妊娠期糖尿病大網膜前脂肪細胞原代培養及增殖分化模型的建立*
2016-11-09 02:40:44潘寶龍馬潤玫
中國現代醫學雜志 2016年19期
潘寶龍,馬潤玫
(昆明醫科大學,云南 昆明 650031)
妊娠期糖尿病大網膜前脂肪細胞原代培養及增殖分化模型的建立*
潘寶龍,馬潤玫
(昆明醫科大學,云南 昆明 650031)
目的建立妊娠期糖尿病(GDM)大網膜前脂肪細胞原代培養及向成熟脂肪細胞增殖分化的模型,為脂肪細胞體外研究奠定基礎。方法使用改良的細胞培養法,以GDM剖宮產患者大網膜純脂肪組織為原材料,作前脂肪細胞的原代培養和傳代,繪制傳代前脂肪細胞的生長曲線,并對傳代前脂肪細胞做誘導分化,對已誘導分化細胞通過油紅O染色和逆轉錄聚合酶鏈反應檢測脂聯素mR NA,并進行細胞凍存和復蘇實驗。結果培養出的前脂肪細胞基本均為梭形細胞,增殖旺盛,第4天細胞開始快速增殖,倍增時間約為48h。第7天原代前脂肪細胞開始出現脂肪顆粒,而傳代細胞仍然保持梭形和無脂滴。傳代前脂肪細胞經成脂誘導,出現定向分化,經油紅O脂肪染色和脂聯素mR NA測定,證明已誘導分化為成熟脂肪細胞。結論GDM大網膜脂肪組織中具有前脂肪細胞,可進行連續傳代和大量擴增。原代前脂肪細胞在培養增殖過程中,會自然出現向成熟脂肪細胞分化,而傳代后前脂肪細胞則失去分化能力,經適當誘導后可定向分化為成熟脂肪細胞。模型的建立,可為后續脂肪細胞體外實驗研究做好前期準備。
前脂肪細胞;細胞培養;細胞模型
脂肪組織是一個功能強大的內分泌器官,可分泌瘦素、脂聯素等脂肪因子,其可能對胰島素抵抗、心血管功能、糖脂代謝、系統炎癥及骨代謝起調節作用[1-3]。目前,學術界對脂肪因子展開的生物學研究的體外實驗載體基本采用嚙齒類動物脂肪細胞[4-6]。不可忽視的一個影響因素是,嚙齒類動物與人類存在明顯的遺傳學差異,兩者的代謝途徑也截然不同。因此,將其作為實驗載體往往難以避免潛在的實驗缺陷問題,從而影響實驗數據和結論的科學性。相對而言,人源前脂肪細胞是人體中一種特異化的前體細胞,具有增殖能力和向成熟脂肪細胞定向分化的能力,在人的一生中均存在并持續作用,且與糖尿病、肥胖、動脈硬化等代謝類疾病關系密切[7]。以人源細胞作為研究載體,更具科學性,并且以取自相應疾病患者網膜組織而培養傳代的前脂肪細胞,可能更多地保留該疾病其他的基因信息,更真實地模擬疾病的基因環境,使結果更具有說服力。本研究在查閱大量文獻的基礎上,對體外脂肪細胞的培養方法做一定程度的改良,嘗試建立妊娠期糖尿病(gestational diabetes mellitus,GDM)患者大網膜前脂肪細胞體外原代培養及增殖分化模型,以期為后續的研究奠定基礎。
1 材料與方法
1.1前脂肪細胞的原代培養
