馬尾松毛蟲質型多角體病毒非結構蛋白p44在Bac-to-Bac系統中的表達和亞細胞定位
2016-10-13 13:20:43彭晗王洪秀王金昌關麗梅靳亮萬翠香
生物技術通報 2016年3期
關鍵詞:檢測
彭晗王洪秀王金昌關麗梅靳亮萬翠香
(1. 南昌大學生命科學與食品工程學院,南昌 330077;2. 江西省農業科學院農業應用微生物研究所,南昌 330200;3. 江西省科學院微生物研究所,南昌 330029)
馬尾松毛蟲質型多角體病毒非結構蛋白p44在Bac-to-Bac系統中的表達和亞細胞定位
彭晗1王洪秀2王金昌3關麗梅3靳亮3萬翠香1
(1. 南昌大學生命科學與食品工程學院,南昌 330077;2. 江西省農業科學院農業應用微生物研究所,南昌 330200;3. 江西省科學院微生物研究所,南昌 330029)
為探尋馬尾松毛蟲質型多角體病毒(DpCPV 1)p44蛋白的功能,構建了DpCPV 1基因組S8片段的原核表達體系,表達純化蛋白后免疫家兔制備了多克隆抗體。利用Bac-to-Bac桿狀病毒表達系統,構建了3種重組的桿狀病毒質粒(Bacmid-p44、Bacmid-p44-eGFP和Bacmid-eGFP)。轉染昆蟲細胞Sf9進行表達,通過Western blot檢測和蛋白的亞細胞定位觀察。Western blot檢測結果顯示,Bacmid-S8在昆蟲細胞Sf9中表達實際蛋白的大小為35 kD,比在原核系統中表達的蛋白(44 kD)略小;利用激光共聚焦顯微鏡觀察p44-eGFP的融合蛋白的亞細胞定位發現,融合p44的綠色熒光蛋白(eGFP)主要聚集在細胞質中,而未融合的eGFP則分布于整個細胞,說明 DpCPV 1的p44蛋白定位于細胞質中。
馬尾松毛蟲質型多角體病毒;多克隆抗體;真核表達;細胞定位
昆蟲質型多角體病毒(cypovirus,CPV)屬于呼腸孤病毒科,質型多角體病毒屬。根據病毒基因組dsRNA片段在聚丙烯酰胺或瓊脂糖凝膠中電泳圖譜的差異,目前CPV已被分為20個電泳型[1,2]。馬尾松毛蟲質型多角體病毒(dendrolimus punctatus cytoplasmic polyhedrosis Virus 1,DpCPV 1)屬于1型,該病毒基因組S8片段編碼一個由390個氨基酸組成、分子量為43.78 kD的非結構蛋白(p44),是我國的特有種類,于1973年首次分離得到[3]。
隨著結構生物學與信息處理等新技術的運用與發展,包括X射線晶體衍射、低溫電鏡與三維重構技術,質型多角體病毒結構學研究方面已取得突破性進展[4]。Yu等[5]利用低溫電子顯微鏡技術在分辨率上研究了衣殼蛋白的三維結構,在與基因組直接作用的區域中觀察到螺旋與β發卡結構之間的構象改變,同時發現了特有的加帽結構和釋放通道。Cheng等[6]通過低溫電子顯微鏡研究發現,CPV的五聚體塔狀蛋白的酶區域是拓撲結構高度保守并且有5個連接著鳥苷酰基轉移酶和甲基轉移酶區域的獨特的通道;通過氨基酸序列推理,LPP是由S7片段編碼P50蛋白修飾后得到的。Yang等[7]研究發現:當CPV病毒粒子進行轉錄時,衣殼蛋白VP1A、VP1B和塔狀蛋白VP3構象發生變化,衣殼空間擴大和塔狀蛋白周圍通道的加寬,使得基因組RNA從緊湊的病毒粒子衣殼更加靈活的轉錄和輸出。
