不同強度鹽脅迫下AM真菌對羊草生長的影響
2016-11-28 07:23:38張義飛張忠輝楊允菲
生態學報 2016年17期
張義飛,王 平,畢 琪,張忠輝,楊允菲
1 吉林省林業科學研究院,長春 130033 2 東北師范大學環境學院,國家環境保護濕地生態與植被恢復重點實驗室,長春 130117 3 東北師大附屬中學,長春 130022 4 東北師范大學生命科學學院,草地科學研究所,長春 130024
?
不同強度鹽脅迫下AM真菌對羊草生長的影響
張義飛1, *,王 平2,畢 琪3,張忠輝1,楊允菲4
1 吉林省林業科學研究院,長春 130033 2 東北師范大學環境學院,國家環境保護濕地生態與植被恢復重點實驗室,長春 130117 3 東北師大附屬中學,長春 130022 4 東北師范大學生命科學學院,草地科學研究所,長春 130024
不同濃度NaCl鹽處理下,AM真菌對羊草(Leymuschinensis)的侵染能力和對植物生長的影響,從植物形態和離子含量角度探討了AM真菌提高羊草耐鹽性的作用機理。結果表明,在高鹽脅迫下,AM真菌顯著降低了鹽脅迫效應,提高了羊草生物量,菌根效應明顯。菌根化羊草的根莖比顯著增加,并且N、P濃度較高,Na+和Cl-離子濃度較低,表明AM真菌即促進羊草對營養元素的吸收,又減少了離子毒害。菌根化羊草的Ca2+和K+離子濃度,以及P/Na+和K+/Na+比高于非菌根化羊草,表明AM真菌可通過調節滲透勢以避免或減緩鹽脅迫造成的生理缺水。隨著鹽脅迫的增加,菌根化羊草對磷的依賴性逐漸轉換為對鉀的依賴性。研究結果有助于揭示AM真菌提高植物耐鹽能力的作用機理,并對應用菌根技術修復鹽化草地具有理論指導意義。
鹽脅迫效應;菌根效應;侵染頻率;侵染強度;磷依賴性
菌根是植物根系和叢枝菌根真菌(Arbuscular Mycorrhizal Fungi,AMF)形成的共生體,是生物界最為廣泛的一種互惠共生現象[1]。借助于AM真菌,宿主植物可在惡劣環境中獲得良好的生長效應[2-3]。大量農作物和林木研究表明,AM真菌能夠提高植物的耐鹽性[4-5],增加作物在鹽漬條件下的產量[6],并且篩選出了促進植物在鹽漬土壤中生長的高效菌種[7]。隨著菌根技術的不斷發展,該研究和應用逐漸由農業和林業擴展到草業[8]。AM真菌-植物共生體在逆境環境中表現出來的互惠效應,為治理和恢復退化草地生態系統提供了新的研究途徑和方向[9]。
目前,全球范圍內共有約9.32億hm2土地遭受鹽化和堿化,這些地區的植被恢復與當地的食品安全、環境健康和經濟發展密切相關[10]。作為松嫩草甸草原的優勢物種和優良牧草,羊草(Leymuschinensis)與AM真菌共生體的特性和互惠效應在治理大面積鹽堿化草地的恢復工作中備受關注。已有研究表明,羊草能與多種AM真菌在自然條件下形成共生體[11-12],并且提高了羊草在重度鹽堿化草地上的定居率[9]。但當鹽脅迫壓力加大時,高鹽環境不僅抑制植物生長,同時還阻礙了AM真菌的孢子萌發和菌絲生長[13]。此外,雙方對共同資源的競爭可能會加劇降低AM真菌對羊草耐鹽性的促進作用,甚至轉變為單方或雙方受抑[14-15]。因此,了解不同強度鹽脅迫下AM真菌對羊草生長的影響,是應用菌根技術治理鹽化羊草草地的基礎。本研究設置了不同濃度的NaCl鹽處理,研究不同強度鹽脅迫對AM真菌的侵染能力和羊草菌根效應的影響,并從植物形態和離子含量角度探討菌根化羊草對鹽脅迫的響應機理。研究結果對于應用菌根技術治理和修復鹽堿退化羊草草地等實踐活動具有理論指導意義。
1 材料與方法
1.1 實驗材料