實驗前,遵循醫學倫理學規定報批,并取得患者知情同意。取GDM剖宮產患者大網膜相對純凈脂肪組織10~20g,去除可見血管及纖維成分,pH 7.2磷酸鹽緩沖溶液(phosphate buffered saline,PBS)洗滌,剪成盡可能小的碎糊狀。按照1∶2體積比加入Ⅰ型膠原酶溶液(2mg/ml),5%二氧化碳CO2培養箱,37℃消化3 h,1 000 r/min離心10 min。棄上清液和漂浮的脂肪細胞,37℃紅細胞裂解液處理沉淀細胞5~10min,分別以100和200目不銹鋼網過濾。1000r/min離心10min,棄上清,將沉淀細胞加入前脂肪細胞基本培養基[杜爾伯科改良伊格爾培養基(dulbecco modified eagle medium,DMEM)],輕輕吹打懸浮細胞,調配細胞濃度為1×105個/cm3,接種于6孔板中,37℃、5%CO2培養。觀察細胞貼壁后立即進行培養基更換,并除去未貼壁細胞。每日監測,每3天進行1次培養基更換。
1.2前脂肪細胞的傳代
待細胞生長至單層融合接近80%時,可進行細胞傳代。實驗前用75%酒精擦拭超凈工作臺,并提前開啟紫外燈靜態殺菌30min。棄培養瓶內的培養基,PBS洗滌細胞2次,棄PBS液,加入2 ml胰酶37℃進行消化,待細胞稍微變圓后,立即將培養瓶豎起使胰酶與細胞分離,棄瓶中胰酶,加入新鮮培養基,并將瓶壁上的細胞吸管反復吹打混勻,調整細胞密度為1×105個/cm3,分別接種至75 cm2培養皿6和24孔板中。每3天進行1次培養基更換。當傳代細胞生長至單層融合接近80%時,可再次進行,往下傳代。
1.3繪制傳代前脂肪細胞的生長曲線
使用1個24孔板作為細胞生長曲線監測板,每2孔為1組,將24孔隨機分成12組,每天檢測1組,計數2孔各自細胞總數,取平均值,以此類推,連續計數12d,至12組結束,并繪制細胞生長曲線。
1.4傳代前脂肪細胞的誘導分化
待傳代細胞生長至單層融合接近80%時,可進行細胞誘導分化。將培養基更換為分化培養基(制備方法見附表),5%CO2培養箱中培養48 h,更換培養基為含10μg/ml胰島素的基本培養基,培養48h后,將培養基更換為基本培養基繼續培養,監測細胞內出現脂肪顆粒的數量和速度。可根據實驗需求繼續或停止培養。
1.5油紅O染色
吸棄孔板內培養液,PBS沖洗細胞3次,每孔分別加入10%甲醛磷酸鹽緩沖溶液1.5ml,在常溫下固定20 min,吸棄固定液,PBS沖洗細胞3次,60%異丙醇沖洗1次,37℃干燥5 min,每孔加入油紅O染液1.5 ml,室溫染色2 h,雙蒸水反復沖洗將油紅洗去,37℃干燥5min,置顯微鏡下觀察和拍照。
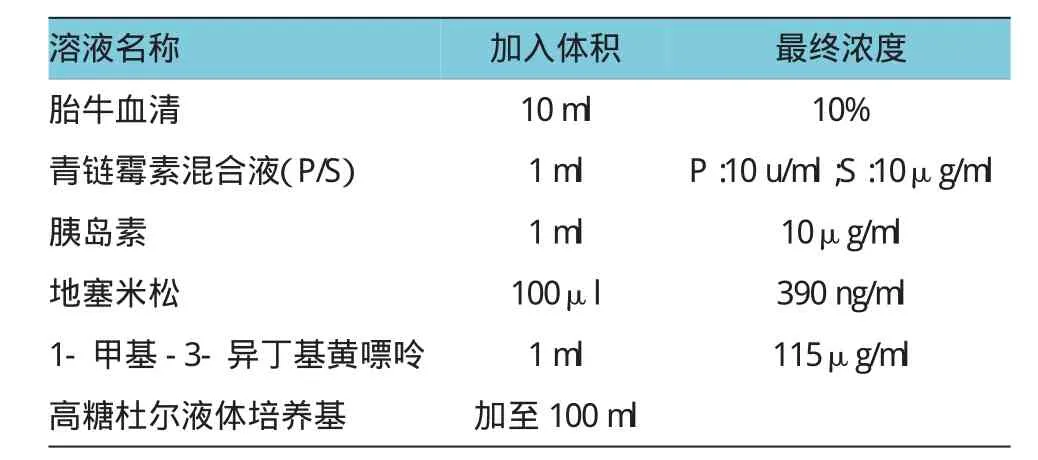
附表 分化培養基配制
1.6前脂肪細胞、成熟脂肪細胞逆轉錄聚合酶鏈反應檢測脂聯素mRNA
1.6.1R NA提取使用美國Invitrogen公司生產的Trizol試劑抽提,按照試劑盒說明書進行操作,以日本UV-2450紫外分光光度計檢測所抽提RNA含量和純度。
1.6.2聚合酶鏈反應(polymerase chain reaction,PCR)擴增和瓊脂糖凝膠電泳脂聯素mRNA引物序列:正向引物:5'-ATCGGTGAAACCGGAGTACC-3',反向引物:5'-GCATGTTGGGGATAGTAACGTAA-3',擴增產物長度148bp,體系為25μl。擴增產物以2%瓊脂糖凝膠電泳,紫外燈下判讀,凝膠成像系統拍照。