相對CPV結構蛋白的深入研究,CPV非結構蛋白的研究相對緩慢。文力[8]、張萬菊[9]等對DpCPV 1基因組中的第9片段(S9)的編碼序列進行了cDNA克隆和序列測定,并對NS5蛋白的表達和功能進行了初步分析。汪洋等[10]研究發現由DpCPV基因組第9片段編碼的非結構蛋白NS5蛋白在感染昆蟲細胞時,定位在昆蟲細胞的細胞膜上。段兵[11]和胡建芳等[12]對馬尾松毛蟲CPV基因組第8片段進行序列分析和原核表達。凝膠遷移阻抑分析(EMSA)顯示,由CPV基因組第8片段編碼的p44蛋白具有序列非特異性的ssRNA結合活性,不與dsRNA、ssDNA、dsDNA結合;進一步研究發現p44氨基酸序列116-197 aa之間的區域(富含谷氨酸區域)為單一的RNA結合區域[13]。
目前,質型多角體病毒S8片段編碼的非結構蛋白p44蛋白的真核表達和昆蟲細胞上定位研究還未見報道。本研究利用Bac-to-Bac桿狀病毒表達系統,構建重組Bacmid-S8-eGFP和Bacmid-S8,將DpCPV 1 S8片段和綠色熒光蛋白基因(eGFP)以C端融合的方式在昆蟲Sf9細胞中進行融合表達及細胞定位,旨在為下一步非結構蛋白p44在CPV復制過程中的功能研究提供基礎數據。
1 材料與方法
1.1 材料
1.1.1 細胞系、菌種及質粒 草地貪夜蛾(Spodoptera frugiperda)細胞系Sf9由中國科學院武漢病毒所昆蟲病毒基因工程學科組提供,于27℃培養,生長培養基為Grace's昆蟲細胞培養基(10%胎牛血清)。大腸桿菌(Escherichia coli)DH5α、BL21和DH10B菌株均為本實驗室保存;載體pMD18-T vector系統、T4 DNA Ligase及其buffer購自寶生物工程(大連)有限公司,載體pET-28a、pFastBacDual、pFastBacDual-eGFP(GFP片段插入在PstⅠ和HindⅢ酶切為點之間)和DpCPV 1基因序列S8片段為本實驗室保存。
1.1.2 主要試劑 DM2000 DNA Marker 購自康為世紀生物科技有限公司;DNA MarkerⅢ購自東盛生物科技有限公司產;預染蛋白Marker購自南京生興生物技術公司;異丙基-β-D硫代半乳糖苷(IPTG)購自武漢貝特生物公司;Ni-NTA His.BindTMResins購自Novagen公司;其余藥品均為國產分析純以上。轉染試劑lipofectin 購自Invitrogen公司(美國);熒光染料 Hoechst 33258 購自 Biosharp公司(美國);PVDF膜購自Millipore公司(美國);PCR純化試劑盒購自Promega公司(上海);質粒提取試劑盒購自Promega公司(上海)。
1.1.3 引物合成 根據DpCPV 1基因組S8片段的核苷酸序列,設計引物;除通用引物M13F 和M13R(Cat. No. N530-02,Invitrogen公司,美國)外,其余引物均由上海生工生物工程技術服務有限公司合成。引物列表,見表1。
1.2 方法