供試土壤取自溫室周圍的農田土。農田土的水溶性無機鹽含量為0.81 g/kg,土壤pH為8.49,有機質含量為2.49%,全氮、全磷、全鉀分別為1.69、0.91、34.8 g/kg,速效氮、速效磷、速效鉀分別為108.65、8.76、23.5 mg/kg。為改善土壤質地,將農田土過2 mm網篩后按1∶3的河沙∶農田土比例混合。將土壤河沙混合均勻后放入高壓蒸汽滅菌鍋中121 ℃滅菌2h,以消除土壤和河沙中的真菌孢子和其它土壤微生物。
羊草種子來自吉林綠源草業公司的羊草種子田。播種前將羊草種子用10 % H2O2浸泡消毒10min后,放入蒸餾水中浸泡2h,然后取出放置于濕潤濾紙上48h催芽備用。
AM真菌菌種取自吉林省西部鹽堿化羊草草地。經鑒定,該土壤中的主要菌種為球囊霉屬(Glomus)的摩西球囊霉(G.mosseae)和地球囊霉(G.geosporum)[16]。將上述兩個菌種在羊草根系上進行5個月的誘集培養,然后將包括菌根真菌菌絲、孢子、侵染根段及含根外菌絲體的根際土壤的混合物作為AM真菌根接種劑。并將一半接種劑進行滅菌處理,方法同土壤滅菌。
1.2 實驗設置
本研究于2005年6—8月在吉林省林業廳種苗基地溫室中進行。實驗期間室溫20—25 ℃,日光照射,光照時間控制為8:00—16:00的8h。本研究包含NaCl鹽和AM真菌接種兩個試驗因素,分別設置0、1、2、3 g NaCl / kg土的4個鹽處理水平,接種(+AM)和不接種(-AM)2個AM真菌處理水平,共計8個處理。每個處理4盆重復,共32盆。
塑料花盆的上口內徑為15 cm,盆底內徑為12 cm,盆高15 cm,盆底無孔。使用前將花盆用10 % H2O2浸泡消毒15min。每盆裝入2 kg已消毒的農田土-河沙混合物。按照設計的鹽處理水平,將相應重量的NaCl溶于蒸餾水中,并與花盆中的土壤充分混勻。在+AM處理的花盆中放入30 g菌根接種劑,將其放置在種子下2 cm深的土層中。在-AM處理的花盆中將30 g滅菌的接種劑放入到相同土層中,并放入30 mL不滅菌接種劑的水濾液(濾膜孔徑0.25 μm),以保持+AM和-AM處理除AM真菌外一致的微生物區系。
在花盆中均勻播種羊草種子,出苗后間苗,每盆留間隔均勻的50株。生長期間每周澆2次營養液,每次50 ml。營養液含N100 mg/L,K、P、Mg、Zn 各50 mg/L。實驗期間維持正常水分狀態。
1.3 樣品采集和測量
幼苗生長75 a后,齊地面收獲羊草地上部分,并挑出根系。用自來水沖洗地上部分和根系后,再用去離子水沖洗3—4遍,用濾紙吸干表面水分。記錄根系總鮮重。隨機選取一部分根系,記錄鮮重后,用于后續的菌根侵染測定。將羊草地上部分和剩余根系放入65 ℃下烘至恒重并稱重。
采用KOH透明-乳酸甘油酸性品紅染色法染色。將沖洗干凈的根系切成1cm左右長度的根段,將根段放入10 % KOH溶液中,水浴(90 ℃)60min,蒸餾水沖洗2次。隨后放入堿性雙氧水中軟化20min,水洗后在2 %的鹽酸中酸化3—4min。然后在90 ℃水浴鍋中用酸性品紅染色30min。取出根段后放入乳酸甘油(1∶1)中浸泡脫色。隨機選取50條根段壓片,在顯微鏡下觀察每條根的侵染長度(以mm記錄),并記錄被侵染的根系數量。
將羊草地上部分粉碎過40目篩。測量羊草地上部分的N、P、K+、Na+、Ca2+、Cl-元素的濃度。采用凱氏測氮儀(Kjeldahl, Foss Tractor)測N,紫外分光光度計(UV2201)測P(NaHCO3浸提-鉬銻抗比色法),原子吸收分光光度計(AA- 220 FS)測K+、Ca2+、Na+,離子色譜儀(DX- 300)測Cl-。