1.7細胞凍存
培養瓶內細胞經消化和吹打制成細胞懸液,1 000 r/min離心10 min,棄上清液,加入1 ml冷凍液保存(DMEM∶胎牛血清∶二甲基亞砜=8∶1∶1)吹打混勻后移入凍存管,封口,4℃放置40 min,置入-20℃冰箱冷凍2 h,再置入-80℃冰箱冷凍保存,如短期內不使用則移入液氮(-196℃)中保存。
1.8細胞復蘇
從液氮中取出冷凍管,迅速投到37℃水中,并不斷搖動(使管中的液體迅速融化),待完全溶解后取出,75%酒精擦拭凍存管外壁,超凈工作臺內取出管內液體,1000r/min離心10min,棄上清液,加入37℃、10 ml基本培養基,吹打制成細胞懸液,計數后調整濃度為1×105個/ml,分裝入培養瓶內,37℃、5%CO2培養箱中進行培養,24h后更換為新鮮培養基,每日監測,每3天進行1次培養基更換。待細胞長到培養瓶的80%后進行傳代培養。
2 結果
2.1原代培養的前脂肪細胞
培養6 h即可見部分細胞貼壁,13 h細胞貼壁率70%,24 h后基本全部貼壁,細胞呈不規則形,較為分散,數目較少。第3天觀察基本變為梭形細胞,少數呈多角形,細胞數目也有所增加。第4天細胞開始進入快速增殖,第7天原代前脂肪細胞形態出現改變,逐漸由梭形變橢圓或圓形,胞內可見脂肪顆粒,而傳代后細胞形態仍然保持梭形,局部可出現單層融合,細胞排列較為緊密。第9天發現原代前脂肪細胞內含有大量脂肪顆粒,說明大量原代細胞已自行分化為成熟脂肪細胞。而傳代細胞至第9天后仍繼續保持梭形,細胞基本單層融合,排列緊密,油紅O染色仍未著色。見圖1~3。

圖1 原代前脂肪細胞第3天(油紅O染色×100)

圖2 原代前脂肪細胞第4天(油紅O染色×100)
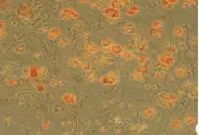
圖3 原代前脂肪細胞第9天(油紅O染色×100)
2.2傳代培養的前脂肪細胞
傳代后前脂肪細胞前4 d細胞形態及生長狀況與原代前脂肪細胞極為相似,基本為梭形,大小較為均勻。5~6d后細胞進入快速增殖,排列平行緊密。通常第7天即可完成單層融合,光鏡下觀察,細胞內無反光小滴,說明無脂肪顆粒產生,即便培養9 d后油紅O染色仍未著色(見圖4)。其連續傳代可大量擴增,但發現傳代到5代后,增殖和分化率開始降低。
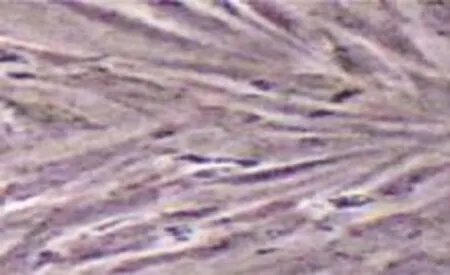
圖4 傳代前脂肪細胞第9天(油紅O染色×100)
2.3前脂肪細胞的生長曲線
以第2代傳代細胞為監測對象,使用1個24孔板作為細胞生長曲線監測板,每天隨機計數2孔細胞,取平均值,連續計數12d。見圖5。
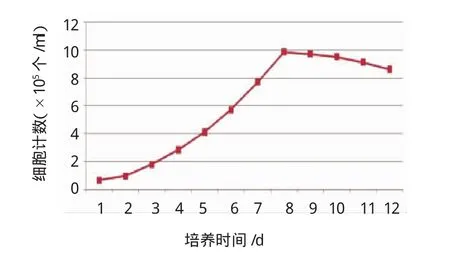
圖5 前脂肪細胞生長曲線
2.4第3代傳代前脂肪細胞誘導分化
第3代傳代細胞誘導后,出現向成熟脂肪細胞的定向分化。第6天即可見部分細胞開始出現單個散在反光小滴。隨培養時間延長,細胞內脂滴逐漸增多,第12天可見胞內已聚集大量脂滴,經油紅O染色初步證實分化成功。見圖6、7。
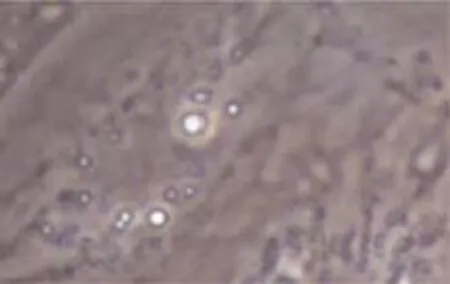