1.2.1 以實驗室保存的DpCPV病毒基因組S8為模板,以pTS8F、pTS8R為引物,利用兩步法進行RT-PCR擴增[14]。PCR擴增的片段通過相應的酶切位點酶切,連接到原核表達載體pET-28a(pET28a-S8)。將陽性重組質粒轉化大腸桿菌感受態細胞BL21,1 mmol/L的IPTG于37℃誘導表達4 h。收集菌體,經超聲破碎后離心,獲得包涵體蛋白。用8 mol/L的尿素溶液溶解包涵體蛋白,然后通過Ni-NTA樹脂柱進行蛋白純化。將收集的洗脫流出液裝入透析袋,對含適量甘油的0.01 mol/L PBS pH8.0透析48 h,每4-8 h更換一次PBS溶液。然后,將上述透析袋包埋在聚乙二醇粉末中,對蛋白溶液進行濃縮。并為下一步制備抗體做準備。

表1 PCR擴增產物所需引物列表
1.2.2 抗體制備 純化的目的蛋白常規方法(淋巴加皮下注射)免疫家兔,制備抗血清。實驗兔、鼠飼養,抗原注射及最終采血均委托武漢愛博泰克生物科技有限公司進行。
1.2.3 重組Bacmid病毒的構建及PCR檢測 以pFS8-1F、pFS8-1R和pFS8-2F、pFS8-2R為引物,以pET28a-S8載體為模板進行PCR擴增。將上述膠回收的S8片段分別連接到pFastBacDual和pFastBacDual-eGFP載體上。依照質粒提取試劑盒說明書中介紹的方法,分別提取質粒。酶切鑒定陽性克隆子,分別命名為pFastBac-S8和pFastBac-S8-eGFP。
將pFastBac-S8、pFastBac-S8-GFP和pFastBac-Dual空載體(用作對照)分別轉座到含有AcBacmid和helper質粒的DH10B感受態細胞,涂布于LA培養基平板[含50 μg/mL卡那霉素(kanamycin,Kana),7 μg/mL慶大霉素(gentamicin,Gm),10 μg/mL四環素(tetracycline,Tetra),100 μg/mL 5溴-4-氯-3-吲哚-β-D-乳糖苷(5-bromo-4-chloro-3-indolyl- β-D-galactoside,X-gal)和40 μg/mL異丙-β-D-硫代吡喃半乳糖苷(thiogalactopyranoside,IPTG)],于37℃培養24-48 h。檢查平板上的藍白斑,白斑即重組Bacmid的菌落。挑取白色單菌落,接種于5 mL LB培養基,37℃振蕩培養14 h。取出3 mL于10 000×g 離心10 min,抽提重組Bacmid質粒。抽提Bacmid質粒的具體方法參考Bac-to-Bac?Baculovirus Expression Systems手冊的3.5節。
以上述提取的重組Bacmid為模板,以通用引物M13F和M13R以及目的片段上兩端的引物(表1)進行PCR檢測。鑒定正確的重組Bacmid分別命名為Bacmid-p44、Bacmid-p44-eGFP和Bacmid-Dual,于4℃冰箱中保存。
