瞬時受體電位通道蛋白的跨膜片段分析
2016-07-22 02:44:01蘇秋香張麗艷汲坤張改改張利華
中國醫科大學學報 2016年7期
蘇秋香,張麗艷,汲坤,張改改,張利華
(沈陽醫學院基礎醫學院1.機能實驗學教研室;2.病理生理教研室,沈陽 110034)
?
瞬時受體電位通道蛋白的跨膜片段分析
蘇秋香1,張麗艷2,汲坤2,張改改2,張利華2
(沈陽醫學院基礎醫學院1.機能實驗學教研室;2.病理生理教研室,沈陽 110034)
摘要目的分析瞬時受體電位(TRP)通道的膜拓撲結構。方法采用糖基化的方法,對傳統TRP通道TRPC5的各疏水片段進行膜整合分析。結果TRPC5通道中,S4~S8片段按順序分別以Ncyt/Cexo和Nexo/Ccyt的方式插入到膜中,C末端位于細胞內。而S1~S3片段可能存在2種膜整合方式:一種是S1和S3整合到膜上,S2位于細胞外;另一種是混合模式,S1和S3分別位于細胞內,其N末端均位于細胞內。結論TRPC5通道的S4~S8為跨膜片段,S1~S3片段需進一步實驗驗證。
關鍵詞瞬時受體電位通道;膜拓撲結構;跨膜片段分析
網絡出版地址
瞬時受體電位(transient receptor potential,TRP)通道是一類非選擇性陽離子通道,位于細胞膜上,最早發現于果蠅的視覺系統[1]。在哺乳動物中已發現28種TRP通道亞型,分屬于7個亞家族,包括TRPC、TRPV、TRPM、TRPA、TRPP、TRPML和TRPN,每個亞家族又包含若干通道[2]。TRP通道蛋白分布廣泛,其調節機制各異且功能多樣,參與到多種生理過程中[3]。哺乳動物TRPC通道又被分為TRPC1、TRPC2、TRPC4/5、TRPC3/6/7四個亞型[4]。由于TRP通道的亞型較多,功能復雜,直至最近才將TRP結構與功能的關系基本闡明[5]。所有亞家族結構上均含6個跨膜拓撲結構,其氨基酸(N)末端和羧基(C)末端均位于胞內,由第5及第6跨膜片段共同構成非選擇性陽離子孔道。TRPC通道N末端由3~4個錨蛋白重復序列、1個卷曲螺旋和1個小窩蛋白結合區組成;C末端包含TRP標志基序(EWKFAR)。雖然TRP通道的基本結構已經提出,但具體的跨膜疏水片段以及拓撲結構形成機制并未闡明。VANNIER等[6]于1998年報道,TRPC3的第1個疏水片段位于細胞內,而其余的7個疏水片段按順序整合到膜上,成為跨膜片段。而DOHKE等[7]于2004年報道,TRPC1的第3個疏水片段位于細胞內,而其余的7個疏水片段按順序整合到膜上,成為跨膜片段。離子通道屬于膜蛋白,其結構的闡明對于了解離子通道的功能非常有幫助。拓撲結構分析常被用來研究膜蛋白的結構,研究膜蛋白的各個跨膜片斷在線粒體蛋白合成的同時是如何整合到內質網膜上的,并最終形成什么樣的跨膜結構[8]。目前,最常用的方法是糖基化和蛋白酶K水解。本研究采用體外蛋白轉錄、翻譯和轉運系統以及糖基化方法分析了TRP家族中比較有代表性的TRPC5通道的膜拓撲結構,為闡明TRP通道蛋白的結構與功能提供數據。