1.4 數據處理和統計分析
依據下列公式計算AM真菌對羊草根系的侵染情況[17-18]:
侵染頻率(CR)
CR%=(侵染根段數/總根段數)× 100
侵染強度(Ⅱ)
Ⅱ%=(侵染根長/總根長)× 100
以羊草的單位面積生物量為基礎,按下列公式計算每個鹽處理的鹽脅迫效應(SR)、菌根生長效應(MGR)、磷依賴性(MPR)和鉀依賴性(MKR)[19]:
依據羊草地上部分各離子的濃度含量和羊草地上生物量,計算羊草地上部分各離子的累積量。
使用混合模型(Mixed Models)分別檢驗NaCl和AM真菌因素對各響應變量的影響,以及兩者是否存在交互作用。以重復盆作為隨機因素納入模型中。對-AM和+AM處理,使用t檢驗進行均值比較。顯著水平均為P< 0.05。本實驗數據分析采用SPSS 19.0軟件。
2 結果
2.1 羊草生物量、根莖比和鹽脅迫效應
鹽和菌根處理對羊草總生物量影響顯著,但兩者的交互作用不明顯(表1)。隨NaCl濃度的增加,菌根化羊草生物量與非菌根化羊草的差異增加,至3 mg NaCl/kg水平時差異達到顯著水平(圖1)。鹽對羊草根莖比的影響不顯著,雖然菌根化羊草的根莖比隨NaCl濃度的增加而降低,但方差檢驗未達到顯著水平(F=2.704,P=0.092)(表1,圖1)。菌根化羊草的根莖比顯著高于非菌根羊草,但隨著鹽脅迫的增加,這種差異逐漸減小(圖1)。

表1 鹽和菌根處理影響羊草生長及鹽脅迫效應的方差分析表
隨著鹽濃度的增加,鹽脅迫效應隨之增加,在3 mg NaCl/kg水平下,鹽脅迫效應近40 %(表1,圖1)。AM真菌降低了羊草的鹽脅迫效應,并在高鹽條件下達到顯著水平(圖1)。
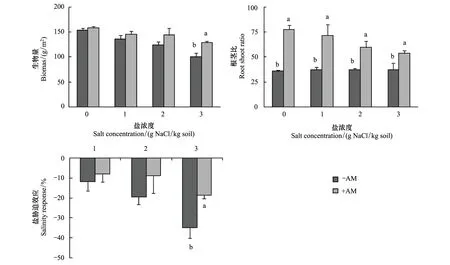
圖1 鹽對菌根化(+AM)和非菌根化(-AM)羊草的生物量和根莖比的影響及鹽脅迫效應Fig.1 The biomass and root/shoot ratio of mycorrhizal (+AM) and non-mycorrhizal (-AM) L. chinensis under different salt treatment and the salinity response不同字母表示不同菌根處理間存在顯著差異
2.2 羊草的侵染頻率、侵染強度和菌根生長效應
鹽脅迫顯著影響AM真菌對羊草根系的侵染頻率(F=11.12,P= 0.007)和侵染強度(F=6.72,P=0.014),尤其是侵染強度隨鹽濃度的增加而持續降低(圖2)。羊草菌根生長效應在無鹽條件下近于零,但隨NaCl濃度的增加而顯著增加(F=4.19,P= 0.041,圖2)。
2.3 羊草的離子濃度、離子累積量和離子比
鹽和菌根處理對羊草N、P濃度的影響達到極顯著水平,并且存在顯著的交互作用(表2)。菌根化羊草的N、P濃度始終高于非菌根化羊草,但這種差異隨著鹽脅迫的增加而不斷減少,至3 mg/kg高鹽時,-AM和+AM羊草的P濃度差異不顯著(圖3)。AM真菌促進羊草累積更多N、P(表2,圖4),但鹽脅迫對羊草N累積量的影響不顯著,卻顯著降低了P累積量(表2)。
方差分析顯示AM真菌對羊草K+濃度影響不顯著,但NaCl和AM真菌的交互作用表明,在不同鹽水平下,AM真菌對羊草K+濃度的影響不同(表2)。在無鹽條件下,菌根化羊草K+離子濃度顯著低于非菌根化羊草,但在2 mg/kg鹽條件下,菌根化羊草K+離子濃度卻顯著高于非菌根化羊草(圖3),并且在高鹽條件下K+累積量顯著高于非菌根化羊草(圖4)。