1.2.4 重組病毒轉染昆蟲細胞 Sf9在35 mm的培養皿中接種5×105的Sf9 細胞,27℃培養過夜。在生物安全柜中棄去上層培養基,加入2 mL無血清的培養基室溫放置1 h。取5 μg重組Bacmid質粒以及6 μL脂質體(Invitrogen,美國),分別用無血清培養基稀釋至100 μL,將兩者混合。靜置15-40 min 后,向脂質體和DNA的混合液中加入800 μL培養基,混勻,移入35 mm培養皿中,培養6 h。移去轉染液,添加2 mL含10%血清的培養基,混勻,培養72 h后倒置熒光顯微鏡下觀察是否有熒光信號。離心回收上清,即為P1病毒貯液,4℃避光保存。
按照2×106個細胞/孔的量將Sf9細胞轉入35 mm的培養皿中,貼壁生長至少1 h。每孔加入適量的上述P1病毒貯液,27℃濕盒孵育72 h。500×g離心5 min取上清,即為P2病毒貯液。重復上述方法擴增P3病毒貯液,用于重組Bacmid的高效表達。
1.2.5 Western blot 檢測 取P3病毒貯液感染Sf9細胞,72 h后將細胞吹起,500×g離心5 min,取適量細胞沉淀,加50 μL的SDS-PAGE上樣buffer,進行SDS-PAGE電泳后轉膜,以p44蛋白多克隆抗體為一抗,辣根過氧化酶標記的羊抗兔的血清(武漢博士德生物工程有限公司)為二抗,用Western blot法檢測p44蛋白的表達。
1.2.6 p44的亞細胞定位 用P2病毒貯液感染貼壁生長24 h的Sf9細胞(覆蓋玻底培養皿50%),27℃培養24 h,移除細胞培養基,用PBS(137 mmol/L NaCl,2.7 mmol/L KCl,10 mmol/L Na2HPO4,2 mmol/L KH2PO4,pH7.4)將細胞洗1遍,加入4%多聚甲醛(北京賽馳生物科技有限公司)(過濾除菌)室溫靜置固定15 min,再用PBS(pH7.4)將細胞洗1遍;加入透化液[0.25% TritonX-100(上海索萊寶生物科技有限公司)],室溫靜置10 min;用PBS(pH7.4)洗滌細胞,加熒光染料hoechst 33258染核5-10 min,然后用PBS洗3遍,再加入1 mL PBS。激光共聚焦顯微鏡(TCS SP2,Leica,德國)觀察綠色熒光分布情況。
1.2.7 序列統計分析與系統發育樹構建 將DpCPV 1基因組S8片段,放入NCBI上進行Blast序列比對(http://www.ncbi.nlm.nih.gov/BLAST/)。根據已經公開的CPV的S8片段進行比對,利用Mega4.0分析軟件進行聚類分析,并采用軟件 ClustalW和PHYLIP3.67的鄰近法構建系統發育樹,其核酸和氨基酸序列比對的bootstrapping數值分別為0.01和0.2。
2 結果
2.1 DpCPV 1基因組S8片段和其編碼非結構蛋白p44的系統發育分析
通過系統發育樹分析,DpCPV 1 基因組S8片段與舞毒蛾質型多角體病毒(LdCPV 1)基因組S8片段具有最高的序列同源性(99%),與另外兩株DPCPV(AY211092.1和AF389469.1)序列同源性均為98%;分別與3株家蠶質型多角體病毒(BmCPV 1)Strain H、Strain I和Strain suzhou 對應片段的同源性為84%、83%和81%(圖1)。
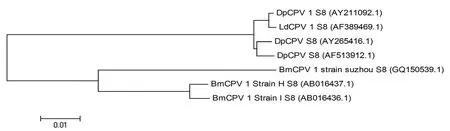