1 材料與方法
1.1質粒構建
1.1.1G?loop導入質粒制作:利用PCR方法擴出編碼TRPC5中含疏水片段S1~S8的DNA片段(V292?D636),同時在DNA片段的5′端導入NcoⅠ酶切位點,3′端導入XhoⅠ酶切位點,利用NcoⅠ和XhoⅠ酶切位點把該DNA片段亞克隆到pCITE?2a載體(美國Novagen公司)中。然后利用點變異方法,分別在每個疏水片段之間的連接中根據有效糖基化的12?14規則導入EcoⅠ酶切位點,利用酶切的方法,把來自人帶3蛋白中含有一個糖基化受體位點的G?loop分別插入到各連接中的EcoⅠ酶切位點上。
1.1.2截短蛋白質粒制作:我們利用PCR方法分別擴出編碼TRPC5中疏水片段S1、S1~S2、S1~S3、S1~S4、S1~S5、S1~S6、S1~S7、S1~S8的DNA片段,然后利用酶切的方法,把這些DNA片段融合到攜帶有PLg報告基因的pCITE?2a載體中,PLg報告基因中含有糖基化位點。
1.2體外蛋白轉錄、翻譯及轉運實驗
利用狗胰腺粗面內質網小泡和兔網織紅細胞抽出液模擬體內蛋白轉錄、翻譯及轉運實驗。首先進行mRNA合成,所有質粒經過ScaⅠ酶切線性化,在標準條件下使用T7 RNA多聚酶進行體外轉錄,37℃,1 h。然后把獲得的mRNA在含有或不含有狗胰腺粗面內質網小泡的兔網織紅細胞抽出液系統中進行體外蛋白合成和轉運,37℃,1 h。對在含有粗面內質網條件下合成的蛋白產物進一步進行蛋白水解試驗,蛋白水解酶采用蛋白酶K。我們用[35S]蛋氨酸標記蛋白,用SDS?PAGE電泳分離蛋白產物,最后進行同位素自顯影。
1.3膜拓撲結構分析
在未添加粗面內質網小泡的條件下,膜蛋白經體外合成后出現一條蛋白帶,這是未被糖基化的蛋白帶。因為糖基化反應僅發生在內質網膜腔內,而且一個糖基化受體位點由于接受一個高甘露糖基寡聚糖,分子量會增加2 500。因此,在粗面內質網存在下,如果出現1條分子量比未被糖基化的蛋白帶多出約3 000的蛋白帶,就說明該蛋白帶被糖基化了,同時也說明該蛋白含有糖基化受體位點的多肽區域位于內質網腔內(細胞外)。在粗面內質網存在下,獲得的蛋白經過蛋白酶K處理后,位于內質網腔外(細胞內)的多肽會被蛋白酶K消化降解,蛋白帶縮短或消失。根據導入的糖基化受體位點是否被糖基化以及合成的蛋白是否被蛋白酶K消化降解,可以推測出蛋白的多肽鏈是位于內質網腔內(細胞外)還是位于內質網腔外(細胞內),從而推測出膜蛋白各疏水區域是否是跨膜片段,即推測出膜蛋白的拓撲結構。
2 結果
2.1TRPC5跨膜疏水片段預測
利用Kyte?Doolittle方法對TRPC5氨基酸序列進行疏水性分析(圖1)。S1(328~350)、S2(359~384)、S3(402~418)、S4(437~456)、S5(476~497)、S6(508~532)、S7(567~587)、S8(598~624)這8個區域疏水性較高,被認為可能成為跨膜片段。