AM真菌對羊草Ca2+濃度的作用受鹽脅迫強度的影響(表2)。在0和1 mg/kg鹽水平下,菌根化羊草Ca2+濃度高于非菌根化羊草,但在2和3 mg/kg鹽水平下,顯著低于非菌根化羊草(圖3)。無論是菌根化羊草還是非菌根化羊草,Ca2+累積量隨鹽濃度的增加而降低。在0和3 mg NaCl/kg條件下,菌根化羊草吸收累積的Ca2+離子高于非菌根化羊草(圖4)。
無論是菌根化還是非菌根化,羊草Na+離子濃度隨鹽濃度的增加而增加。并且菌根化羊草Na+濃度一般低于非菌根化羊草(圖3)。在無鹽條件下,菌根化羊草鈉累積量低于非菌根化羊草,而在3 mg NaCl/kg高鹽條件下,菌根化羊草吸收累積的Na2+離子量高于非菌根羊化草(圖4)。
無論是Cl-濃度還是Cl-積累量,均隨著鹽濃度的增加而逐漸增加。除2 mg/kg處理外,非菌根化羊草的Cl-濃度顯著高于菌根化羊草(圖3)。與非菌根化羊草相比,菌根化羊草在無鹽條件下累積較少的氯離子,但在高鹽條件下吸收累積較多氯離子(圖4)。
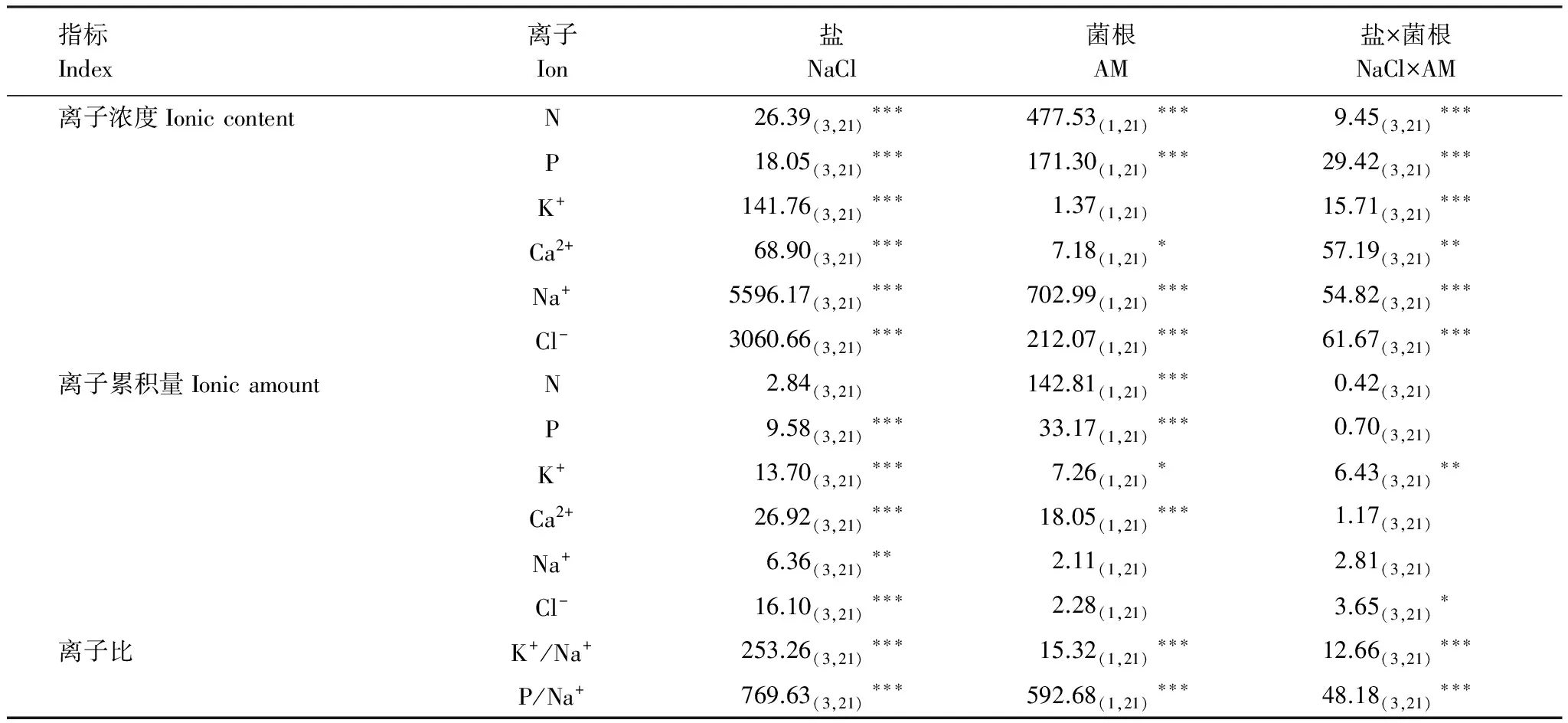
表2 鹽和菌根處理影響羊草離子濃度和累積量的方差分析表
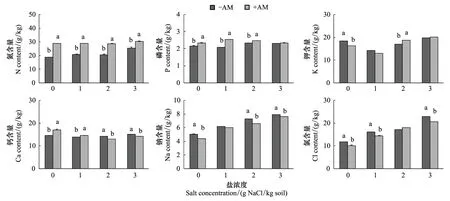
圖3 不同鹽條件下菌根化(+AM)和非菌根化(-AM)羊草的離子濃度含量Fig.3 The ionic contents of mycorrhizal (+AM) and non-mycorrhizal (-AM) L. chinensis under different salt treatments不同字母表示不同菌根處理間存在顯著性差異
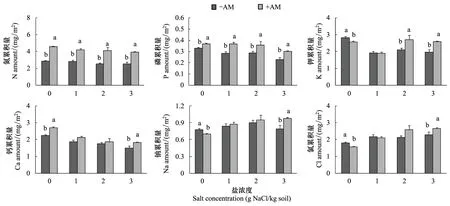
圖4 不同鹽條件下菌根化(+AM)和非菌根化(-AM)羊草的離子累積量Fig.4 The ionic amounts of mycorrhizal (+AM) and non-mycorrhizal (-AM) L. chinensis under different salt treatments不同字母表示不同菌根處理間存在顯著性差異
無論是K+/Na+還是P/Na+,均在有鹽條件下低于無鹽條件下(表2,圖5)。AM真菌對這兩個指標的影響在不同鹽水平下表現不一。在無鹽和低鹽條件下,有無菌根對羊草的K+/Na+沒有顯著影響,但在2、3 mg/kg高鹽水平下,AM真菌顯著提高了羊草的K+/Na+(圖5)。P/Na+與此相反,在無鹽和低鹽條件下,AM真菌顯著提高了羊草的P/Na+,但在2、3 mg /kg高鹽水平下有無菌根對該指標無顯著影響(圖5)。
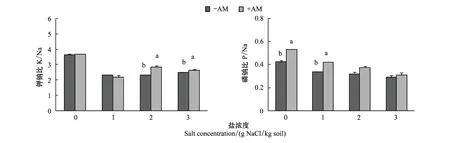
圖5 不同鹽條件下菌根化和非菌根化羊草的鉀鈉比和磷鈉比Fig.5 The K+/Na+ and P/Na+ ratios of mycorrhizal and non-mycorrhizal L. chinensis under different salt stress
2.4 菌根化羊草的磷、鉀依賴性
磷依賴性指標在所有鹽處理中表現為正值,表明無論土壤中有無NaCl鹽,菌根化羊草對磷的依賴性始終存在。但在低鹽條件下(1 mg/kg)菌根化羊草對磷的依賴性最強,隨著鹽脅迫的不斷增加,磷依賴性隨之下降(圖6)。菌根化羊草僅在2、3 mg/kg高鹽水平條件下表現出鉀依賴性(圖6)。
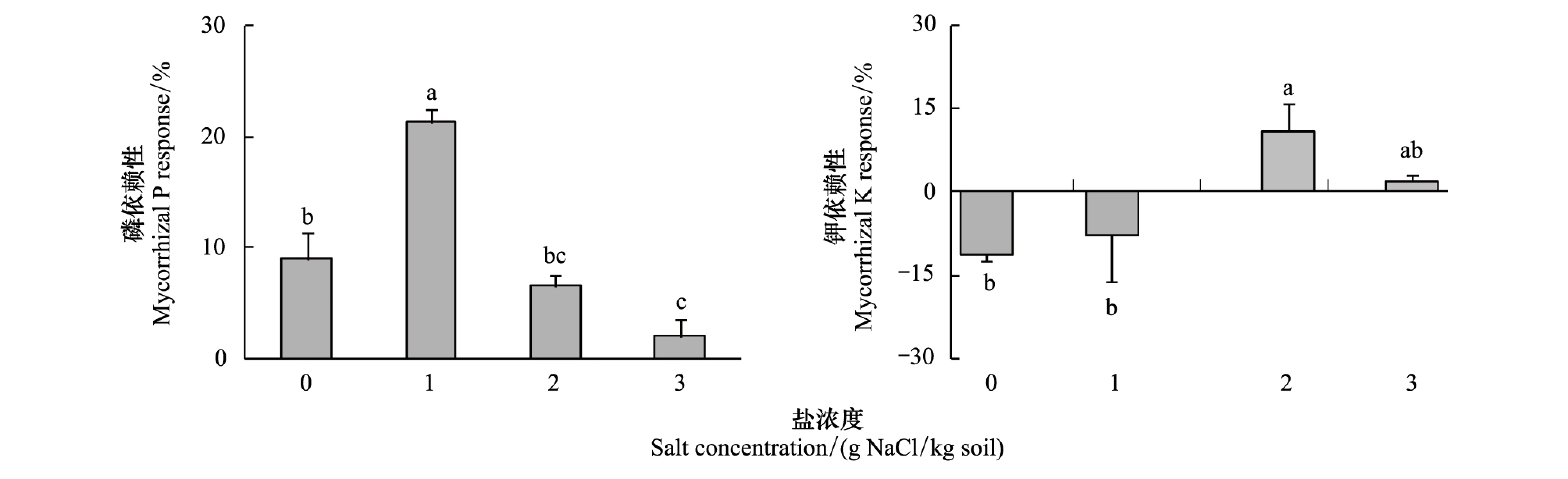
