加入放歸個體后的栗子坪保護區大熊貓小種群遺傳結構研究
2016-09-02 09:06:18陳子君黃蜂何可馮菲菲古曉東楊旭煜戴強齊敦武楊志松
河池學院學報 2016年2期
陳子君, 黃蜂,2, 何可, 馮菲菲,古曉東,楊旭煜,戴強, 齊敦武,楊志松
(1.西華師范大學 西南野生動植物資源保護教育部重點實驗室, 四川 南充 637002;2.四川栗子坪國家級自然保護區管理處, 四川 石棉 625400;3.四川省野生動物資源調查保護管理站, 四川 成都 610081;4.中國科學院成都生物研究所, 四川 成都 610041;5.成都大熊貓繁育研究基地, 四川 成都 610086)
?
加入放歸個體后的栗子坪保護區大熊貓小種群遺傳結構研究
陳子君1, 黃蜂1,2, 何可1, 馮菲菲1,古曉東3,楊旭煜3,戴強4, 齊敦武5,楊志松1
(1.西華師范大學 西南野生動植物資源保護教育部重點實驗室, 四川南充637002;2.四川栗子坪國家級自然保護區管理處, 四川石棉625400;3.四川省野生動物資源調查保護管理站, 四川成都610081;4.中國科學院成都生物研究所, 四川成都610041;5.成都大熊貓繁育研究基地, 四川成都610086)
小相嶺山系是現有大熊貓種群數量最少的山系之一。國家林業局擬通過在該山系的栗子坪國家級自然保護區放歸大熊貓“瀘欣”、“淘淘”、“張想”來復壯大熊貓小種群。為評估加入3只放歸個體后的小種群遺傳結構,2012~2013年在研究區域采集大熊貓糞便樣品92份,鑒定出14只個體(含放歸個體)。放歸個體加入后,測得放歸個體與本地大熊貓小種群親緣關系較遠。種群年齡組成為青年組8只、成年組4只、未知年齡個體2只。平均觀測雜合度(Ho)為0.557,平均期望雜合度(He)為0.581,遺傳多樣性處于中等偏上水平。5個位點上共計增加6個等位基因,平均等位基因數(MNA)達到3.667。遺傳基因簇由1簇增加為4簇,出現新的基因頻率。以上結果顯示放歸個體加入后該區大熊貓小種群遺傳多樣性有所提高,放歸成效初顯。
大熊貓;小種群;遺傳多樣性;放歸;栗子坪國家級自然保護區
放歸是促進小種群復壯,恢復破碎化棲息地的有效方法[1-2]。國內外已有不少成功案例,國內的如普氏野馬(Equus ferus przewalskii)、麋鹿(Elaphurus davidianus )、揚子鱷(Alligator sinensis)等。國外的如加拿大盤羊(Ovis canadensis)、美洲野牛(Bison bison)、美洲黑熊(Ursus americanus)等。然而,也有少數放歸活動因近交[3]、遺傳多樣性丟失[4]或者人類干擾、天敵捕食[5]而成為失敗的嘗試。因此,及時分析放歸成效,總結經驗、教訓,對放歸工作的進一步開展有著極為重要的意義。
