基于cox1和D-loop區序列雷州半島紅樹林海區2種野生彈涂魚的遺傳變異
2016-11-12 03:41:48沈自慧黃永勝龍水生黃承勤王中鐸
廣東海洋大學學報 2016年4期
關鍵詞:分析
廖 健,賀 亮,張 順,沈自慧,黃永勝,龍水生,黃承勤,王中鐸
(廣東海洋大學水產學院,南海水產經濟動物增養殖廣東普通高校重點實驗室,廣東 湛江 524025)
基于cox1和D-loop區序列雷州半島紅樹林海區2種野生彈涂魚的遺傳變異
廖健,賀亮,張順,沈自慧,黃永勝,龍水生,黃承勤,王中鐸
(廣東海洋大學水產學院,南海水產經濟動物增養殖廣東普通高校重點實驗室,廣東湛江524025)
設計雙層洞穴式彈涂魚捕捉器,于雷州半島紅樹林海區捕獲彈涂魚類野生群體,篩選出彈涂魚(Periophthalmus modestus)、大彈涂魚(Boleophthalmus pectinirostris)2個優勢種群,并獲取其線粒體控制區(D-loop)和cox 1基因部分序列,進行序列變異分析及遺傳分析。結果表明,cox 1基因和D-loop區堿基組成A+T含量均大于C+G。cox 1數據揭示,大彈涂魚變異位點占總長度2.1%,彈涂魚占2.4%,種內平均遺傳距離分別為0.003、0.006,種間遺傳距離為0.158,37個個體共定義22個單倍型,單倍型多樣性為0.950 45。而D-loop區數據指明,大彈涂魚變異位點占總長度4.9%,彈涂魚占7.5%,種內平均遺傳距離分別為0.012、0.019,種間距離為0.409,37個個體定義32個單倍型,單倍型多樣性水平極高(0.989 49)。中性檢驗結果顯示,2種彈涂魚進化歷程符合中性進化假設,且存在群體擴張的可能。綜合cox 1基因和控制區數據顯示:彈涂魚種群遺傳多樣性高于大彈涂魚。
紅樹林;彈涂魚;捕捉器;序列變異;系統進化
彈涂魚類(mudskipper)隸屬于蝦虎魚科背眼蝦虎魚亞科,我國常見的有大彈涂魚(Boleophthalmus pectinirostris)、彈涂魚(Periophthalmus modestus)、青彈涂魚(Scartelaos histophorus),主要分布于廣東、廣西、海南等地的河口、潮溝及紅樹林海區附近灘涂[1]。彈涂魚類在物種進化上處于極重要位置,是一類進化程度較低的兩棲穴居魚類。彈涂魚類有較高的經濟價值和營養價值,同時是近岸灘涂環境監測的重要指示生物[2]。
細胞色素C氧化酶I(cox 1或COI)基因,是線粒體中為數不多的氨基酸編碼基因之一[3]由于其結構簡單、分子小、進化速率適中以及遵循母系遺傳等特征[4-6],被用來分析群體間遺傳結構及遺傳多樣性及系統進化關系,逐漸應用于各類物種的分類鑒定。廖健等[7]利用cox 1基因部分序列作為分子標記,分析了雷州半島紅樹林區5種蝦虎魚類的系統進化關系,并指出cox 1可用來發現潛在種。線粒體D-loop區又稱控制區或D環區,是線粒體中進化速率最快的一段不編碼DNA片段,常用于遺傳結構分析,在魚類遺傳分析方面,已應用于南海金槍魚(Thunnus albacares和T.obesus)和大眼鯛(Priacanthus macracanthus)等[8-9]。關于彈涂魚類的研究主要集中于大彈涂魚養殖群體,而對于彈涂魚類野生群體僅見零星的報道[2,10-14]。筆者利用模擬彈涂魚生境設計的捕捉裝置采集野生彈涂魚類樣本,通過獲取線粒體cox 1基因和控制區部分序列,分析彈涂魚類的遺傳變異,為雷州半島彈涂魚類野生資源的生物學保護和合理開發提供參考。