圖6 不同鹽脅迫條件下菌根化羊草的磷依賴性和鉀依賴性Fig.6 The mycorrhizal P response and mycorrhizal K response of L. chinensis under different salt stress
3 討論
3.1 AM真菌降低了羊草的鹽脅迫效應,但侵染活動隨鹽脅迫的增加而下降
一般而言,叢枝菌根真菌侵染可改善植物的生長狀況,但改善程度取決于土壤性質[2, 20]。當生長環境良好,植物未受到脅迫或脅迫較低時,AM真菌與植物的互利現象并不明顯。如本研究中的無鹽和低鹽處理,AM真菌對羊草生物量的促進作用并未達到顯著水平,即羊草的菌根生長效應并不明顯。當然,該研究結果也可能由于數據來源自羊草生物量積累較為緩慢的幼苗階段,AM真菌的菌根生長效應并為完全發揮出來。但即使是在幼苗階段,當羊草植株面臨較高的鹽脅迫時,AM真菌顯著降低了NaCl對羊草產生的鹽脅迫效應,這充分證明了AM真菌-植物共生體對植物在逆境中生存的重要性。
但當逆境中的脅迫強度加大時,不僅植物生長受到抑制,同時還阻礙了AM真菌的孢子萌發和菌絲生長[13]。雖然本研究使用的摩西球囊霉(G.mosseae)和地球囊霉(G.geosporum)是重度鹽土中最優勢的AM真菌物種[4],但在高鈉鹽脅迫下,其孢子萌發和菌絲生長也會受到抑制[21]。這不僅降低了AM真菌對羊草根系的侵染能力,并且有可能導致AM真菌和羊草為爭奪有限資源而形成相互抑制的競爭關系。雖然羊草的菌根生長效應并沒有隨鹽分濃度的增加而下降,但AM真菌對羊草根系侵染頻率和侵染強度卻隨鹽分的增加而顯著下降。因此,隨著脅迫繼續加劇, AM真菌對羊草耐鹽性的促進作用將會降低,甚至轉變為單方或雙方受抑[14-15]。事實上,在環境惡劣的生態系統中,AM真菌和植物共生體面臨著多種影響其互利關系的生物和非生物因素,其綜合作用可能會加速削弱AM真菌對植物生長的促進作用。
3.2 菌根化羊草的根莖比增加
一些研究證明AM真菌能夠增強植物的存活率和生物量[22-23]。在無鹽或鹽脅迫較低時,AM真菌并沒有顯著改變羊草的生物量,卻極大改變了羊草生物量的地上地下分配比例,并且在所有鹽處理中,菌根化植物一直保持較高的根莖比。有研究表明增加的根生長能夠促進水分和營養的吸收[24- 26]。菌根化羊草將更多的光合產物分配到根系,為植株的后期生長發育打下了堅實的基礎和保障。相對增加的根系將比無菌根化羊草植株具有更強的水分和營養元素吸收和傳輸能力,從而能為菌根化植株拓展更為廣闊的資源空間和生存機會。因此,菌根化植株重新分配根莖比可能是AM真菌促進植物生長的一條重要作用途徑。
3.3 菌根化羊草的離子含量和離子比發生變化
除形態變化外,菌根化植物的營養元素含量也發生變化,從而增強了其生長能力[27]。在鹽脅迫條件下,因土壤中鹽離子過多而排斥植物對其他離子的吸收,導致植物營養失調,抑制生長。一些研究表明,接種AM真菌能夠提高植株的營養元素含量[27-28]。本實驗中的菌根化羊草一直維持較高的N、P濃度,但隨著鹽脅迫的加大,該優勢逐漸減少。一方面可能由于鹽分增加抑制了AM真菌對羊草根系的侵染能力,從而降低其營養吸收能力;另一方面可能由于AM真菌在惡劣環境里與寄主植物之間的營養競爭較激烈,從而減少了寄主一方的利益。雖然高鹽條件下羊草P濃度無差異,但在整個實驗過程中,菌根化羊草比非菌根化羊草吸收積累了更多的P元素,證明了AM真菌能夠促進植物吸收利用更多的N、P元素。