小相嶺山系是現有大熊貓種群數量最少的山系之一,種群生存力分析表明,如果與其他種群沒有遺傳交流或者無外來個體補充,該小種群極有可能在80年內會趨于滅絕[6]。為提高該區大熊貓小種群生存能力,促進大熊貓小種群的復壯,(截止到本研究開展的2013年)國家林業局在小相嶺山系的栗子坪國家級保護區先后放歸了3只大熊貓(Ailuropoda melanoleuca):“瀘欣”已放歸超過6年,“淘淘”已放歸超過3年,“張想”也已放歸超過2年。然而,加入放歸個體后,栗子坪自然保護區大熊貓小種群遺傳結構是否有所改善,目前仍不清楚。本研究利用野外調查收集的大熊貓糞便,通過糞便咬節測量、糞便DNA性別鑒定以及微衛星分析,確定了放歸地大熊貓種群的年齡結構、性比和種群遺傳結構,并從種群遺傳上對放歸成效進行了評估。這一評估結果有助于確定將來大熊貓放歸和小種群復壯項目的方向和重點,對其他瀕危珍稀野生動物的小種群復壯工作也有很好的參考價值。
1 研究對象及區域

圖1 研究區域區位示意圖
栗子坪自然保護區位于四川石棉縣境內,地理位置為E 102°10′33″~102°29′07″,N 28°51′02″~29°08′42″;南北長23 km,東西寬17.8 km,總面積47 940 hm2。區內最高海拔為4 551 m,最低海拔1 330 m,最大相對高差達3 221 m,平均相對高差大于2 000 m。保護區是小相嶺山系大熊貓種群的核心分布區和關鍵棲息地,也是小相嶺山系大熊貓種群遺傳基因交流的關鍵走廊帶和重要集中分布區。
本項目研究區域位于栗子坪自然保護區內公益海保護站周邊,面積約為23 251.45 hm2。“瀘欣”、“淘淘”、“張想”先后放歸于該區(詳見圖1和表1),遺傳分析所用糞便也采集自這一區域。這一區域的大熊貓棲息地與周邊大熊貓棲息地存在較為嚴重的隔離[7-8],第四次全國大熊貓調查也認為該區域大熊貓種群是一個相對獨立的局域種群[9]。

表1 放歸大熊貓個體信息
2 研究方法
2.1野外采樣
野外樣品于2012年4月至2013年12月采集自保護區內公益海保護站周邊(見圖1)。 研究區域內調查樣線的抽樣強度為1 條/200 hm2,采用非損傷性遺傳取樣方法在野外收集糞便樣品[10]。 在采樣同時,對所有的樣品采集點均進行GPS定位并記錄。
2.2大熊貓年齡組劃分
測量采集到的大熊貓糞便的咬節長度,采用“離差平方和法”對咬節長度進行聚類,參照胡錦矗的年齡組劃分標準[11]確定個體年齡段:幼年組(小于1.5 歲),咬節值小于30 mm,糞便直徑30 mm以下;青年組(大于等于1.5歲,并且小于5.5 歲),咬節值在30~35 mm(包含30 mm) 之間, 糞便直徑30~55 mm(包含30 mm);成年組(大于等于5.5歲,并且小于20 歲),咬節長度在35~40 mm(包含35 mm)之間,糞便直徑55~65 mm (包含55 mm);老年組(大于等于20 歲),咬節長度在40 mm(包含40 mm)以上,糞便直徑在65 mm(包含65 mm)以上。
2.3糞便微衛星DNA提取與分析