圖1 DpCPV1基因組S8片段的核酸系統發育樹
DpCPV 1 S8片段編碼非結構蛋白p44的系統發育樹分析顯示:p44蛋白與LdCPV 1、BmCPV 1 strain H和冬尺蠖蛾質型多角體病毒(ObCPV 18)的相應非結構蛋白氨基酸序列同源性分別為99%、85%和 35%。另外,我們檢測到了p44蛋白與3株鞘脂菌屬(Sphingobium)細菌的氨基脫氧分枝酸裂解酶(aminodeoxychorismate lyase)有27%的氨基酸序列同源性(圖2)。
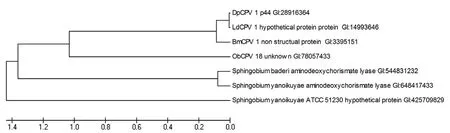
圖2 DpCPV 1非結構蛋白p44的系統發育樹
2.2 DpCPV S8片段的分子克隆和抗體制備
以pTS8F和pTS8R為正反向引物,以DpCPV 1基因組S8片段為模板進行RT-PCR擴增,擴增產物經1.0%瓊脂糖凝膠電泳檢測,擴增片段大小約為1 170 bp(圖3-A)。將S8連接至pET28a載體上,構建表達質粒。用BamHⅠ和Hind III雙酶切驗證正確后(圖3-B),命名為pET28a-S8。經測序,將鑒定正確的克隆質粒pET28a-S8轉化大腸桿菌BL21,加IPTG誘導后離心收集菌體,進行SDSPAGE分析。結果(圖4)顯示,檢測到插入的S8片段的載體有相應蛋白的表達,蛋白大小約為51 kD。因插入的S8片段后面攜帶了一個His純化標簽(大小約7 kD),所以實際插入S8片段表達的蛋白大小為44 kD,與p44蛋白大小相符。
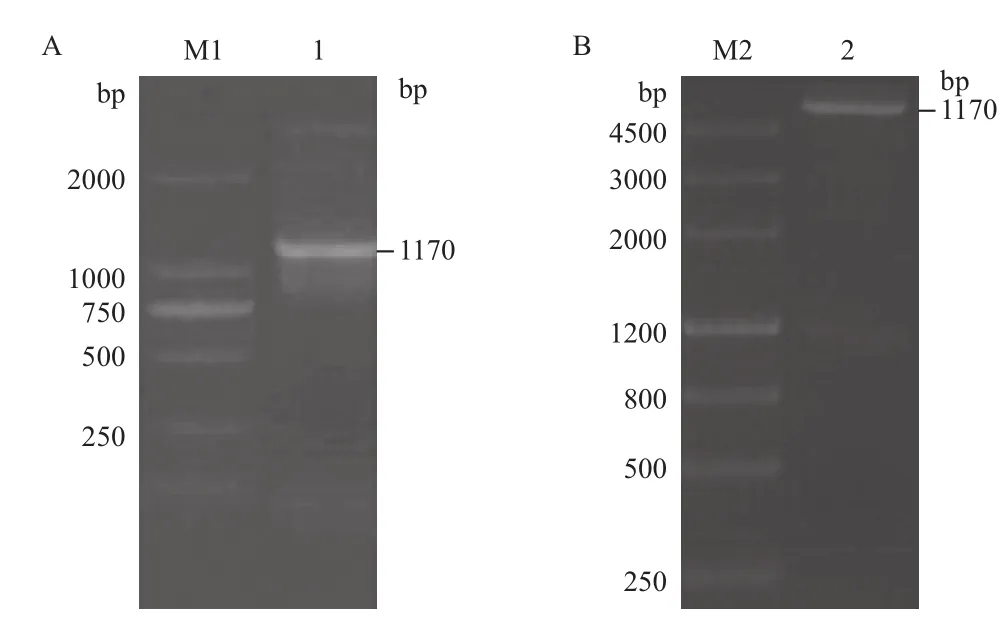
圖3 S8原核表達載體的構建

圖4 SDS-PAGE檢測p44誘導表達結果
通過His純化標簽經Ni-NTA純化樹脂純化包涵體的目的蛋白后,所得純化蛋白免疫家兔,制備p44蛋白的多克隆抗體 anti-p44。
2.3 重組病毒載體構建
將S8片段克隆至pFastBac-eGFP和pFastBac-Dual質粒上,構建載體。用XhoⅠ和KpnⅠ雙酶切驗證后,分別命名為pFast-S8和pFast-S8-eGFP(圖5)。分別將pFast-S8、pFast-S8-eGFP和pFastBacDual(空載對照)轉座到DH10B感受態細胞,經培養后提取并鑒定正確的Bacmid DNA(圖6)。
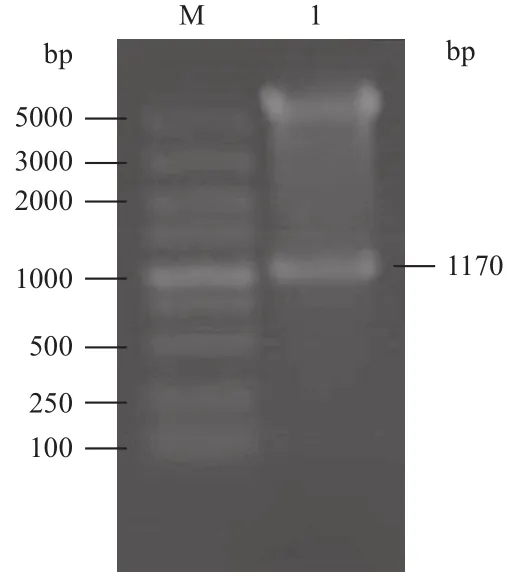
圖5 酶切驗證pFast-S8

圖6 Bac-S8的PCR驗證
2.4 p44的真核表達的Western blot 檢測
用構建的重組Bacmid-S8病毒轉染Sf9 細胞,經SDS-PAGE檢測蛋白的表達情況(圖7-A)。然后以anti-p44為一抗,HRP標記的羊抗兔血清抗體為二抗,經Western blot檢測DpCPV 1 S8編碼蛋白p44在昆蟲細胞Sf9內表達情況。結果(圖7-B)顯示,Bacmid-S8感染的Sf9細胞(第1泳道)在35 kD處有明顯條帶,而空白的Sf9細胞(第3泳道)和經Bacmid-S7感染的Sf9細胞(第2泳道)作為陰性對照,未檢測出條帶。第1泳道檢測到的條帶是p44蛋白的真核表達蛋白,大小為35 kD,但是比原核表達的p44蛋白小了9 kD。