AM真菌有效減輕了鹽脅迫條件下的離子毒害作用[29]。菌根化羊草一直維持著較低的Na+和Cl-離子濃度。有關AMF減緩鹽離子對植物毒害作用的機理還未有一致的結論。可能是由于菌根化植株的P、K+和Ca2+等離子含量增加,不僅降低了細胞滲透勢,還有效降低了鹽離子的相對含量,從而減輕鹽離子的毒害作用[30]。也可能由于AM真菌促進植株生長,由此相對稀釋了有毒離子的濃度,減緩了離子毒害[13]。
鹽脅迫對植物的危害還表現在滲透勢增加所導致的生理缺水[31]。菌根化植物除了大量吸收外界營養元素促進生長外,還大量吸收一些其它離子來調節滲透勢,諸如K+,從而提高植物的耐鹽能力[32]。低鹽條件下菌根化羊草的Ca2+離子濃度和中高鹽條件下的K+離子濃度均高于非菌根化羊草。Ca2+、K+離子濃度的增加有效降低了細胞滲透勢,其中K+在高鹽脅迫環境下對改善細胞滲透勢更為重要。此外,低鹽條件下菌根化羊草的P/Na+比,和中高鹽條件下的K+/Na+比均高于非菌根化羊草。因此,維持較高的P/Na+、K+/Na+可能也是AM真菌增強羊草耐鹽性的機制之一[33]。隨著鹽脅迫的增加,菌根化羊草對磷的依賴性逐漸轉換為對鉀的依賴性,羊草的抗鹽能力隨著鹽脅迫的增加越來越依賴于鉀元素。
[1] Smith S E,Facelli E,Pope S,Smith A F. Plant performance in stressful environments:interpreting new and established knowledge of the roles of arbuscular mycorrhizas. Plant and Soil,2010,326(1/2):3- 20.
[2] Evelin H,Kapoor R,Giri B. Arbuscular mycorrhizal fungi in alleviation of salt stress:a review. Annals of Botany,2009,104(7):1263- 1280.
[3] 李曉林,馮固. 叢枝菌根生態生理. 北京:華文出版社,2001.
[4] Bothe H. Arbuscular mycorrhiza and salt tolerance of plants. Symbiosis,2012,58(1/3):7- 16.
[5] Talaat N B,Shawky B T. Protective effects of arbuscular mycorrhizal fungi on wheat (TriticumaestivumL.) plants exposed to salinity. Environmental and Experimental Botany,2014,98:20- 31.
[6] Talaat N B,Shawky B T. Influence of arbuscular mycorrhizae on root colonization,growth and productivity of two wheat cultivars under salt stress. Archives of Agronomy and Soil Science,2012,58(1):85- 100.
[7] Estrada B,Aroca R,Barea J M,Ruiz-Lozano J M. Native arbuscular mycorrhizal fungi isolated from a saline habitat improved maize antioxidant systems and plant tolerance to salinity. Plant Science,2013,201- 202:42- 51.