2.2使用N和C末端切除蛋白進行TRPC5各片段的膜整合分析
N末端(氨基酸末端)常常會影響蛋白的體外表達量,所以在膜蛋白的拓撲結構分析中常常使用N 和C末端切除蛋白進行體外蛋白的表達及轉運實驗。如圖2A,先構建N和C末端切除的TRPC5質粒TRPC5(V292?D636),然后在每2個可能的跨膜疏水片段(S1~S8)的中間連接(loop)中分別導入來自于人帶3蛋白的含有一個糖基化位點的G?loop,質粒構建完成后,體外合成mRNA,在粗面內質網和蛋白酶K分別存在的條件下,進行蛋白合成和轉運,最后進行SDS?PAGE電泳實驗。
SDS?PAGE結果(圖2B)顯示,在未添加粗面內質網小泡的條件下,TRPC5(V292?D636)經體外合成后,出現1條分子量約在55 000的蛋白帶,這是未被糖基化的蛋白帶。在粗面內質網存在下,L358 (S1?S2 loop)、L393(S2?S3 loop)、E467(S4?S5 loop)和P550(S6?S7 loop)出現一條明顯的糖基化蛋白帶,說明S1?S2連接、S2?S3連接、S4?S5連接和S6?S7連接位于內質網腔內(細胞外),人工導入的G?loop上的的糖基化受體位點被糖基化;其糖基化效率分別為30%、66%、20%和39%(圖2C)。在粗面內質網存在下獲得的蛋白經過蛋白酶K處理后,位于內質網腔外(細胞內)的肽鏈被蛋白酶K消化降解,蛋白帶消失(圖2B)。
2.3使用截短蛋白進行TRPC5各片段的膜整合分析
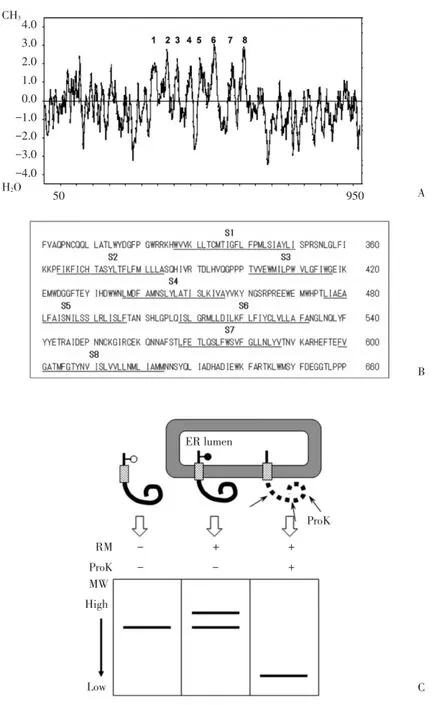
圖1 TRPC5跨膜片段預測和膜拓撲結構分析方法圖示Fig.1 Prediction of membrane spanning segments in TRPC5 and cartoons for topological analysis
本研究采用另一種策略,使用截短蛋白對TRPC5各片段的膜整合又進行了分析。在含有不同疏水片段的構建體的C末端融合一個報告基因PLg,該報告基因含有一個糖基化位點(圖3A)。如果該報告基因在翻譯轉運時被糖基化,說明截短蛋白的C末端位于內質網腔內(細胞外)。SDS?PAGE結果(圖3B)顯示,S1?PLg、S1?S2?PLg、S1?S4?PLg和S1?S6?PLg經體外合成后,在粗面內質網存在下分別出現了1條糖基化蛋白帶,且經蛋白酶K處理后,仍然有蛋白帶殘留,說明截短蛋白的C末端,即S1?S2連接、S2?S3連接、S4?S5連接和S6?S7連接,是位于內質網腔內(細胞外)的,其之前的最后一個疏水片段是以Ncyt/Cexo的形式整合到膜上的。每個截短蛋白的糖基化效率(圖3C)S1?PLg為37%,S1?S2?PLg 為51%,S1?S4?PLg為17%,S1?S6?PLg為34%。