使用QIAGEN公司的QIAamp DNA Stool kit試劑盒進行大熊貓糞便DNA的提取,性別鑒定PCR、微衛星PCR以及結果判定參考胡義波[12]、詹祥江[10]的方法,我們挑選了12對微衛星引物(Ame -μ10, Ame -μ11, Ame -μ22, Ame -μ13, Ame -μ15, Ame -μ24, Ame -μ26, Ame -μ27以及AY161179, AY161195, AY161213, AY161217)來對大熊貓糞便DNA進行微衛星擴增[12]。
因大熊貓糞便中DNA含量較低,為避免因引物選取、PCR擴增、數據讀取等引起的誤差,我們利用MICRO-CHECKER軟件[13]檢測基因分型數據中是否存在無效等位基因(Null alleles)、大片段等位基因丟失(Large allele dropout)及鬼影峰誤判(Stutter scoring error)等分型錯誤。利用GIMLET 1.3.3軟件[14]計算了12個座位的聯合P(ID)值,以確定選擇的微衛星引物數量是否能將影子效應(Shadow effect)[15]減少到一個不顯著的水平。
2.4個體鑒定與種群數量估計
利用MStools插件[16],尋找基因分型數據中相匹配的基因型。判斷為同一個個體的標準是:所有座位上的基因型都相同,或者只有一個座位上的一個等位基因存在差別[17]。由于我們采樣分析是針對放歸成效的研究,糞便采集也集中在放歸大熊貓主要活動的區域(依據放歸大熊貓佩戴的GPS項圈確定),可以認為出現在此區域外或是很少出現在該區域的個體被捕捉概率很低,因此我們選擇CAPWIRE軟件里TIRM(Two Innate Rates Model)模型[10]進行種群數量估計。
2.5種群遺傳多樣性及親緣關系分析
利用ARLEQUIN 3.5[18]和GENEPOP 3.4[19]計算期望雜合度(He, expected heterozygosity)、表觀雜合度(Ho, observed heterozygosity)、等位基因數(A),測算總體和各個位點是否符合哈溫平衡(Hardy-Weinberg)。利用FSTAT 2.9.3.2[20]來檢驗位點是否存在連鎖不平衡(Linkage disequilibrium)。利用STRUCTURE方法[21]檢測研究區域所有個體中存在的遺傳基因簇。利用Kingroup v2.08[22]來測算個體間的親緣關系遠近,其中個體間的親緣關系值(pairwise relatedness, r)參考Wang[23]的研究。
3 結果
3.1種群數量、性比及年齡結構
本研究從92 份樣品(含糞便樣品及放歸個體血樣)中得到52個可靠基因型。TIRM模型分析表明,研究區域大熊貓種群數量(含3 只放歸個體)為25 只(95%置信范圍:14~28)。種群數量最低估計值(lowCI)為14 只(與實驗室分析鑒定結果一致),因此研究區域中大熊貓種群最保守的數量應該是14 只。14只個體分布在公益海保護站區域,海拔集中在2 291~3 510 m(見圖2),分別是8 只雌性、6只雄性,雌雄比例為4∶3。其中“瀘欣”、“淘淘”和“張想”為放歸個體,年齡有準確記錄,無需通過糞便咬節確定年齡,另有兩只個體未能獲得有效咬節數據,無法判斷年齡,其余個體的年齡段均根據咬節法測得。結果表明,本地種群的年齡組成為:青年組6 只,成年組3 只,未知年齡個體2只(見表2)。從年齡結構上來看,青年組個體數明顯多于成年組個體數,年齡組成為增長型。 加入放歸個體后的大熊貓種群年齡組成為:青年組8 只、成年組4只,未知年齡個體2只(見表2),與放歸前比較,年齡組結構的增長型特征更為明顯。