圖6 分化第6天(油紅O染色×400)

圖7 分化第12天(油紅O染色×400)
2.5逆轉錄聚合酶鏈反應
前脂肪細胞無脂聯素mRNA表達,成熟脂肪細胞有脂聯素mRNA表達。見圖8。

圖8 逆轉錄聚合酶鏈反應
3 討論
在傳統醫學中,脂肪組織僅被認為是人體儲存脂質的場所,在人體能量不足的情況下,受刺激而釋放能量。直到上世紀90年代,學者們發現脂肪瘦素的存在,傳統醫學對脂肪的認知才得以改變。脂肪組織不但可以作為人體能量的儲存“庫房”,同時其自身也具有活躍的內分泌功能,包括脂肪因子等,在人體能量穩態及調控糖、脂代謝平衡的維持中,發揮著重要作用。近年來,針對脂肪因子的研究越來越受到學術界的關注[8-10],因涉及醫學倫理學原因,許多研究只能是局限于人體血清學和體內組織學的表象研究,而進一步更深層次的研究,如對于基因表達(過表達或沉默)與疾病的關系研究,只能以體外實驗方式進行,常見的是細胞培養和動物模型。體外前脂肪細胞的培養,對于研究脂肪細胞、脂肪因子、相應細胞因子等與相關疾病的關系,具有重要的作用。同時,作為理想的脂肪細胞研究模型,有利于觀察各種相關因素在脂肪組織發生、增殖及肥大整個演變過程中的調控作用,有助于對相關機制進行深入研究。
近年來,隨著對人體脂肪組織大量研究工作的開展,對前脂肪細胞的原代培養方法、分離獲取能力也提出更高的要求。學者們的培養方法不盡一致,各操作者自身的經驗不一,實驗室的條件也不盡相同,因而得出的結果和經驗也各具特點[11-13]。對于人源前脂肪細胞培養,目前采用較多的是血管基質組分細胞培養法,操作較為簡便,但最大的缺點是細胞的增殖和分化速度較慢,培養基中雜質較多,并且容易出現細胞大小不均,導致后期實驗結果容易受到影響[12-13]。GARCIA等[14]曾提出細胞島學說,認為細胞島存在于成熟脂肪組織中,島內細胞均為前脂肪細胞,具有高增殖速度和高分化能力,是脂肪組織的發源地,對脂肪組織的增生和肥大起至關緊要的作用。本研究筆者基于上述學說,總結前人經驗,將天花板培養與脂肪組織塊貼壁的方法相結合,著重加強和改良消化方式,將多數文獻使用的消化方式(37℃、1 mg/mlⅠ型膠原酶,30~120 min)改良為按照1∶2體積比加入Ⅰ型膠原酶溶液(2 mg/ml),5%CO2培養箱,37℃消化3h,增加消化液使用量和濃度,延長消化時間,并且使用紅細胞裂解液37℃處理沉淀細胞5~10min,以去除殘留紅細胞的污染,從而盡可能地達到較好地消化效果。實踐證明上述改良是較為成功的,所得到培養細胞成分均一,數量較多,增殖較為旺盛,可能延長消化時間,更完全地消化成熟脂肪細胞,也更多地分離出細胞島中前脂肪細胞。