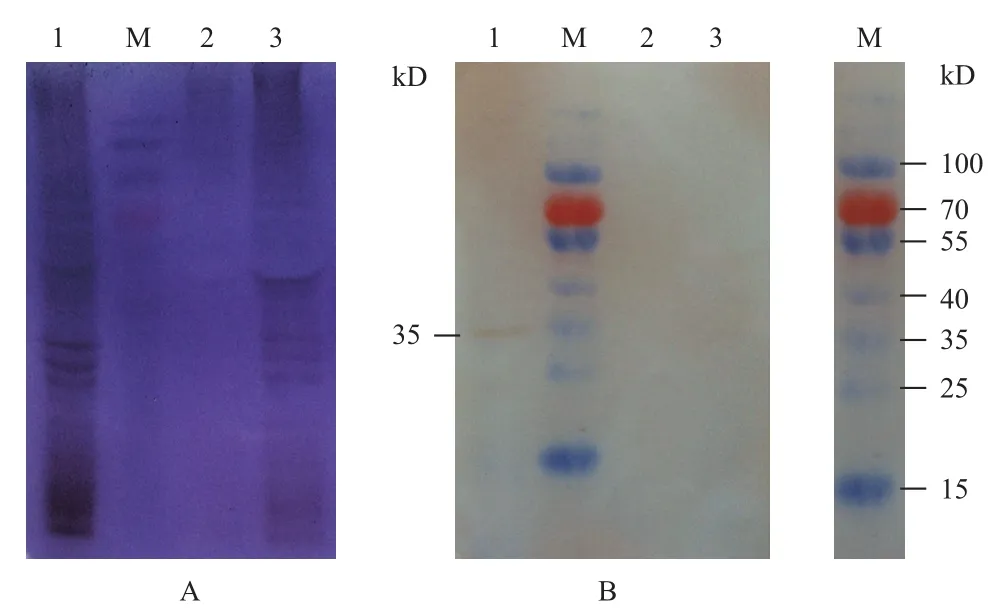
圖7 p44蛋白的真核表達SDS-PAGE(A)及Western blot(B)檢測
2.5 非結構蛋白p44蛋白的亞細胞定位
用P3代病毒感染昆蟲細胞Sf9后24 h,用激光共聚焦顯微鏡觀察蛋白的亞細胞定位情況。結果表明,昆蟲細胞Sf9被桿狀病毒感染后,細胞核明顯增大,經熒光染料 Hoechst 33258染洗后,紫色部位為細胞核。融合了p44的eGFP的蛋白則主要聚集在細胞質中(圖8-A),而未融合p44的EGFP均勻地分布于整個細胞中(圖8-B),說明S8片段編碼的非結構蛋白p44主要定位于細胞質中。
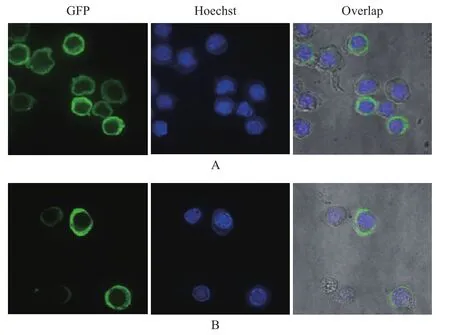
圖8 融合了EGFP的p44在昆蟲細胞中的定位
3 討論
本研究通過克隆 DpCPV 1基因組S8片段到原核表達載體BL21內,原核表達p44蛋白。經SDSPAGE電泳檢測到誘導表達的蛋白大小約51 kD,這是因為在構建表達S8片段的后面添加了一個His純化標簽,純化標簽的蛋白大小約7 kD,因此DpCPV 1基因組S8片段原核表達蛋白的大小約44 kD,與p44的蛋白大小相符。將DpCPV 1 S8片段原核表達蛋白免疫家兔,制備了p44的多克隆抗體。Western blot結果顯示DpCPV 1 S8片段編碼的非結構蛋白p44實際大小為44 kD,與推測的蛋白分子量大小結果一致。
采用Bac-to-Bac桿狀病毒表達系統,將DpCPV 1基因組 S8片段單獨插入以及S8片段與綠色熒光蛋白基因(egfp)以C端融合的方式插入苜蓿銀紋夜蛾核多角體病毒(Autographa californica multiple nucleocapsid NPV,AcMNPV)基因組中,獲得重組的桿狀病毒質粒(Bac-S8和Bac-S8-eGFP),轉染昆蟲細胞Sf9后進行Western blot檢測和蛋白的亞細胞定位的觀察。Western blot檢測結果顯示,Bac-S8在昆蟲細胞Sf9中獲得高效表達,但表達的p44蛋白的大小為35 kD,比原核表達的p44蛋白(44 kD)略小。推測DpCPV 1 S8片段表達的蛋白p44在昆蟲細胞內發生了切割修飾,這種蛋白切割現象此前也在DpCPV 1的其它片段表達的蛋白中發現過。如Jin等[15]將DpCPV 1感染甜菜夜蛾幼蟲,取中腸樣品經Western blot檢測發現,DpCPV 1 S7片段編碼的蛋白p50在感染的第3天表達的蛋白大小為50 kD,在感染的第5天發現該蛋白發生切割修飾,修飾后的蛋白大小為31 kD,上述數據證實了DpCPV S7片段在真核細胞內表達的蛋白發生了切割。因此,我們推測p44蛋白在真核細胞內發生了降解或者被昆蟲細胞的某些酶切割或修飾。