表2 本研究大熊貓種群年齡組組成
3.2本地小種群遺傳結構
MICRO-CHECKER檢驗表明,本研究基因分型數據結果未受無效等位基因、大片段等位基因丟失及鬼影帶誤判等分型錯誤的顯著干擾。GIMLET分析顯示,12 個微衛星座位的聯合區分率很高,兩只個體具有相同基因型的無偏概率P(ID)為3.286×10-9,即使為雙胞胎,判斷錯誤的概率P(sib) 也僅為0.04778%,小于0.01[15]。哈溫平衡檢驗結果顯示研究區域本地大熊貓小種群未偏離哈溫平衡(P>0.05)。連鎖不平衡分析結果顯示本研究采用的12 個位點,對當地大熊貓小種群而言不存在連鎖遺傳。12個微衛星位點分析結果顯示研究區域本地大熊貓小種群的平均觀測雜合度(Ho)為0.635,平均期望雜合度(He)為0.608。平均等位基因數(MNA)為3.455(見表3)。STRUCTURE 分析結果顯示K=1時(圖3A),LnP(D)具有最大的平均值,圖3B表明研究區域本地大熊貓小種群整體上是一個遺傳基因簇。
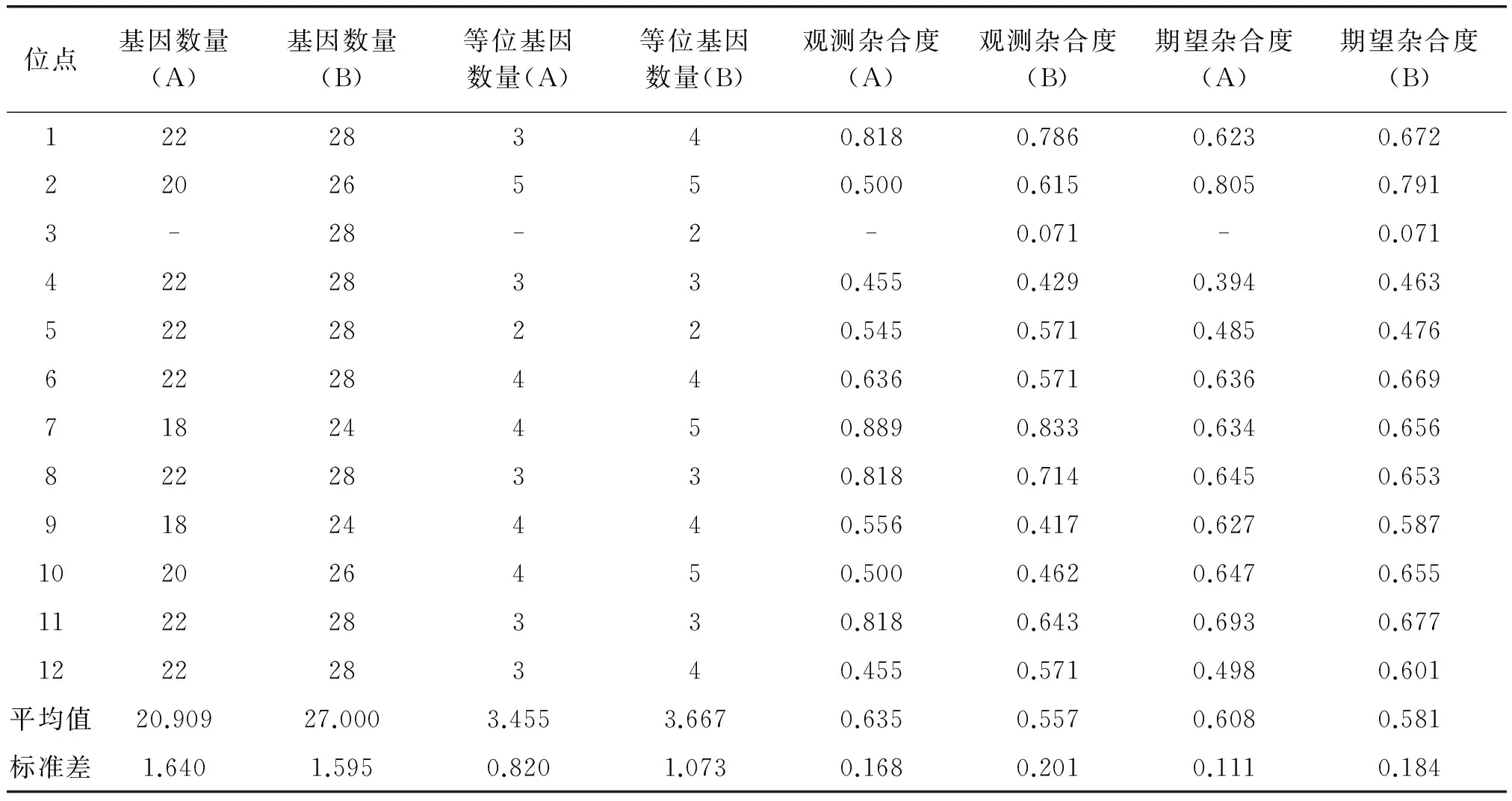
表3 研究區域本地大熊貓小種群(代號A)與加入放歸個體后的種群(代號B)遺傳多樣性信息

圖3 LnP(D)值及遺傳基因簇圖圖A:本地小種群LnP(D)值;圖C:放歸個體加入后的種群LnP(D)值;圖B圖D分別為K=1、K=4的遺傳基因簇圖
3.3加入放歸個體后小種群遺傳結構
3.3.1放歸個體和本地大熊貓親緣關系
親緣關系分析結果顯示(見表4),“瀘欣”、“淘淘”、“張想”和整個本地小種群的親緣關系都較遠,與其它13 只個體的親緣系數(r)平均值平均為-0.07、-0.21、-0.23。
雖然“瀘欣”和lzp6、lzp7、lzp10、lzp11、“張想”的親緣系數(r)>0,但是這五只個體均為雌性,不存在因親緣關系近而帶來的近交風險。“淘淘”和lzp2、lzp7、“張想”的親緣系數(r)分別為0.05、0.10、0.10,但是r<0.125,“淘淘”和這三只個體的親緣關系水平低于雙重表親(Double first cousins),不存在明顯的因親緣關系較近而帶來的近交風險。“張想”和lzp6、“淘淘”、“瀘欣”的親緣系數(r)分別為0.03、0.10、0.11,r<0.125,“張想”和這三只個體的親緣關系水平也低于雙重表親(Double first cousins),不存在明顯的因親緣關系近而帶來的近交風險。因lzp7是雌性大熊貓,雖然它和“張想”親緣系數(r)為0.30,屬于親子/全同胞(parent-offspring/full sibs)的水平,但也不存在因親緣關系近的近交風險。