[8] Karanika E D,Mamolos A P,Alifragis D A,Kalburtji K L,Veresoglou D S. Arbuscular mycorrhizas contribution to nutrition,productivity,structure and diversity of plant community in mountainous herbaceous grassland of northern Greece. Plant Ecology,2008,199(2):225- 234.
[9] Zhang Y F,Wang P,Yang Y F,Bi Q,Tian S Y,Shi X W. Arbuscular mycorrhizal fungi improve reestablishment ofLeymuschinensisin bare saline-alkaline soil:Implication on vegetation restoration of extremely degraded land. Journal of Arid Environments,2011,75(9):773- 778.
[10] Rengasamy P. World salinization with emphasis on Australia. Journal of Experimental Botany,2006,57(5):1017- 1023.
[11] Carvalho L M,Ca?ador I,Martins-Lou??o M. Temporal and spatial variation of arbuscular mycorrhizas in salt marsh plants of theTagusestuary(Portugal). Mycorrhiza,2001,11(6):303- 309.
[12] 張偉溪,王洪峰,趙昕,閻秀峰. 松嫩鹽堿草地植物叢枝菌根的初步調查. 土壤通報,2010,41(6):1380- 1385.
[13] Juniper S,Abbott L. Vesicular-arbuscular mycorrhizas and soil salinity. Mycorrhiza,1993,4(2):45- 57.
[14] Johnson N C,Graham J H,Smith F A. Functioning of mycorrhizal associations along the mutualism-parasitism continuum. New Phytologist,1997,135(4):575- 586.
[15] Karst J,Marczak L,Jones M D,Turkington R. The mutualism-parasitism continuum in ectomycorrhizas:A quantitative assessment using meta-analysis. Ecology,2008,89(4):1032- 1042.
[16] 張義飛. 松嫩平原鹽堿化草地叢枝菌根真菌資源及其生態作用的研究[D]. 長春:東北師范大學,2011.
[17] McGonigle T P,Miller M H,Evans D G,Fairchild G L,Swan J A. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytologist,1990,115(3):495- 501.
[18] Biermann B,Linderman R G. Quantifying vesicular-arbuscular mycorrhizae:a proposed method towards standardization. New Phytologist,1981,87(1):63- 67.
[19] Colla G,Rouphael Y,Cardarelli M,Tullio M,Rivera C M,Rea E. Alleviation of salt stress by arbuscular mycorrhizal in zucchini plants grown at low and high phosphorus concentration. Biology and Fertility of Soils,2008,44(3):501- 509.
[20] 江龍,王茂勝,黃建國,袁玲. 叢枝菌根真菌對煙苗吸收基質養分的影響. 貴州農業科學,2010,38(6):50- 54.
[21] Carvalho L M,Correia P M,Ca?ador I,Martins-Lou??o M A. Effects of salinity and flooding on the infectivity of salt marsh arbuscular mycorrhizal fungi inAstertripoliumL.. Biology and Fertility of Soils,2003,38(3):137- 143.
[22] Kaya C,Ashraf M,Sonmez O,Aydemir S,Tuna A L,Cullu M A. The influence of arbuscular mycorrhizal colonisation on key growth parameters and fruit yield of pepper plants grown at high salinity. Scientia Horticulturae,2009,121(1):1- 6.
[23] Sheng M,Tang M,Chen H,Yang B W,Zhang F F,Huang Y H. Influence of arbuscular mycorrhizae on the root system of maize plants under salt stress. Canadian Journal of Microbiology,2009,55(7):879- 886.
[24] Müller I,Schmid B,Weiner J. The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plants. Perspectives in Plant Ecology,Evolution and Systematics,2000,3(2):115- 127.
[25] Eissenstat D M,Graham J H,Syvertsen J P,Drouillard D L. Carbon economy of sour orange in relation to mycorrhizal colonization and phosphorus status. Annals of Botany,1993,71(1):1- 10.
[26] Johnson D,Leake J R,Ostle N,Ineson P,Read D J.Insitu13CO2pulse-labelling of upland grassland demonstrates a rapid pathway of carbon flux from arbuscular mycorrhizal mycelia to the soil. New Phytologist,2002,153(2):327- 334.
[27] Rabie G H. Influence of arbuscular mycorrhizal fungi and kinetin on the response of mungbean plants to irrigation with seawater. Mycorrhiza,2005,15(3):225- 230.
[28] Giri B,Kapoor R,Mukerji K G. Improved tolerance ofAcacianiloticato salt stress by arbuscular mycorrhiza,Glomusfasciculatummay be partly related to elevated K/Na ratios in root and shoot tissues. Microbial Ecology,2007,54(4):753- 760.
[29] Garg N,Chandel S. Role of arbuscular mycorrhizal (AM) fungi on growth,cadmium uptake,osmolyte,and phytochelatin synthesis inCajanuscajan(L.) millsp. under NaCl and Cd stresses. Journal of Plant Growth Regulation,2012,31(3):292- 308.
[30] 馮固,張福鎖. 叢枝菌根真菌對棉花耐鹽性的影響研究. 中國生態農業學報,2003,11(2):21- 24.
