清絡(luò)通痹方調(diào)控miRNA網(wǎng)絡(luò)干預(yù)類風(fēng)濕關(guān)節(jié)炎的研究①
2016-12-13 11:28:51朱亞梅周玲玲彭孝武周學(xué)平
中國免疫學(xué)雜志 2016年4期
關(guān)鍵詞:小鼠
朱亞梅 周玲玲 彭孝武 耿 姍 周學(xué)平
(南京中醫(yī)藥大學(xué),南京210023)
?
·中醫(yī)中藥與免疫·
清絡(luò)通痹方調(diào)控miRNA網(wǎng)絡(luò)干預(yù)類風(fēng)濕關(guān)節(jié)炎的研究①
朱亞梅 周玲玲 彭孝武 耿 姍 周學(xué)平②
(南京中醫(yī)藥大學(xué),南京210023)
目的:觀察清絡(luò)通痹方(QLT)對膠原性關(guān)節(jié)炎(CIA)小鼠miRNA網(wǎng)絡(luò)的調(diào)控作用,探討其干預(yù)類風(fēng)濕關(guān)節(jié)炎的機(jī)制。方法:DBA/1小鼠,復(fù)制CIA模型,隨機(jī)分為空白對照、CIA、QLT組,采用miRCURYTM LNA Array芯片技術(shù)篩選外周血miRNA異常表達(dá)譜,以Realtime PCR對異常變化的miRNAs進(jìn)行驗證,并利用多種生物信息學(xué)軟件進(jìn)行分析。結(jié)果:與空白對照組比較,CIA小鼠具有差異表達(dá)的miRNAs有221個;與CIA比較,QLT差異表達(dá)的miRNAs有169個。RT-PCR驗證結(jié)果與芯片檢測所示具有較好的一致性。PCR結(jié)果顯示CIA中miR-143下調(diào)明顯,而QLT干預(yù)可明顯上調(diào)miR-143的表達(dá)水平。miR-143靶基因顯著富集在VEGF、T細(xì)胞受體、MAPK信號通路等信號通路中。結(jié)論:多種miRNA的異常表達(dá)參與CIA的病理過程,QLT可能通過干預(yù)miRNA調(diào)控網(wǎng)絡(luò),多環(huán)節(jié)、多途徑干預(yù)RA免疫、炎癥、疼痛等多種病理過程,miR-143可能作為QLT治療RA的重要環(huán)節(jié)。
miRNA;膠原性關(guān)節(jié)炎;清絡(luò)通痹方
類風(fēng)濕關(guān)節(jié)炎(Rheumatoid arthritis,RA)是一種臨床常見的風(fēng)濕免疫性疾病,主要表現(xiàn)為自身免疫的紊亂,進(jìn)而引起滑膜炎和骨質(zhì)破壞。微小 RNAs(microRNAs,miRNAs)作為一類內(nèi)源性、短鏈非編碼RNA主要功能是調(diào)節(jié)蛋白編碼基因的表達(dá),越來越多的研究揭示miRNA調(diào)控網(wǎng)絡(luò)在自身免疫性疾病的發(fā)病機(jī)理中扮演著十分重要的角色[1],并且大量研究發(fā)現(xiàn)在RA關(guān)節(jié)滑膜組織以及滑液成纖維細(xì)胞中miR-155、miR-146a、miR-203等多種miRNA 的表達(dá)失調(diào)。清絡(luò)通痹方(Qingluo Tongbi Compound,QLT)由生地黃、青風(fēng)藤、三七、炙僵蠶、雷公藤組成,為國醫(yī)大師周仲瑛教授臨床應(yīng)用幾十年的經(jīng)驗方,針對RA臨床常見的“陰虛絡(luò)熱證”,具有養(yǎng)陰清熱、祛風(fēng)除濕、活血祛瘀、化痰散結(jié)的作用。前期研究提示QLT可通過抑制細(xì)胞因子網(wǎng)絡(luò)紊亂、調(diào)控多條信號通路抑制RA免疫功能的紊亂和滑膜炎癥[2,3],從多環(huán)節(jié)、多途徑干預(yù)RA。單個miRNA可調(diào)控生物體內(nèi)多個基因的表達(dá),而QLT亦具有類似的多靶點調(diào)控的特點,因此我們推測QLT能通過調(diào)節(jié)miRNA的表達(dá)干預(yù)RA病理過程。本研究將應(yīng)用高通量miRNA表達(dá)譜芯片分析膠原性關(guān)節(jié)炎(Collagen-induced arthritis,CIA)模型組和QLT給藥組miRNA的差異表達(dá),并通過數(shù)據(jù)整合和應(yīng)用系統(tǒng)生物學(xué)分析方法進(jìn)一步挖掘數(shù)據(jù)中的生物學(xué)功能信息,為進(jìn)一步認(rèn)識QLT干預(yù)RA機(jī)制提供新的線索。
1 材料與方法
1.1 材料
1.1.1 藥品與試劑 清絡(luò)通痹方水提液由江蘇省植物藥研究與新藥開發(fā)中心提供,清絡(luò)通痹方水提液含生藥1.29 g/ml,使用前蒸餾水稀釋。