表4 研究區域大熊貓個體親緣關系
3.3.2加入放歸個體后的遺傳結構
根據哈溫平衡檢驗分析結果,加入放歸個體后的大熊貓小種群仍未偏離哈溫平衡(p>0.05)。連鎖不平衡分析結果顯示對于加入放歸個體后的大熊貓小種群,也不存在連鎖遺傳。12個微衛星位點分析結果顯示:加入放歸個體后種群平均觀測雜合度(Ho)為0.557,平均期望雜合度(He)為0.581(見表3)。相比Zhang等[24]的研究,加入放歸個體后的遺傳多樣性仍處于中等偏上的水平。平均等位基因數(MNA)增加到3.667,表現在Ame-μ11(Locus1)、Ame-μ22(Locus3)、Ame-μ24(Locus7)、Ame-μ13(Locus10)和Ame-μ27(Locus12)5個位點上,共計增加6 個等位基因。 STRUCTURE分析結果表明,K=4時(見圖3C),LnP(D)具有最大的平均值,遺傳基因簇由一簇變為了四簇(見圖3D),這表明放歸個體加入后,出現了新的基因頻率。
4 討 論
三只放歸個體加入當地種群后,研究區域大熊貓小種群的等位基因數有所增加,同時也出現了新的基因頻率,這表明該區大熊貓小種群的遺傳多樣性確實呈增長趨勢。遺傳多樣性的高低在一定程度上反映了大熊貓種群的進化潛力[25],因此放歸個體的加入有助于研究區域大熊貓小種群的種群遺傳多樣性的提高,將對該種群的復壯帶來積極影響。
本研究分析結果顯示放歸個體與研究區域本地大熊貓小種群親緣關系較遠。楊波等[26]的研究表明:親緣關系越遠,釋放個體與本地種群組成的混合種群內近交率會越低,從而減少近交衰退的風險,更有利于改善野生種群遺傳水平。因此這三只放歸個體的加入,能有效提高研究區域本地大熊貓小種群的遺傳多樣性水平。
迄今,“瀘欣”、“淘淘”、“張想”已放歸分別超過6年、3年、2年,它們在大熊貓放歸地——栗子坪國家級自然保護區生存狀況良好。年齡組結構分析顯示,加入放歸個體后,研究區域大熊貓小種群年齡結構得到進一步改善。此外,該區域的紅外相機監測結果顯示,“瀘欣”已成功帶崽,表明放歸個體已經初步融入當地種群,放歸工作取得初步成效。若其余兩只放歸個體也能成功繁殖,將進一步促進該地區大熊貓小種群的復壯。
目前,小相嶺山系大熊貓種群數量仍然很少,成功放歸三只大熊貓遠不足以復壯當地小種群。針對這一問題,該山系還需進一步持續開展大熊貓放歸工作。而在放歸個體的選擇上,需要比較放歸個體和當地小種群的遺傳背景,優先選擇能有效提高當地小種群遺傳多樣性的個體。
[1]ZHU L F, ZHAN X J, MENG T, et al. Landscape features influence gene flow as measured by cost-distance and genetic analyses: a case study for giant pandas in the Daxiangling and Xiaoxiangling mountains [J]. BMC genetics, 2010(1):72.