用激光共聚焦顯微鏡觀察Bac-S8-eGFP的融合蛋白的亞細胞定位發現,融合p44的綠色熒光蛋白(eGFP)主要聚集在細胞質中,沒有融合eGFP的分布于整個細胞,說明 DpCPV 1的非結構蛋白p44定位于細胞質中。類似的情況也發生在DpCPV 1 S7片段編碼的結構蛋白p50,Jin等[15]研究發現p50-eGFP同樣定位于細胞的細胞質中。而DpCPV 1的S9片段編碼的非結構蛋白NS5,免疫熒光實驗發現NS5蛋白定位于細胞的細胞質膜上并且跨膜結構域位于蛋白質N端的57-71 aa[16]。相關蛋白的定位研究發現,為研究病毒的生命周期具有重要作用。
4 結論
本研究利用Bac-to-Bac系統首次對DpCPV 1 S8片段進行了真核表達和昆蟲細胞的亞細胞定位,發現S8片段能夠定位到細胞的細胞質。
[1]Shapriro A, Green T, Rao S, et al. Mrophological and molecular characterization of a cypvirus from the mosquito Uranotaenia sapphirina(Diptera:Culicidae)[J]. J Virol, 2005, 79(15):9430-9438.
[2]King AMQ, Adams MJ, Lefkowitz EJ, Carstens EB. Virus taxonomy:classification and nomenclature of viruses. Ninth report of the International Committee on Taxonomy of Viruses[M]. Elsevier Academic Press, 2012:537-536.
[3] 廣東省林業科學研究所. 馬尾松毛蟲多角體病毒研究初報[J].林業科技通訊, 1974, 10:13.
[4]賀倩, 劉小俠, 張青文. 昆蟲質型多角體病毒的研究進展[J].昆蟲知識, 2010, 47(5):834-840.
[5]Yu XK, Jin L, Zhou ZH. 3.88 ? structure of cytoplasmic polyhedrosis virus by cryoelectron microscopy[J]. Nature, 2008, 453:415-419.
[6]Cheng L, Sun JC, Zhang K, et al. Atomic model of a cypovirus built from cryo-EM structure provides insight into the mechanism of mRNA capping[J]. PNAS, 2011, 108:1373-1378.
[7]Yang CW, Gang J, Liu HR, et al. Cryo-EM structure of a transcribing cypovirus[J]. PNAS, 2012, 109(16):6118-6123.
[8]文力, 張珈敏. 馬尾松毛蟲質型多角體病毒NS5蛋白基因的cDNA克隆及序列分析[J]. 中國病毒學, 2003(1):49-53.
[9]張萬菊, 趙淑玲, 張小霞, 等. 馬尾松毛蟲質多角體病毒NS5蛋白的表達和功能初步分析[J]. 中國病毒學(英文版),2006(4):401-404.