不完全弗氏佐劑(Incomplete Freund′s adjuvant,IFA,美國Sigma公司,批號12401);完全弗氏佐劑(Complete Freund′s adjuvant,CFA,美國sigma公司,批號12441);牛Ⅱ型膠原(type II collagen,CⅡ,美國Chondrex公司,批號130217);Trizol試劑(美國Life Technologies公司,批號79403);miRCURYTM Array Power Labeling kit(丹麥Exiqon公司,批號208032-A); miRCURYTM Array,Wash buffer kit(丹麥Exiqon公司,批號208021)。
1.1.2 儀器 熒光定量PCR(羅氏公司 型號:Lishtcycler96);紫外分光光度計(USA Thermo公司 型號:ND-1000);Axon GenePix 4000B 掃描儀(Axon Instruments,Foster City,CA);GenePix Pro 6.0 軟件(Axon,CA)。
1.1.3 動物 DBA/1小鼠,雄性,6~8周,9只,常州卡文斯動物公司提供[SCXK(蘇)2011-0003],試驗在南京中醫(yī)藥大學(xué)實驗動物中心內(nèi)完成,室溫(23±3)℃,濕度40%~70 %。動物給予全價顆粒飼料(江蘇協(xié)同醫(yī)藥生物有限公司供給)喂養(yǎng),自由飲水,每日光照/黑暗各12 h。動物適應(yīng)性飼養(yǎng)3 d后進(jìn)行試驗。
1.2 方法
1.2.1 CIA小鼠的制備[4]將牛CⅡ溶于0.01 mol/L的醋酸中,配制成2 g/L溶液,4℃震蕩過夜,將溶解的CⅡ與CFA以1∶1比例混,并充分乳化(在冰上操作)。在DBA/1 小鼠尾根靠后處皮內(nèi)注射100 μl CⅡ與CFA 的混合乳劑進(jìn)行初次免疫,當(dāng)天記為d1。到d21用CⅡ與IFA等比混勻,尾根部注射100 μl進(jìn)行加強(qiáng)免疫。
1.2.2 分組及給藥 隨機(jī)分為3組,每組3只:空白對照組(Normal)不予任何處理,其余組復(fù)制CIA模型,分為CIA組,清絡(luò)通痹方組(QLT)8.7 g/kg,空白對照組和CIA組給予等體積生理鹽水,QLT 組d21起每日灌胃給藥1次,給藥4周后,眼眶取血,-80℃冰箱保存。
1.2.3 總RNA抽提 將處理好的樣本中加入Trizol混合按Trizol 說明抽提總RNA,并測定RNA濃度和純度。
1.2.4 MiRNA芯片分析 采用miRCURYTM Array Power Labeling kit標(biāo)記,然后應(yīng)用miRCURYTMLNA Array(v.18.0)進(jìn)行芯片雜交。雜交后芯片用Wash buffer kit進(jìn)行洗片,具體步驟按各試劑盒說明書進(jìn)行。采用Axon GenePix 4000B掃描儀對雜交后芯片進(jìn)行掃描,使用GenePix Pro6.0軟件對掃描圖像所得原始數(shù)據(jù)分析,通過原始值減去背景值來做修正,并用中值做標(biāo)準(zhǔn)化芯片上每個探針的綠色信號強(qiáng)度經(jīng)過去背景化后,并將4個重復(fù)探針取平均值,用中位數(shù)標(biāo)準(zhǔn)化的方法對數(shù)據(jù)進(jìn)行標(biāo)準(zhǔn)化處理,組間信號值進(jìn)行比較,比值>1.5為表達(dá)上調(diào),<0.5為表達(dá)下調(diào)。
1.2.5 real-time PCR 采用Primer 5.0設(shè)計引物,引物序列見表1,按照試劑盒指定操作步驟擴(kuò)增,進(jìn)行real-time PCR反應(yīng),PCR程序為:95℃,10 min;40個PCR循環(huán)[95℃,10 s;60℃,60 s(收集熒光)]。擴(kuò)增反應(yīng)結(jié)束后,按 (95℃,10 s;60℃,60 s;95℃,15 s);并從60℃緩慢加熱到99℃(儀器自動進(jìn)行-Ramp Rate為0.05℃/s)建立PCR產(chǎn)物的熔解曲線。獲得擴(kuò)增曲線及CT值。各樣品的目的miRNA和內(nèi)參(mmu-miR-425-5p)分別進(jìn)行realtime PCR反應(yīng)。采用mmu-miR-425-5p作為實驗的內(nèi)參基因進(jìn)行歸一化,數(shù)據(jù)采用2-△△ CT法進(jìn)行分析,設(shè)置 3 個平行復(fù)孔,檢測結(jié)果取平均值。
表1 實時定量RT-PCR引物
Tab.1 List of primers used in Real time PCR