[2]ZHU L F, ZHANG S N, GU X D, et al. Significant genetic boundaries and spatial dynamics of giant pandas occupying fragmented habitat across southwest China [J]. Molecular Ecology, 2011, 20(6):1122-1132.
[3]BIEBACH I, KELLER L F. Inbreeding in reintroduced populations: the effects of early reintroduction history and contemporary processes [J]. Conservation Genetics, 2010(2):527-538.
[4]ALCAIDE M., NEGRO J J., SERRANO D, et al. Captive breeding and reintroduction of the lesser kestrel Falco naumanni: a genetic analysis using microsatellites [J]. Conservation Genetics, 2010(11): 331-338.
[5]GREY-ROSS R, DOWNS C T, KIRKMAN K.. Reintroduction Failure of Captive-Bred Oribi (Ourebia ourebi) [J]. South African Journal of Wildlife Research, 2009, 39(1): 34-38.
[6]朱磊, 吳攀文, 張洪, 等. 小相嶺山系大熊貓種群生存力分析 [J]. 西華師范大學學報( 自然科學版), 2008, 29(2): 112-116.
[7]ZHU L F, ZHAN X J, WU H, et al. Conservation implications of drastic reductions in the smallest and most isolated populations of giant pandas [J]. Conservation Biology, 2010, 24(5): 1299-1306.
[8]青菁, 胥池, 楊彪, 等. 小相嶺山系大熊貓廊道規劃[J]. 生態學報, 2016, 36(4): 1-9.
[9]四川省林業廳. 四川的大熊貓[M]. 成都:四川科學技術出版社,2015.
[10]詹祥江.利用非損傷性遺傳取樣研究大熊貓的種群數量和擴散模式 [D]. 中國科學院動物研究院, 2006.
[11]胡錦矗. 從野外大熊貓的糞便估計年齡及其種群年齡結構的研究 [J]. 獸類學報, 1987, 7 (2): 81-84.
[12]胡義波. 涼山山系大熊貓的種群歷史、景觀與空間遺傳格局研究 [D]. 北京:中國科學院研究生院, 2008.
[13]VAN OOSTERHOUT C, HUTCHINSON W F, WILLS D P M, et al. Micro-checker: software for identifying and correcting genotyping errors in microsatellite data [J]. Molecular Ecology Notes, 2004(3): 535-538.
[14]VALIERE N. GIMLET: a computer program for analysing genetic individual identification data [J]. Molecular Ecology Notes, 2002, 2: 377-379.
[15]MILLS L S, CITTA J J, LAIR K P, et al. Estimating animal abundance using noninvasive DNA sampling: Promise and pitfalls [J]. Ecological Applications, 2000, 10(1):283-294.
[16]PARK S D E. Trypanotolerance in west African cattle and the population genetic effects of selection [D]. Dublin:University of Dublin, 2001.
[17]BELLEMAIN E, SWENSON J E, TALLMON D, et al. Estimating population size of elusive animals with DNA from hunter-collected feces: Four methods for brown bears [J]. Conservation Biology, 2005, 19(1): 150-161.
[18]EXCOFFIER L, LAVAL G, SCHNEIDER S. Arlequin (version 3.0): an integrated software package for population genetics data analysis [J]. Evolutionary Bioinformatics Online, 2005, 1: 47-50.
[19]RAYMOND M, ROUSSET F. Genepop (Version1.2): Population Genetics Software for Exact Tests and Ecumenicism [J]. Journal of Heredity, 1995,86(3):248-249.
