Exercise Combined with Acupuncture on Mitochondria Dynamics Protein Expression on Rat Skeletal Muscle
YU Ying
Objective:The skeletal muscle mitochondria dynamics of proteins were observed after aneccentric exercise and acupuncture intervention.Methods:SD rats were divided into 4 groups: control,acupuncture,eccentric exercise, acupuncture and exercise. Movement way to run downhill,slope - 16 °, the speed is 16 m/min, 90 min time; Acupuncture method of longitudinal from the distal leg triceps stop at leg triceps muscle belly, angle is about 30°,retaining needle 2 minutes.After the termination of experiments,each group was divided into 0h,12h,24h,48h and 72h group,and the soleus was detected in the corresponding time points. Western Blot was used to detect mitochondrial dynamics protein ( Mfn2, Drp1) expression after exercise and acupuncture intervention.Results:Acupuncture group Mfn2 protein expression increased. Drp1 protein expression increased in 12 hours.Exercise group Drp1 protein expression decreased in 0 hours.Acupuncture and exercise group Mfn2 and Drp1 protein expression had significant differences compared with exercise group.Conclusion:
Mitochondrial fusion was inhibited after an eccentric exercise, and acupuncture could ameliorate dynamics state, and affected the mitochondria distribution and function.
Eccentric exercise; Skeletal muscle; Mitochondria; Acupuncture
2016-06-08;
2016-07-26
國家自然基金項(xiàng)目《運(yùn)動(dòng)影響骨骼肌微管調(diào)控線粒體結(jié)構(gòu)和功能的機(jī)制研究》(編號(hào):31271277);中央高校基本科研業(yè)務(wù)費(fèi)專項(xiàng)基金資助課題(編號(hào):2015ZD004,2016ZX020)。
于 瀅(1979-),女,講師,博士,研究方向?yàn)檫\(yùn)動(dòng)與骨骼肌損傷。
哈爾濱體育學(xué)院 運(yùn)動(dòng)科學(xué)與健康系,黑龍江 哈爾濱 150008
G804
A
1008-2808(2016)04-0015-06
猜你喜歡
中學(xué)生數(shù)理化·中考版(2022年8期)2022-06-14 06:55:24
今日農(nóng)業(yè)(2021年19期)2022-01-12 06:16:36
中老年保健(2021年11期)2021-08-22 03:15:44
中國民間療法(2021年1期)2021-04-20 02:30:34
基層中醫(yī)藥(2021年7期)2021-03-26 18:00:27
中學(xué)生數(shù)理化(高中版.高考數(shù)學(xué))(2021年1期)2021-03-19 08:28:38
無線電工程(2020年11期)2020-10-29 01:25:46
現(xiàn)代出版(2020年3期)2020-06-20 07:10:34
中醫(yī)研究(2014年8期)2014-03-11 20:29:23
中醫(yī)研究(2014年6期)2014-03-11 20:29:02