對于前脂肪細胞的判別標準,有學者提出必須滿足3個條件[15-16]:①細胞為經典的梭形形態,脂肪組織來源,胞漿內極少或無脂肪顆粒;②細胞增殖速度與同一個體的成纖維細胞類似,增倍時間短;③胰島素促進下,細胞在單層融合后能形成成熟脂肪細胞,胞內可見脂肪滴,或者能檢出特異性酶或脂聯素。本研究取材于剖宮產GDM患者大網膜組織,經過消化、過濾、離心等系列處理后得到一些長梭形細胞,光鏡下胞漿內不見反光小滴,也不能被油紅O著色,培養之后細胞增殖快速,從細胞生長曲線可以看出,倍增時間約為48h,控制條件下可進行傳代培養,傳代細胞經誘導后可出現分化,經過油紅O染色,初步證實分化成功。脂聯素是目前國內外已經公認的成熟脂肪細胞特有的細胞因子[17-19],運用逆轉錄聚合酶鏈反應對誘導分化前后的細胞進行脂聯素mRNA表達檢測,發現誘導分化后的細胞有脂聯素mRNA的表達,而未經誘導的傳代細胞未能檢測到脂聯素mRNA,從而證實誘導分化后的細胞為成熟脂肪細胞。而本實驗從網膜組織中分離得到的細胞,上述3個條件,是人源原代前脂肪細胞。
值得一提的是,本實驗中筆者在原代和傳代培養中發現,原代前脂肪細胞第7天開始有部分細胞形態發生改變,逐漸由梭形變橢圓乃至圓形,并且光鏡下胞內可見脂肪顆粒,第9天發現原代前脂肪細胞內含有大量脂肪顆粒,說明大量原代細胞已自行分化為成熟脂肪細胞。而傳代后前脂肪細胞直至第12天仍繼續保持梭形,油紅O染色仍未著色,必須經過誘導,才會出現細胞分化。本實驗中經過多次重復培養,仍然出現該現象,證實GDM原代前脂肪細胞在體外培養過程中,會自然出現向成熟脂肪細胞分化,并不需要外加高濃度胰島素和地塞米松,而傳代后前脂肪細胞則出現失去分化的能力。原代前脂肪細胞會出現自然分化,無法傳代,可能與該現象有關。而該現象的發生,可能與原代細胞來源于體內成熟脂肪組織,在體內可能已經被激活,或者仍然會攜帶某些體內分化因子,與外加的誘導劑作用相類似[20]。而傳代后前脂肪細胞出現失去分化的能力,可能是隨著不斷傳代,與人體內環境比較,體外培養環境出現越來越大的偏差,導致傳代細胞發生一定程度生物學特性的變化,并且該變化隨傳代數增加而表現越發明顯,具體表現為增殖和分化能力的不斷下降。本研究結果也證實,通常傳代在5代以內時,細胞的增殖及分化能力尚可,再往后傳代則增殖和分化率開始降低。本實驗后期所做細胞凍存和復蘇實驗中,也發現冷凍保存的細胞最佳選擇為傳代第2代或第3代細胞,并且只宜凍存復蘇1次。同樣涉及到細胞增殖和分化能力降低的問題,也從一個側面說明本實驗目前所使用凍存和復蘇技術還存在一定的缺陷,有待進一步改進。
綜上所述,GDM大網膜脂肪組織中具有前脂肪細胞,可進行連續傳代和大量擴增。經適當誘導后可定向分化為成熟脂肪細胞。本課題組成功建立GDM患者大網膜前脂肪細胞原代培養及增殖的模型,為脂肪組織體外細胞學研究、脂肪因子體外表達研究、疾病發病機理等實驗研究奠定基礎。
[1]BAUMGARD L H,HAUSMAN G J,SANZ FERNANDEZ MV,et al.Insulin:pancreatic secretion and adipocyte regulation[J].Domest Anim Endocrinol,2016,54(1):76-84.
[2]MAO J,BATH S C,VANDERLELIE J J,et al.No effect of modest selenium supplementation on insulin resistance in UK pregnant women,as assessed by plasma adiponectin concentration[J]. Br J Nutr,2016,115(1):32-38.