2.4TRPC5通道蛋白膜整合分析
根據圖2和圖3的數據結果,推測TRPC5通道蛋白的膜整合可能采取3種基本模式(圖4)。模式Ⅰ:N末端位于細胞內,S1跨膜,而S2位于細胞外(內質網腔內),S3~S8疏水片段按順序跨膜,C末端位于細胞內;模式Ⅱ:N末端和S1位于細胞內,S2~S8疏水片段按順序跨膜,C末端位于細胞內(與VANNIER等[6]1998年報道的TRPC3模型相同);模式Ⅲ:N末端位于細胞內,S1和S2跨膜,而S3位于細胞內,S4~S8疏水片段按順序跨膜,C末端位于細胞內(與DOHKE等[7]2004年報道的TRPC1模型相同)。根據2種實驗方法均得出的S1?S2連接、S2?S3連接、S4?S5連接和S6?S7連接位于內質網腔內(細胞外)的結果,我們推測出TRPC5各疏水片段可能的膜整合方式。S4?S5連接和S6?S7連接位于細胞外,可以推測出TRPC5的S4~S8疏水片段按順序整合到膜上,成為跨膜片段,C末端位于細胞內。而S1?S2連接、S2?S3連接位于細胞外,可以推測出S1~S3疏水片段的膜整合方式存在2種可能性。第1種是TRPC5通道蛋白采用模式Ⅰ的跨膜方式;第2種是TRPC5采用模式Ⅰ與模式Ⅱ的混合跨膜方式。
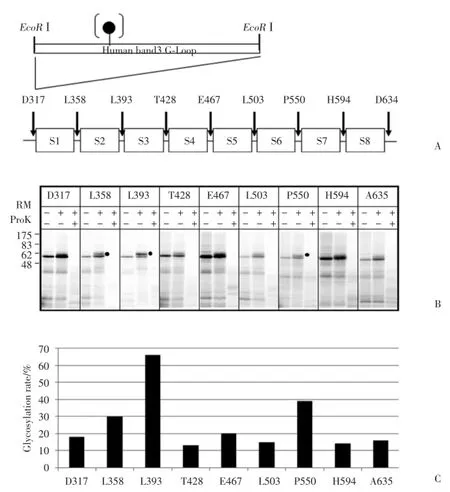
圖2 使用N和C末端切除蛋白進行TRPC5各片段的膜整合分析Fig.2 Membrane integration assays of each transmembrane segment of TRPC5 using N?and C?terminus?deleted proteins
3 討論
通常膜蛋白在內質網合成的同時,腔內會發生糖基化現象,而腔外發生蛋白水解的現象。根據這一特點,科學家們常采用各種無細胞系統進行蛋白的體外轉錄、翻譯和轉運,在粗面內質網存在的情況下,體外模擬膜蛋白合成和轉運過程,觀察目標蛋白的糖基化和蛋白水解的現象,從而分析膜蛋白的拓撲結構[9]。但膜蛋白的糖基化反應有效進行需要一定的條件,那就是膜蛋白的糖基化受體位點應該與膜上寡糖基轉移酶的活性位點結合,而這個位點與內質網膜之間的距離遵守12?14規則。該規則認為,糖基化受體位點的N端距內質網膜表面至少有12個殘基的距離,其C端至少有14個殘基的距離,該受體位點才能被有效糖基化[10]。此規則已被用來分析膜蛋白的拓撲結構[11?12]。
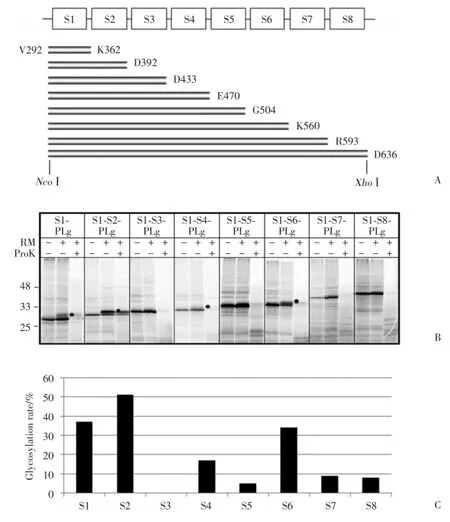
圖3 使用截短蛋白進行TRPC5各片段的膜整合分析Fig.3 Membrane integration assays of each transmembrane segment of TRPC5 using truncated proteins