1 材料與方法
1.1樣本的采集
根據彈涂魚特有的灘涂鉆孔棲息習性,模擬其生境,采用塑化玻璃防腐蝕材質制作雙層梯形單向通道洞穴式彈涂魚自動捕捉裝置(圖1),利用該捕捉器采集彈涂魚野生群體。2015年11月至2016年3月,于雷州半島高橋紅樹林保護區(N21°36'24",E109°47'8")捕獲彈涂魚類一批,鮮活運回實驗室,經鑒定、篩選,用于群體分析的大彈涂魚(B.pectinirostris,BP)19尾、彈涂魚(P.modestus,PM)18尾。
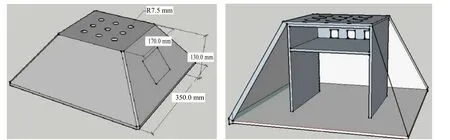
圖1 雙層洞穴式彈涂魚捕捉器Fig.1 Double cave mudskipper catcher
1.2DNA提取、cox 1和D-loop擴增及測序
鮮活彈涂魚樣本用體積分數75%酒精處理,剪取肌肉組織約0.5g,采用傳統苯酚-氯仿法提取基因組DNA[15]。cox 1基因的擴增引物參照孟瑋等[16],引物正、反向序列分別為:Fish F:5′-TCAACCAAC CACAAAGACAATGGCAC-3′;Fish R:5′-TAGA CTTCTGGGTGGCCAAAGAATCA-3′。控制區的擴增的正、反向引物為:D-loop F:5′-CCCATCTCT AGCTCCCAAAGC-3′,D-loop R:5′-CTGTAGAGT GAACGCTTGGCATG -3′[17]。引物由上海生工生物有限公司合成。PCR反應體系:10×Buffer(含Mg2+) 2.5μL、2.5 mmol/L Mix dNTP 2μL、5μmol/L引物各1μL、DNA模板1μL、500 u/L Taq DNA聚合酶0.2μL,補充ddH2O至25μL。程序為:94℃ 1min 20 s;94℃ 30 s,T℃ 30 s,72℃ 50 s,30個循環;72℃下延伸5min;4℃下保存,其中cox 1擴增退火溫度T為55℃,控制區擴增退火溫度T為54℃。PCR擴增用Hema9600擴增儀進行,產物經10 mg/mL瓊脂糖凝膠電泳檢測,選取條帶明亮單一的產物送至上海生工生物有限公司測序。
1.3數據處理
原始序列數據經MEGA6.0多重序列比對、人工核查校正,統計序列基本信息,包括變異位點、單一突變位點、簡約信息位點等[18]。堿基組成由MEGA6中Compute Nucleotide Composition計算,Kimura 2-parameter種內、種間遺傳距離分別由Within Group Mean Distance和Between Group Mean Distance程序計算。利用Modeltest選擇適合進化模式[19],構建 Neighbor-Joining系統發育樹,其中Bootstrap 重復抽樣1 000次。單倍型數、單倍型多樣性、核苷酸多樣度、平均核苷酸差異及中性檢驗的Tajima's D和Fu's Fs值等遺傳多樣性參數均由DnaSP v5計算[20],群體擴張的堿基不配對分布曲線由DnaSP v5中population size change程序完成。最終的序列遞交由 Sequin編輯,并 Email至gb-admin@ncbi.nlm.nih.gov獲取登錄號。