[3]GARCIA-CARRIZO F,NOZHENKO Y,PALOU A,et al.Leptin effect on acetylation and phosphorylation of pgc1alpha in muscle cells associated with ampk and akt activation in high-glucose medium[J].J Cell Physiol,2016,231(3):641-649.
[4]聶緒強,楊建文,史海霞,等.IR-3T3-L1脂肪胰島素抵抗細胞的建立[J].南方醫科大學學報,2015,35(1):103-108.
[5]PRIETO-HONTORIA P L,PEREZ-MATUTE P,FERNANDEZGALILEA M,et al.Effects of alpha-lipoic acid on chemerin secretion in 3T3-L1 and human adipocytes[J].Biochim Biophys Acta,2016,1861(3):260-268.
[6]WANG C,WANG L,LI W,et al.Irisin has no effect on lipolysis in 3T3-L1 adipocytes or fatty acid metabolism in HepG2 hepatocytes[J].Endocrine,2015,49(1):90-96.
[7]LUJAN-HERNANDEZ J,LANCEROTTO L,NABZDYK C,et al. Induction of adipogenesis by external volume expansion[J].Plast Reconstr Surg,2016,137(1):122-131.
[8]BIL-LULA I,SOCHOCKA M,ZATONSKA K,et al.Adenovirus type 9 enhances differentiation and decreases cytokine release from preadipocytes[J].J Med Virol,2015,87(2):230-239.
[9]徐曉麗,丁攀,李曉霞,等.瘦素、脂聯素與代謝綜合征相關指標的關系[J].寧夏醫科大學學報,2016,38(2):169-171.
[10]WURST U,EBERT T,KRALISCH S,et al.Serum levels of the adipokine Pref-1 in gestational diabetes mellitus[J].Cytokine,2015,71(2):161-164.
[11]閆美玲.LOX-1在棕櫚酸誘導前脂肪細胞死亡、炎癥及脂肪生成中的作用及機制[D].長沙:中南大學,2014.