本研究根據人工導入或自然存在的糖基化位點發生糖基化需要滿足該位點位于內質網膜腔內(即細胞外)的原理,設計2種實驗策略來推測TRPC5的膜拓撲結構。一種是在TRPC5的可能跨膜片段之間的連接上導入含糖基化位點的G?loop,觀察G?loop是否被糖基化,從而推出該連接是否位于細胞外;另一種是在含可能跨膜片段的TRPC5截短蛋白的C末端連接含糖基化位點的PLg報告基因,觀察該報告基因是否被糖基化,推出TRPC5截短蛋白的C末端是否位于細胞外,從而推出C末端所在的連接是否位于細胞外。
2種實驗策略的結果均顯示,TRPC5通道蛋白的S1?S2連接、S2?S3連接、S4?S5連接和S6?S7連接位于細胞外。根據該結果,可推測出S4~S8疏水片段為跨膜片段,與之前VANNIER[6]和DOHKE[7]發表的TRPC3和TRPC1的S4~S8為跨膜片段的結果一致。而S1~S3部分與之前的數據不符。我們分析了TRPC3、TRPC1和本研究的TRPC5實驗方法,發現結果不同可能由于實驗方法的差異。VANNIER的TRPC3實驗用的是全長的通道蛋白,而DOHKE的TRPC1實驗和我們的TRPC5實驗用的均是去除N端的通道蛋白。TRP通道家族的N端含有錨蛋白結構域,有助于通道形成四聚體結構,而這種四聚體結構的形成很可能抑制疏水片段S1的膜插入功能,這樣S2~S8會按通常的模式插入膜中,形成TRPC3模式。而N端去除之后,疏水片段S1沒有限制,但S1應該具有一定的跨膜能力,所以S1會引導一部分通道蛋白插入膜中,形成跨膜片段,最終使得TRPC5的膜拓撲結構可能形成TRPC3(Ⅱ型)模式和TRPC1(Ⅲ型)模式的混合體。我們需做進一步的實驗來驗證。首先,我們需要檢驗各個疏水片段的膜插入能力,然后,設計使用全長的通道蛋白來檢測TRPC5的膜拓撲結構,并同時檢測錨蛋白結構域在TRP通道蛋白跨膜過程中的作用。本研究的糖基化結果顯示,TRPC5的S4~S8疏水片段按順序整合到膜上,成為跨膜片段;而S1~S3疏水片段的膜整合方式存在2種可能性,一種是S2疏水片段位于細胞外,S1、S3為跨膜片段,另一種是同時存在2種方式,一部分TRPC5通道的S1位于細胞內、S2~S3為跨膜片段,而另一部分的TRPC5通道的S1~S2為跨膜片段、S3位于細胞內。我們需要進行進一步研究來明確TRPC5的S1~S3疏水片段的膜整合方式。
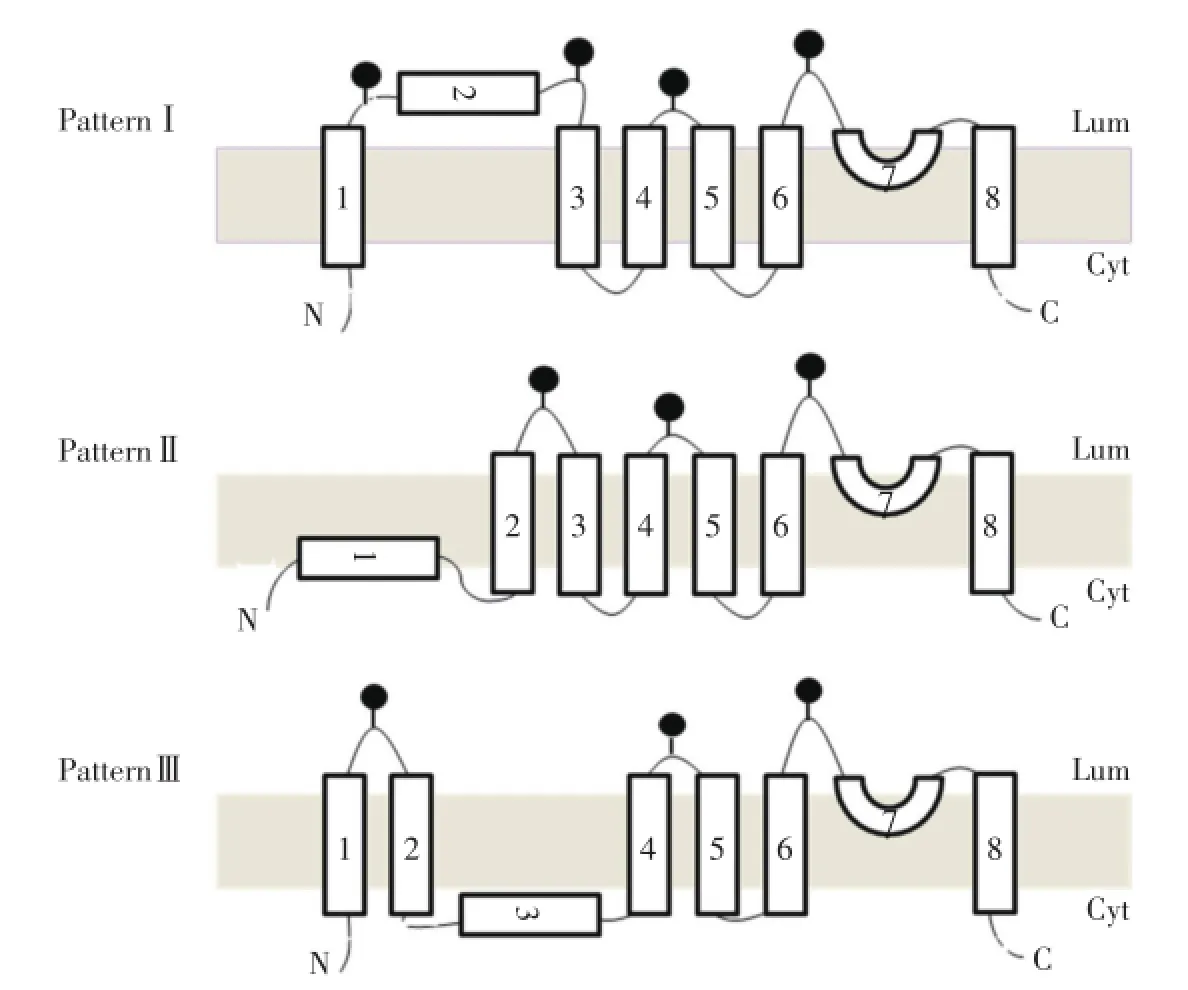
圖4 TRPC5可能的膜拓撲結構模型Fig.4 Possible topological model of TRPC5
參考文獻:
[1]COSENS DJ,MANNING A.Abnormal electroretinogram from a Dro?sophila mutant[J].Nature,1969,224(5216):285-287.