miRNAPrimersequencemmu-miR-425-5pGSP:5'GGGGAATGACACGATCACTC3'R:5'GTGCGTGTCGTGGAGTCG3'mmu-miR-183-5pGSP:5'GGGTATGGCACTGGTAGAA3'R:5'TGCGTGTCGTGGAGTC3'mmu-miR-203-3pGSP:5'GGGGTGAAATGTTTAGGA3'R:5'TGCGTGTCGTGGAGTC3'mmu-miR-122-5pGSP:5'GGGTGGAGTGTGACAATGG3'R:5'GTGCGTGTCGTGGAGTCG3'mmu-miR-143-3pGSP:5'GGGATGAGATGAAGCACT3'R:5'TGCGTGTCGTGGAGTC3'
Note:GSP.Corresponding to the specific primers of micrornas;R.The primers match the RT primers.
1.2.6 生物信息學(xué)分析 應(yīng)用miRanda、Target-Scan、miRRDB、miRWalk、RNA22靶基因預(yù)測網(wǎng)站預(yù)測靶基因,取三者交集的靶基因以最大程度的降低預(yù)測假陽性率,利用Cytoscape中的插件BINGO對 mmu-miR-143 預(yù)測靶基因集合進(jìn)行功能注釋(Gene Ontology ,GO)富集分析,并采用 DAVID 數(shù)據(jù)庫,對預(yù)測靶基因集合進(jìn)行基于KEGG數(shù)據(jù)庫的生物通路富集分析。
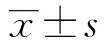
2 結(jié)果
2.1RNA提取結(jié)果 各RNA樣本的A260/A280比值均在1.6~1.9之間,提示各樣本中總RNA純度良好,符合miRNA芯片的質(zhì)量要求,保證了下一步實驗的順利進(jìn)行。
2.2miRNA芯片篩選結(jié)果miRNA芯片提示:與正常對照組小鼠比較,CIA組小鼠具有差異表達(dá)的miRNA有221個,其中115個miRNAs表達(dá)上調(diào)(↑),106個miRNAs表達(dá)下調(diào)(↓);與CIA組比較,QLT組差異表達(dá)的miRNA有169個,93個miRNAs表達(dá)上調(diào)(↑),76個miRNAs表達(dá)下調(diào)(↓)。CIA/Normal與QLT/CIA具有反向作用的miRNA共49個,QLT干預(yù)可上調(diào)其表達(dá)水平的miRNA有36個,其中20個miRNAs變化明顯(CIA/Normal下調(diào)變化倍數(shù)<0.5,QLT/CIA上調(diào)變化倍數(shù)>1.5倍),見表2;QLT干預(yù)可下調(diào)其表達(dá)水平的miRNA有13個,其中6個miRNAs變化明顯(CIA/Normal上調(diào)>1.5倍,QLT/CIA下調(diào)變化倍數(shù)<0.5),見表3。
2.3real-timePCR芯片結(jié)果驗證 為檢測芯片結(jié)果的真實可靠性從芯片結(jié)果中選取4個差異表達(dá)的miRNA進(jìn)行real-timePCR,對芯片結(jié)果進(jìn)行初步驗證,結(jié)果顯示CIA組與空白對照組相比,mmu-miR-183-5p、mmu-miR-203-3p、mmu-miR-143-3p、mmu-miR-122-5p變化倍數(shù)CIA/Normal<0.5倍,表達(dá)均上調(diào)(圖1A),QLT/CIA變化倍數(shù)>1.5倍,清絡(luò)通痹方干預(yù)均可下調(diào)其表達(dá)(圖1B),結(jié)果顯示,所選miRNA在芯片結(jié)果中的表達(dá)均與RT-PCR變化倍數(shù)表達(dá)情況一致,如圖1,說明本研究的芯片結(jié)果基本真實可靠。
PCR結(jié)果顯示:空白對照組miR-143相對表達(dá)量為1.52±0.44,CIA組中miR-143相對表達(dá)量為0.59±0.23,與空白對照組相比CIA組miR-143下調(diào)明顯,差異具有統(tǒng)計學(xué)意義(P<0.05);QLT組其相對表達(dá)量為1.08±0.08,與CIA組相比miR-143表達(dá)水平上調(diào),差異顯著(P<0.05),如圖2。
表2 清絡(luò)通痹方具有上調(diào)干預(yù)作用miRNA的變化倍數(shù)
Tab.2 Fold changes of miRNAs up-regulated by QLT