[12]高鵬.生長激素與其信號分子STAT5B對脂肪細胞分化的影響[D].濟南:山東大學,2015.
[13]張文瑾,孫建軍.人脂肪間充質干細胞的分離及培養和鑒定[J].臨床耳鼻咽喉頭頸外科雜志,2016,24(9):57-60.
[14]GARCIA de HERREROS A,BIRNBAUM M J.The acquisition ofincreased insulin-responsive hexose transportin3T3-L1 adipocytes correlates with expression of a novel transporter gene[J]. J Biol Chem,1999,264(33):19994-19999.
[15]JANKOVIC A,KORAC A,SRDIC-GALIC B,et al.Differences in the redox status of human visceral and subcutaneous adipose tissues-relationships to obesity and metabolic risk[J].Metabolism, 2014,2(14):S0026-S0034.
[16]FERNANDEZ-TRASANCOS A,GUEROLA-SEGURA R,PARADELA-DOBARRO B,et al.Glucose and inflammatory cells decrease adiponectin in epicardial adipose tissue cells:paracrine consequences on vascular endothelium[J].J Cell Physiol,2016, 231(5):1015-1023.
[17]BAO W,BAECKER A,SONG Y,et al.Adipokine levels during the first or early second trimester of pregnancy and subse quent risk of gestational diabetes mellitus:a systematic review[J]. Metabolism,2015,64(6):756-764.
[18]WHITE U A,MAIER J,ZHAO P,et al.The modulation of adiponectin by STAT5-activating hormones[J].Am JPhysiol Endocrinol Metab,2016,310(2):E129-E136.
[19]CAWTHORN W P,SCHELLER E L,PARLEE S D,et al.Expansion of bone marrow adipose tissue during caloric restriction is associated with increased circulating glucocorticoids and not with hypoleptinemia[J].Endocrinology,2016,157(2):508-521.
[20]ABUNA R P,de OLIVEIRA F S,SANTOS TDE S,et al.Participation of TNF-alpha in inhibitory effects of adipocytes on osteoblast differentiation[J].J Cell Physiol,2016,231(1):204-214.
(童穎丹編輯)
Establishment of primary culture,proliferation and differentiation model of GDM preadipocytes*
Bao-long Pan,Run-mei Ma
(Kunming Medical University,Kunming,Yunnan 650031,China)
Objective To establish the model of primary culture,proliferation and differentiation of the gestational diabetes mellitus(GDM)epiploon preadipocytes,so as to lay the foundation for the follow-up study. Methods Using improved cell culture method,the pure fat tissue of greater omentum from GDM patients having cesarean section was used for the primary culture and passage.The growth curve of the preadipocytes was drawn.The adipocytes differentiated from preadipocytes after induction were determined by oil red O staining,and their adiponectin mRNA level was detected with RT-PCR to confirm the differentiation,and then the experimentsofcellcryopreservationandresuscitationwerecarriedout.ResultsTheprimarycultured preadipocytes were almost spindle cells with strong proliferation capacity.They began to proliferate rapidly on the 4th day,the doubling time was about 48 h.On the 7th day,the original generation of preadipocytes began to have fat particles,and the passage cells remained to be spindle without lipid drops.After induction,the passage preadipocytes showed oriented differentiation,oil red O staining and adiponectin mRNA measurement proved that the cells had differentiated into mature adipocytes.Conclusions Preadipocytes exit in GDM mature adipose tissue,can be continuously passaged and greatly amplified.In the culture progress,the original generation of preadipocytes will naturally differentiate into mature adipocytes,but the passaged preadipocyteslose the ability of differentiation,after appropriate induction,they can differentiate into mature fat cells. Keywords:preadipocyte;cell culture;cell model
R 714.255;R 589.2
A
10.3969/j.issn.1005-8982.2016.19.003
1005-8982(2016)19-0011-06
2016-05-23
國家自然科學基金(No:81160082)
馬潤玫,E-mail:pbl6916@163.com