2 結 果
2.1序列多態性分析
Sanger法測序獲得的片段經比對剪切后,獲得大彈涂魚cox 1部分序列633 bp,D-loop區部分序列872 bp;彈涂魚cox 1部分序列634 bp,D-loop區部分序列454 bp;所有序列遞交至NCBI,獲取登錄 號 (cox 1:KX263765-KX263801,D-loop:KX263728-KX263747)。堿基組成如表1。cox 1基因部分序列中,堿基T在密碼子的第1、2和第3位具有明顯的偏倚性,明顯偏向堿基第3位,其中大彈涂魚A+T含量(56.1%)大于C+G含量(43.9%),彈涂魚A+T含量(57.2%)亦大于C+G含量(42.8%)。D-loop區部分序列A、T、C、G 4種堿基在密碼子的第1、2和3位并未出現明顯的偏倚性,大彈涂魚A+T含量(64.9%)大于C+G含量(35.1%),彈涂魚A+T含量(64.0%)亦大于C+G含量(36.0%)。經多態性分析,cox 1數據揭示,633 bp的大彈涂魚cox 1部分序列中,保守位點620個,變異位點13個,變異率為2.1%,簡約信息位點5個,單一突變位點8個;634 bp的彈涂魚cox 1部分序列中,保守位點619個,變異位點15個,變異率為2.4%,簡約信息位點9個,單一突變位點6個。D-loop結果表明:872 bp的大彈涂魚D-loop區部分序列中,保守位點828 bp,變異位點43個,變異率達4.9%,簡約信息位點17個,單一突變位點26個;454 bp的彈涂魚D-loop區部分序列中,保守位點420個,變異位點34個,變異率達7.5%,簡約信息位點17個,單一突變位點17個。
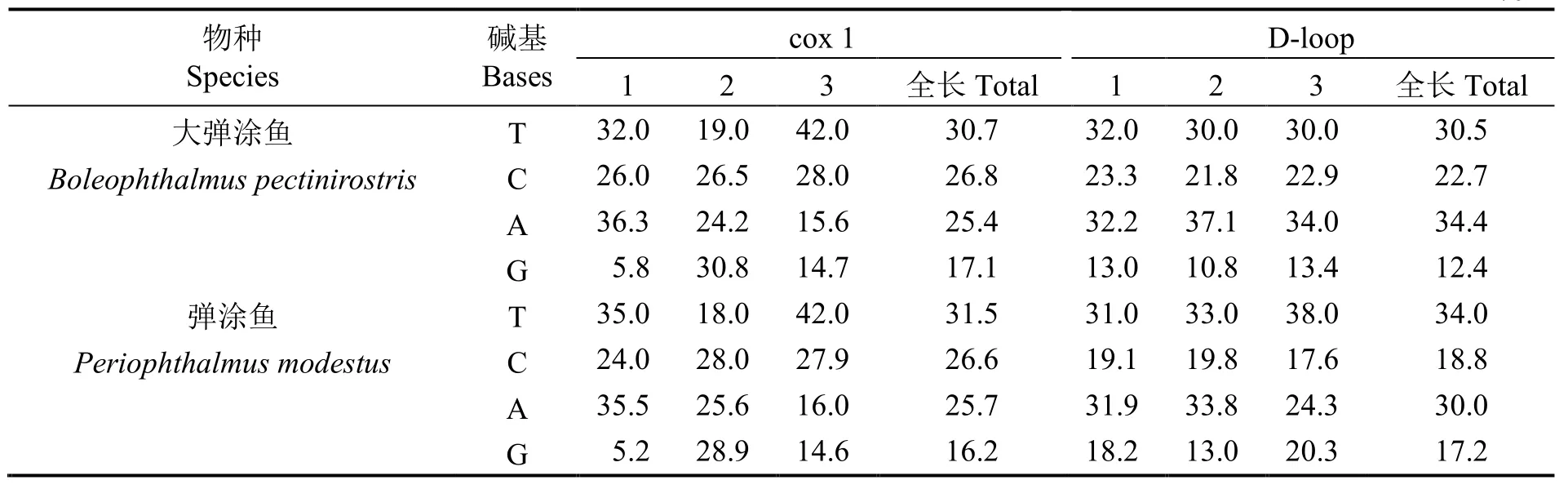
表1 基于cox 1基因及控制區部分序列的2種彈涂魚堿基組成Table 1 Nucleotide composition of 2 species of mudskipper based on cox 1 gene and D-loop region partial sequences %
2.2遺傳距離與聚類分析
Kimura 2-parameter模型的種內種間平均遺傳距離結果如表2所示:基于cox 1基因部分序列的2種彈涂魚類種間平均遺傳距離為0.158,其中大彈涂魚種內平均遺傳距離為 0.003,彈涂魚種內平均遺傳距離為0.006;而基于D-loop區部分序列的2種彈涂魚類種間平均遺傳距離較cox 1高,為0.409,種間平均遺傳也均高于以cox 1基因計算的結果,其大彈涂魚、彈涂魚種間平均遺傳距離分別為0.012、0.019。以湖棲鰭蝦虎魚(Gobiopterus lacustris)為外群分別構建2種彈涂魚類cox 1基因和控制區的NJ聚類圖(圖2),由圖2可見,基于cox 1基因及控制區部分片段構建的NJ進化樹均聚為2大分類群,且均與外群物種分離。