[10]汪洋, 張珈敏, 李楊, 等. 馬尾松毛蟲CPV基因組第7片段的cDNA克隆及序列分析[J]. 武漢大學學報:理學版, 2004(2):216-222.
[11]段兵, 趙淑玲, 張海元, 等. 文山松毛蟲質型多角體病毒S8片段cDNA克隆與原核表達[J]. 中國病毒學, 2004(6):627-631.
[12]胡建芳, 張珈敏, 楊娟, 等. 單引物法擴增馬尾松毛蟲CPV基因組第8片段及其序列分析[J]. 中國病毒學, 2003, 18(1):39-43.
[13]Zhao SL, Liang CY, Zhang WJ, et al. Characterization of the RNA-binding domain in the Dendrolimus punctatus cytoplasmic polyhedrosis virus nonstructural protein p44[J]. Virus Research,2005, 114(1):80-88.
[14]Paul RL, Susan JC, Owen EC, et al. Cloning of noncultivatable human rotavirus by single primer amplification[J]. Journal of Virology, 1992, 66(3):1817-1822.
[15]Jin L, Dai CW, Qin TC, et al. Molecular characterization of protein p50 of Dendrolimus punctatus cytoplasmic polyhedrosis virus[J]. Journal of Basic Microbiology, 2013, 53(1):37-44.
[16] Chen WG, Zhang, JM, Dong CJ, et al. Identification of transmembrane domain of a membrane associated protein NS5 of Dendrolimus punctatus cytoplasmic polyhedrosis virus[J]. J Biochem Mol Biol, 2006, 39:412-417.
(責任編輯 馬鑫)
Expression of Dendrolimus puntatus Cytoplasmic Polyhedrosis Virus(DpCPV1)Non-structural Protein p44 in Bac-to-Bac System and Localization in Infected Cells
PENG Han1WANG Hong-xiu2WANG Jin-chang3GUAN Li-mei3JIN Liang3WAN Cui-xiang1
(1. School of Life Sciences and Food Engineering,Nanchang University,Nanchang 330077;2. Institute of Agricultural Applied Microbiology,Jiangxi Agricultural Academy of Sciences,Nanchang 330200;3. Institute of Microbiology,Jiangxi Academy of Sciences,Nanchang 330029)
In order to study the function of the Dendrolimus puntatus cytoplasmic polyhedrosis virus(DpCPV1)protein p44,PCR primers were designed according to the sequence of genome segment S8,the prokaryotic expression vector for genome segment 8 of DpCPV1 was constructed,and the polyclonal antibodies were prepared by immunizing rabbits with the purified expressed protein. Three recombinant plasmids(Bacmid-p44,Bacmid-p44-eGFP and Bacmid-eGFP)were constructed using Bac-to-Bac Baculovirus expression system.,and they were transfected into insect cell Sf9 for the expression. The detection and subcellular localization of the protein were determined by Western blot. The results from Western blot showed that the actual expressed protein of Bacmid-S8 in Sf9 was 35 kD,smaller than the one(44 kD)expressed in the prokaryotic expression vector. The subcellular localization of fusion protein of p44-eGFP was determined by laser scanning confocal microscope,and the fused green fluorescent protein(eGFP)of p44 concentrated in the cytoplasm of the cells,while infused eGFP distributed throughout the whole cell,indicating that p44 protein of DpCPV1 was localized mainly in the cytoplasm of the cell. This work was the first study of having the eukaryotic expression of DpCPV1 S8 and subcellular location of it in insect cells,
Dendrolimus puntatus cytoplasmic polyhedrosis virus;polyclonal antibody;eukaryotic expression;cellular localization
10.13560/j.cnki.biotech.bull.1985.2016.03.020
2015-05-19
國家自然科學基金項目(31260031),江西省科技重大專項基金項目(2014ACF60002),中科院開放基金項目(2014AEM003)
彭晗,女,碩士,研究方向:昆蟲病毒學;E-mail:penghan0830@163.com;王洪秀為本文并列第一作者
靳亮,男,博士,研究方向:昆蟲病毒學;E-mail:Jinliang079@163.com萬翠香,女,博士,副教授,研究方向:應用微生物學;E-mail:cuixiangwan@ncu.edu.cn
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