[20]GOUDET J. FSTAT, a program to estimate and test gene diversities and fixationindices (version 2.9.3). 2001. Updated from Goudet J. FSTAT (Version 1.2): A computer program to calculate F-statistics [J]. Journal of Heredity, 1995, 86: 485-486.
[21]PRITCHARD J K, STEPHENS M, DONNELLY P. Inference of population structure using multilocus genotype data [J]. Genetics, 2000, 155(2): 945-959.
[22]KONOVALOV D A, MANNING C, HENSHAW M T. Kingrouf: a program for pedigree relationship reconstruction and kin group assignments using genetic markers [J]. Molecular Ecology Notes, 2004(4): 779-782.
[23]WANG J. COANCESTRY: a program for simulating, estimating and analysing relatedness and inbreeding coefficients [J]. Molecular Ecology Resources, 2011, 11(1): 141-145.
[24]ZHANG B W, LI M, ZHANG Z J, et al. Genetic Viability and Population History of the Giant Panda, Putting an end to the "Evolutionary dead end"? [J]. Molecular Biology and Evolution, 2007, 24(18): 1801-1810.
[25]張澤鈞, 張陜寧, 魏輔文, 等. 移地與圈養大熊貓野外放歸的探討 [J]. 獸類學報, 2006, 26 (3): 292-299.
[26]楊波, 楊承忠, 涂飛云, 等. 大熊貓野化放歸中的遺傳學分析 [J]. 四川動物, 2013, 32(1):149-155.
[Abstract]The population of giant panda in Xiaoxiangling is one of the smallest . In order to rejuvenate this small panda population, the State Forestry Administration has tried to release three giant pandas(Ailuropoda melanoleuca): “Luxin”, “Taotao” and “Zhangxiang” in the Liziping National Nature Reserve. To assess the genetic structure of this small population after reintroduction, 92 stools of giant pandas were collected in research area from 2012 to 2013, and 14 individuals(including three reintroduced individuals)were identified. After reintroduced the three individuals, measurement proved that the kinship between the reintroduced individuals and local small population was far. The population age composition was: eight individuals in youth group,four individuals in adult group and two individuals in unknown group.The mean observed heterozygosity (Ho) was 0.557. The mean expected heterozygosity (He) was 0.581. Genetic diversity was in the medium level.Five sites increased six alleles.And the mean allele number (MNA) achieved 3.667. What’s more ,the genetic cluster increased to four meaning that new gene frequency had appeared. All these results suggest that the genetic diversity of the small giantpanda population in this region is improved. The reintroduction is beginning to bear results.
[Key words]giant panda(Ailuropoda melanoleuca); small population; genetic diversity; reintroduction; Liziping National Nature Reserve
[責任編輯劉景平]
The Genetic Structure of Small Giant Panda Population in the Liziping Nature Reserve after Reintroduction
CHEN Zi-jun1, HUANG Feng1,2, HE Ke1, FENG Fei-fei1, GU Xiao-dong3, YANG Xu-yu3,DAI Qiang4, QI Dun-wu5, YANG Zhi-song1
(1.Key Laboratory of Southwest China Wildlife Resource Conservation, China West Normal University,Nanchong, Sichuan 637002, China;2.Sichuan Liziping Nature Reserve Management Office, Shimian, Sichuan 625400, China;3.Sichuan Station of Wildlife Survey and Management, Chengdu, Sichuan 610081, China;4.Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu, Sichuan 610041, China;5.Chengdu Research Base of Giant Panda Breeding, Chengdu, Sichuan 610086, China)
Q953
A
1672-9021(2016)02-0001-07
陳子君(1989-),女,四川綿陽人,西華師范大學生命科學學院碩士研究生,主要研究方向:大熊貓保護生物學;楊志松(1975-),男,四川南充人,博士,西華師范大學生命科學學院副教授,碩士生導師,主要研究方向:大熊貓保護生物學、分子系統地理學、 鳥類生態學。
香港海洋公園保育基金資助項目(GP07_1213)。
2016-03-01
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
今日農業(2021年9期)2021-11-26 07:41:24
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
發明與創新·小學生(2021年3期)2021-03-25 11:48:49
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