表2 基于cox 1基因和D-loop的彈涂魚類種內種間遺傳距離Table 2 Intraspecific genetic distance of mudskipper based on cox 1 gene and D-loop region partial sequences
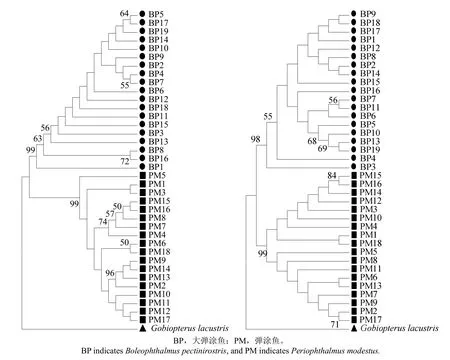
圖2 基于cox 1基因(左)及控制區(右)部分序列的構建的2種彈涂魚NJ聚類樹Fig.2 Neighbor-Joining tree of 2 species of mudskipper based on cox 1 gene(left)and D-loop region(right)partial sequences
2.3遺傳變異
所統計的遺傳多樣性參數如表3。37個彈涂魚個體中,基于cox1基因部分序列共檢測出22個單倍型,總單倍型多樣性處較高水平(0.950 45),總核苷酸多樣度為 0.074 77,總核苷酸差異平均為47.103 60。基于D-loop序列統計的各項遺傳多樣性參數值多高于cox1基因,37個個體中存在32個單倍型,單倍型比例高達84.5%,總單倍型多樣性處極高水平(0.989 49),總核苷酸多樣度也較 cox1基因高(0.168 32),總核苷酸差異平均為75.070 57。基于cox 1和控制區部分序列的分析結果均表明,彈涂魚群體遺傳多樣性較大彈涂魚高。
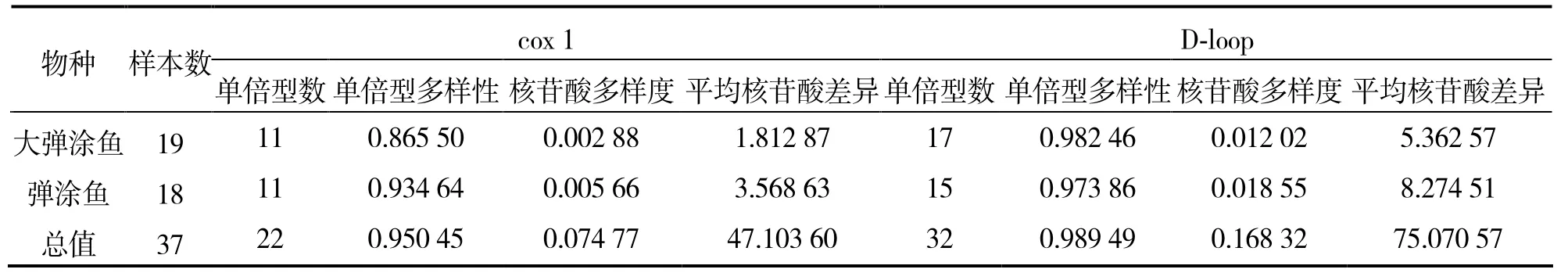
表3 基于cox 1基因及D-loop區部分序列的雷州半島群體遺傳多樣性參數Table 3 Parameters of genetic diversity in Leizhou Peninsula populations based on cox 1 gene and D-loop partial sequence
2.4中性檢驗與群體擴張
中性檢驗結果顯示,Tajima's D和Fu's Fs值均為負數,均不顯著,表明2個彈涂魚類種群在進化上符合中性進化假設。基于cox 1基因和控制區部分序列的2種彈涂魚群體增長均服從constant模型,核苷酸不配對分布曲線均呈現單峰形(圖3),表明2種彈涂魚類近期可能經歷了群體擴張過程。

表4 基于cox 1基因和D-loop區部分序列的中性檢驗參數Table 4 Parameters of neutrality test based on cox 1 gene and D-loop region sequence
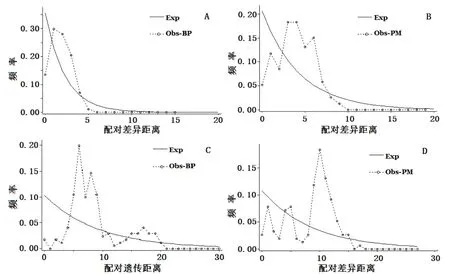
圖3 基于cox 1基因及D-loop區序列的2種彈涂魚mismatch分布分析Fig.3 Mismatch distribution for all pairwise combinations of mitochondrial cox 1 gene and control region data sets of 2 species of mudskipper from Leizhou Peninsula