[2]CLAPHAM DE.TRP channels as cellular sensors[J].Natrue,2003,426(6966):517-524.
[3]NILIUS V,VOETS T.TRP channels:a TR(I)P through a world of multifunctional cation channels[J].Pflugers Arch,2005,451(1):1-10.
[4]FUSCO FR,MARTORANA A,GIAMPA C,et al.Cellular localiza? tion of TRPC3 channel in rat brain:preferential distribution to oligo?dendrocytes[J].Neurosci Lett,2004,365(2):137-142.
[5]韓重陽,王曉良.瞬時受體電位通道研究進展[J].生理科學進展,2008,39(1):27-32.
[6]VANNIER B,ZHU X,BROWN D,et al.The membrane topology of human transient receptor potential 3 as inferred from glycosylation?scanning mutagenesis and epitope immunocytochemistry[J].J Biol Chem,1998,273(15):8675-8679.
[7]DOHKE Y,OH YS,AMBUDKAR IS,et al.Biogenesis and topology of the transient receptor potential Ca2+channel TRPC1[J].J Biol Chem,2004,279(13):12242-12248.
[8]VON HEIJNE G.Membrane?protein topology[J].Nat Rev Mol Cell Biol,2006,7(12):909-918.
[9]ZHANG L,SATO Y,HESSA T,et al.Contribution of hydrophobic and electrostatic interactions to the membrane integration of the Shaker K+channel voltage sensor domain[J].Proc Natl Acad Sci U S A,2007,104(20):8263-8268.
[10]NILSSON IM,VON HEIJNE G.Determination of the distance be?tween the oligosaccharyltransferase active site and the endoplas?mic reticulum membrane[J].J Biol Chem,1993,268(8):5798-5801.
[11]張麗艷,尚德志,汲坤,等.糖基化掃描方法分析KvAP通道電壓感受器S3?S4在膜中所處位置[J].中國醫科大學學報,2013,42 (1):10-13.
[12]尚德志,張麗艷,汲坤,等.Shaker通道電壓感受器S3?S4片段末端氨基酸分析[J].中國醫科大學學報,2012,41(12):1077-1079.
(編輯陳姜)
網絡出版時間:
中圖分類號R338.8
文獻標志碼A
文章編號0258-4646(2016)07-0610-06
DOI:10.12007/j.issn.0258?4646.2016.07.008
基金項目:遼寧省科技廳科學事業公益研究基金(2014001024);沈陽醫學院科學研究基金(20141005)
作者簡介:蘇秋香(1962-),女,副教授,本科.
通信作者:張麗艷,E-mail:1580471013@qq.com
收稿日期:2016-04-06
Transmembrane Segment Analysis of Transient Receptor Potential Channel
SU Qiuxiang1,ZHANG Liyan2,JI Kun2,ZHANG Gaigai2,ZHANG Lihua2
(1.Department of Medical Functional Experimentation,College of Basic Medical Sciences,Shenyang Medical College,Shenyang 110034,China;2.Department of Pathophysiology,College of Basic Medical Sciences,Shenyang Medical College,Shenyang 110034)
AbstractObjectiveTo investigate the membrane topology of transient receptor potential(TRP)channel.MethodsGlycosylation method was used to investigate the membrane integrations of each hydrophobic segment of canonical TRP(TRPC5).ResultsIn TRPC5 channel,S4?S8 segments were integrated into membrane with Ncyt/Cexo and Nexo/Ccyt orientations sequentially,and C?terminus was intracellular.S1?S3 segments were integrated into membrane with two possible types.One was that S1 and S3 were integrated into membrane,whereas S2 was left out of the membrane on the cytosolic side;and the other was a mixed type that S1 and S3 were exposed to cytoplasm respectively.Both of them,the N?termi?nus was intracellular.ConclusionS4?S8 segments of TRPC5 are transmembrane segments.The integrations of S1?S3 segments into membrane need to be further investigated.
Keywordstransient receptor potential channel;membrane topology;transmembrane segment analysis