[31] 馮固,李曉林,張福鎖,李生秀. 鹽脅迫下叢枝菌根真菌對玉米水分和養分狀況的影響. 應用生態學報,2000,11(4):595- 598.
[32] 賀忠群,賀超興,張志斌,鄒志榮. 叢枝菌根真菌提高植物耐鹽性的作用機制. 西北植物學報,2007,27(2):414- 420.
[33] 馮固,楊茂秋,白燈莎. 鹽脅迫下VA菌根真菌對無芒雀麥體內礦質元素含量及組成的影響. 草業學報,1998,7(3):21- 28.
The effect of the arbuscular mycorrhizal fungi on the growth of Leymus chinensis under saline stress of different intensities
ZHANG Yifei1,*, WANG Ping2, BI Qi3, ZHANG Zhonghui1, YANG Yunfei4
1 Jilin Academy of Forestry Science, Changchun 130033, China 2StateEnvironmentProtectionKeyLaboratoryofWetlandEcologyandVegetationRestoration,SchoolofEnvironment,NortheasternNormalUniversity,Changchun130117,China3HighSchoolAttachedtoNortheastNormalUniversity,Changchun130022,China4InstituteofGrasslandScience,SchoolofLifeScience,NortheastNormalUniversity,Changchun130024,China
Arbuscular mycorrhizal fungi (AMF) could improve the tolerance of plants under saline stress. However, the degree of this improvement might differ relative to salt stress intensity. We conducted an experiment to explore the ability of AMF to colonizeLeymuschinensisroots and evaluate the effect of AMF inoculation on the growth of plants under four NaCl concentrations (0, 1, 2, and 3 g NaCl/kg soil). The mechanism behind the saline tolerance of mycorrhizalL.chinensisplants was also discussed in terms of their root:shoot ratio and ionic content. The inoculants used in this study wereGlomusmosseaeandG.geosporum, two dominant species found in highly saline-alkaline natural grasslands. Sixteen pots of inoculatedL.chinensisseedlings were randomly subjected to one of four NaCl treatments, as were sixteen pots of non-inoculated plants of the same species. The plants were harvested and measured after 75 d of growth. The results show that under high saline stress, the AMF significantly decreased the strength of the salinity response and increased plant biomass. The mycorrhizal growth response under high saline stress was significantly positive, which indicated that the AMF-plant symbiotic relationship is important to plants under salt stress. However, the colonization rate and infection intensity of AMF inL.chinensisroots both decreased as salt stress increased. The ability of AMF to improve plant growth might decrease, and one-sided or mutually harmful negative effects might occur when salt stress becomes worse, because of the increasing intensity of the competition between AMF and plants. The AMF significantly increased the root:shoot ratio in each NaCl treatment. More photosynthetic product was allocated to the roots of mycorrhizal plants than those of non-mycorrhizal plants, which increased both the water and nutrient absorption, and transportation capacity of the AMF-plant symbiont. The re-allocation of resources between roots and shoots might be one of the primary strategies used by mycorrhizal plants to increase their fitness under harsh environmental conditions. The higher N and P concentrations observed in mycorrhizal plants also support the idea that AMF helpL.chinensisabsorb and utilize more nutrients under saline stress. The concentrations of Na+and Cl-were both lower in mycorrhizal plants than in non-mycorrhizal plants, decreasing the ionic content of inoculated plants. Ca2+and P/Na+concentrations were higher in mycorrhizal plants under low saline stress, while the K+and K+/Na+concentrations were higher in mycorrhizal plants under high saline stress, indicating that the mycorrhiza could reduce the physiological drought response of plants under saline stress by adjusting their osmotic potential via absorbing other ions and/or increasing ionic ratios such as K+/Na+. The results also showed that the saline tolerance of mycorrhizal plants was increased by the mycorrhizal P response under lower saline stress, and by the mycorrhizal K response under high saline stress. This study has improved the understanding of the mechanism underlying the improved saline tolerance of mycorrhizal plants, and provides some guidelines for using AMF technology to restore saline-degraded grasslands.
salinity response; mycorrhizal growth response; colonization rate; infection intensity; mycorrhizal phosphorus response
國家自然科學基金項目(31170504);吉林省林業廳育林基金林業科技項目(2014-001)
2015- 02- 18;
日期:2015- 12- 14
10.5846/stxb201502180372
*通訊作者Corresponding author.E-mail: yifeii@hotmail.com
張義飛,王平,畢琪,張忠輝,楊允菲.不同強度鹽脅迫下AM真菌對羊草生長的影響.生態學報,2016,36(17):5467- 5476.
Zhang Y F, Wang P, Bi Q, Zhang Z H, Yang Y F.The effect of the arbuscular mycorrhizal fungi on the growth ofLeymuschinensisunder saline stress of different intensities.Acta Ecologica Sinica,2016,36(17):5467- 5476.
猜你喜歡
核科學與工程(2021年4期)2022-01-12 06:30:26
小讀者(2021年2期)2021-03-29 05:03:48
今日農業(2020年19期)2020-12-14 14:16:52
華人時刊(2019年13期)2019-11-17 14:59:54
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
中學物理·高中(2016年12期)2017-04-22 11:53:03
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