3 討 論
3.1cox 1基因和控制區在群體遺傳分析上的應用
真核生物線粒體環狀DNA上的多個基因或片段根據其變異程度及速率不同而具有不同的遺傳分析潛力。例如,較為保守的cox 1、Cyt b、16s rRNA基因除作為Barcoding輔助物種鑒定外,還用于群體系統關系分析。而變異速率較快的控制區(D-loop)則常用于種內群體間遺傳多樣性分析。李玉龍等[21]利用cox 1基因分析遼寧沿海不同體色花紋的細紋鰤子魚(Liparis tanakae)二野生群體遺傳多樣性和遺傳結構時表明,cox 1適用于種內遺傳分析,也是本研究應用cox 1的原因之一。董麗娜等[22]通過 cox 1基因聚類分析推測日本金線魚(Nemipterus japonicus)與紅棘金線魚(N.nemurus)為同一物種,而確定蘇門答臘金線魚(N.mesoprion)與日本金線魚為不同物種,分析三者間的遺傳變異時揭示,cox1基因可快速有效地鑒定物種,亦可在一定程度上檢測種群間或種群內的變異。線粒體控制區在群體遺傳上的應用也受到普遍關注,王中鐸等[8]測定南海海域大眼金槍魚和黃鰭金槍魚控制區序列,分析了2種金槍魚的遺傳結構,并與GenBank中印度洋、大西洋、太平洋數據比較分析,分析不同海域黃鰭金槍魚與大眼金槍魚群體間的基因交流情況及群口擴張。熊丹等[9]利用控制區分析了南海短尾大眼鯛的6個地理群體,發現其間并未出現明顯的分化現象。本研究在通過 cox1基因對野生彈涂魚類進行準確定種基礎上,利用 cox1基因分析彈涂魚、大彈涂魚的遺傳結構,結合控制區分析數據,發現2種彈涂魚線粒體控制區所檢測出的變異位點與保守位點比值均高于 cox1基因,這與彭士明等[3]研究銀鯧的養殖與野生群體的結果相似。遺傳距離分析表明,基于線粒體控制區的種內種間遺傳距離均大于利用 cox1基因所統計的遺傳距離值,而基于 cox1基因的彈涂魚與大彈涂魚之間遺傳距離則較小(0.158),種內遺傳距離同樣是D-loop區所計算的數值較cox1基因大(表2)。同時,基于線粒體控制區的2種彈涂魚單倍型數目、單倍型比例、單倍型多樣性、核苷酸多樣度、平均核苷酸差異均高于 cox1基因(表3)。中性檢驗表明,2種彈涂魚均符合中性進化假設,且均經歷過群體擴張過程。綜上,cox1基因和線粒體控制區均可用于群體遺傳分析,但 cox1基因在分析上不及線粒體控制區敏感。
3.22種彈涂魚類群體遺傳特征
大彈涂魚和彈涂魚是雷州半島沿岸廣泛分布的2種彈涂魚類,具有重要的生物進化研究價值和經濟價值[23]。本研究中,綜合cox1數據與D-loop區結果,2種彈涂魚中大彈涂魚遺傳多樣性水平相對較低,這與金春華等[14]利用RAPD技術研究的結果類似,雖然金春華等早年的研究技術(RAPD)未能從分子水平定量評價大彈涂魚野生群體的遺傳特征,且選擇的隨機引物數量也極有限,但仍有一定參考價值,筆者對雷州半島大彈涂魚線粒體cox1和D-loop區的研究則深入探討了大彈涂魚遺傳水平,準確地從分子水平定性定量闡述大彈涂魚種群現狀。楊帆等[2]在利用cox1基因作為DNA條形碼鑒定彈涂魚類同時亦指出,我國東南沿海3種彈涂魚類中,青彈涂魚(S.histophorus)遺傳多樣性最低,其次是大彈涂魚,而彈涂魚則最高,與本研究結果一致,進一步驗證了彈涂魚遺傳多樣性高于大彈涂魚的結果。主要原因可能是,人們對個體大、營養豐富、經濟價值高的野生大彈涂魚種群的捕獲量大,加之日益嚴重的環境問題,導致大彈涂魚種群數量減少,群體間基因交流減弱,種群退化。
4 結 論
1)cox 1基因和控制區(D-loop)序列均可用于彈涂魚、大彈涂魚等魚類系統發生及遺傳分析,但cox 1不及控制區敏感。
2)基于cox 1和控制區(D-loop)的彈涂魚、大彈涂魚種群綜合分析表明,彈涂魚遺傳多樣性高于大彈涂魚,種群遺傳結構較大彈涂魚穩定。
[1]伍漢霖,鐘俊生.中國動物志[M].北京:科學出版社,2008:1-14.
[2]楊帆,何利軍,雷光春,等.中國東南沿海彈涂魚科常見魚類的遺傳多樣性和 DNA條形碼[J].生態學雜志,2012,31(3):676-683.
[3]彭士明,施兆鴻,侯俊利.基于線粒體D-loop區與COI基因序列比較分析養殖與野生銀鯧群體遺傳多樣性[J].水產學報,2010,34(1):19-25.
[4]陳子安,杜曉東,王慶恒,等.3種星蟲線粒體 16S rRNA、COI和Cyt b基因片段的序列比較[J].廣東海洋大學學報,2007,27(4):3-10.
[5]朱惠敏,連一陽,王中鐸,等.基于cox1條形碼的魚肚DNA分子鑒定[J].廣東海洋大學學報,2014,34(6):1-5.
[6]位正鵬,孔曉瑜,吳相云,等.基于 COI序列的翡翠股貽貝 Perna viridis線粒體遺傳特性分析及其近緣種間的系統關系探討[J].熱帶海洋學報,2009,28(6):72-78.
[7]廖健,張順,龍水生,等.5種蝦虎魚類線粒體COI基因序列變異及系統進化[J].廣東海洋大學學報,2016,36(1) :7-12.
[8]王中鐸,郭昱嵩,顏云榕,等.南海大眼金槍魚和黃鰭金槍魚的群體遺傳結構[J].水產學報,2012,36(2):191-201.
[9]熊丹,李敏,陳作志,等.南海短尾大眼鯛的種群遺傳結構分析[J].南方水產科學,2015,11(2):27-34.
[10]洪萬樹,張其永,王明雄,等.人工管道誘導大彈涂魚產卵研究[J].熱帶海洋學報,2001,20(3):75-80.
[11]張其永,洪萬樹,陳仕璽,等.潮間帶大彈涂魚產卵習性和自然受精卵孵化研究[J].熱帶海洋學報,2006,25(5):68-73.
[12]劉至治,楊金權,王正琦,等.長江口及其南部鄰近地區大彈涂魚種群遺傳結構及種群歷史分析[J].動物學研究,2009,30(1):1-10.
[13]陳維,洪萬樹,陳仕璽,等.西北太平洋大彈涂魚群體遺傳結構的 AFLP分析[J].中國水產科學,2014,25(5):1020-1028.
[14]金春華,鐘愛華,黃福勇,等.大彈涂魚自然種群遺傳多樣性的 RAPD分析[J].海洋科學,2004,28(12):26-30.
[15]SAMBROOK J,FRITSCH E P,MANIATIS T.Molecular cloning:a laboratory manual[M].Cold Spring Harbor:Cold Spring Harbor Laboratories Press,1989.
[16]孟瑋,楊天燕,海薩,等.基于線粒體 COI基因序列的亞東鮭 DNA條形碼研究[J].水產學雜志,2010,23(1):2010-2014.
[17]宋娜,宋林,高天翔,等.斑尾復蝦虎魚群體遺傳多樣性比較分析[J].水產學報,2011,35(3):321-326.
[18]TAMURA K.MEGA5:molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance,and maximum parsimony methods[J].Molecular Biology and Evolution,2011,28(10):2731-2739.
[19]POSADA D,CRANDALL K A.Modeltest:testing the model of DNA substitution[J].Bioinformatics,1998,14(9):817-818.
[20]LIBRADO P,ROZAS J.DNA SP V5:a software for comprehensive analysis of DNA polymorphism data[J].Bioinformatics,2009,25(11):1451-1452.
[21]李玉龍,劉俢澤,李軼平,等.基于線粒體 COI基因序列的遼寧沿海細紋鰤子魚群體遺傳多樣性分析[J].海洋漁業,2016,38(2):120-129.
[22]董麗娜,黃梓榮,艾紅,等.北部灣3種金線魚類COI基因序列的比較分析[J].中國水產科學,2011,18(3) :508-514.
[23]王從濤.中國沿海大彈涂魚群體的親緣地理與種群遺傳結構分析[D].上海:上海海洋大學,2013.
(責任編輯:劉慶穎)
Genetic Variation of 2 Wild Mudskipper Populations from Mangroves of Leizhou Peninsula Based on cox 1 Gene and Control Region
LIAO Jian,HE Liang,ZHANG Shun,SHEN Zi-hui,HUANG Yong-sheng,LONG Shui-sheng,HUANG Cheng-qin,WANG Zhong-duo
(Key Laboratory of Aquaculture in South China Sea for Aquatic Economic Animal of Guangdong Higher Education Institutes,Fisheries College,Guangdong Ocean University,Zhanjiang 524025,China)
Double-cavern trap were designed to capture wild mudskipper populations that could screen Periophthalmus modestus and Boleophthalmus pectinirostris populations to analyze sequence variation and genetic structure based on mitochondrial control region (D-loop) and cox 1 gene sequence.The results showed that base composition of A + T was greater than that of C + G,which occurred on both cox 1 gene and the D-loop region.The data of cox 1 revealed 2.1% variation sites of B.pectinirostris,while P.modestus hold 2.4%.The average genetic distance of intraspecific was 0.003,0.006,respectively,and the interspecific was 0.158.Besides,37 individuals defined a total of 22 haplotypes based on cox 1 gene,while haplotype diversity was 0.950 45.The data of D-loop region indicated that B.pectinirostris hold 4.9% variation sites of the total length and P.modestus had 7.5%.The average genetic distances of intraspecific and interspecific were 0.012,0.019 and 0.409,respectively.Furthermore,37 individuals contained 32 haplotypes based on D-loop region.Haplotype diversity was at a very high level (0.989 49).Neutral test results showed that evolution hypothesis of the two kindsmudskippers were consistent with neutral evolution,and population expansion might exist.Considering all these facts,it can be inferred that the genetic diversity of P.modestus was higher than B.pectinirostris.
Mangroves;mudskipper;trapper;Sequence variation;Phylogenetic
S917.4
A
1673-9159(2016)04-0006-06
10.3969/j.issn.1673-9159.2016.04.002
2016-05-28
廣東海洋大學“海之帆—起航計劃”大學生科技創新項目(hzfqhjhkjfm2015b11);廣東海洋大學優秀學位論文培育項目(201605);國家自然科學基金項目(31201996)
廖健(1990-),男,碩士研究生,從事南海經濟魚類系統進化研究。E-mail:liaojiancc@foxmail.com
王中鐸(1980-),男,副教授。E-mail:aduof@gmail.com
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