miRNAFoldchange(CIA/Normal)Foldchange(QLT/CIA)mmu-miR-29a-5p0.469507014↓1.699634138↑mmu-miR-181a-5p0.402391583↓2.093295151↑mmu-miR-27a-3p0.450875355↓1.588985215↑mmu-miR-183-5p0.428255255↓1.900674813↑mmu-miR-20b-5p0.495110635↓2.096817098↑mmu-miR-33-5p0.420156433↓2.024470612↑mmu-miR-194-5p0.306229664↓2.327504931↑mmu-miR-203-3p0.305148223↓1.801923623↑mmu-miR-199a/199b-3p0.344007709↓2.240078583↑mmu-miR-335-5p0.199094743↓2.907107386↑mmu-miR-19a-3p0.415888524↓1.914543554↑mmu-miR-32-5p0.31344649↓2.239427598↑mmu-miR-101a/c-3p0.413383695↓3.109887676↑mmu-miR-195a-5p0.419610889↓1.978721831↑mmu-miR-122-5p0.194012485↓2.238766954↑mmu-miR-344i0.228540999↓2.055180739↑mmu-miR-215-5p0.380389709↓2.484540029↑mmu-miR-544-5p0.446442457↓1.731614896↑mmu-miR-301a-3p0.437327066↓1.919305201↑mmu-miR-143-3p0.451400018↓2.507247079↑
Note:↑.Fold change>1.5,up-regulated expression;↓.Fold change<0.5,down-regulated expression.
表3 清絡(luò)通痹方具有下調(diào)干預(yù)作用miRNA的變化倍數(shù)
Tab.3 Fold changes of miRNAs down-regulated by QLT

miRNAFoldchange(CIA/Normal)Foldchange(QLT/CIA)mmu-miR-295-5p1.791609005↑0.23593644↓mmu-miR-449a-3p2.628849627↑0.186170898↓mmu-miR-679-5p2.671667893↑0.426010122↓mcmv-miR-m01-12.106437656↑0.29378167↓mmu-miR-3109-5p2.910775869↑0.398391097↓mmu-miR-493-5p1.851070637↑0.346558879↓
Note:↑.Fold change>1.5,up-regulated expression;↓.Fold change<0.5,down-regulated expression.

圖1 RT-PCR與miRNA芯片結(jié)果比較Fig.1 Comparison of miRNA fold-changes by miRNA microarray and real-time PCR

圖2 QLT對miR-143相對表達(dá)水平的影響Fig.2 Effects of QLT on the expression of miR-143 by real time PCRNote: *.P<0.05 vs normal;#.P<0.05 vs CIA.

圖3 miR-143潛在作用靶標(biāo)GO生物學(xué)過程富集分析Fig.3 Gene Ontology enrichment analysis of target genes of miR-143
2.4 mmu-miR-143生物信息學(xué)方法分析
2.4.1 mmu-miR-143的靶基因預(yù)測 利用軟件對所有靶基因分析,獲得microRNA-基因調(diào)控網(wǎng)絡(luò),應(yīng)用miRanda、TargetScan、miRRDB、miRWalk、RNA22預(yù)測 mmu-miR-143 的靶基因,取其三者交集的靶基因有1305個,其中結(jié)合文獻(xiàn)已證實的 mmu-miR-143靶標(biāo)有:FNDC3B、 MAPK7、 MAPK3、KRAS、COX2、COL1A1、MMP-2、MMP-13、IL13、IL3ra1、Casp3、TNF、Akt1等118個。
2.4.2 mmu-miR-143預(yù)測靶基因的功能注釋GO分析 根據(jù)預(yù)測靶基因集合進(jìn)行GO富集分析,發(fā)現(xiàn)有多個GO被miR-143調(diào)節(jié),mmu-miR-143靶基因主要參與了離子轉(zhuǎn)運、蛋白質(zhì)氨基酸磷酸化、骨骼系統(tǒng)發(fā)育、核糖核苷酸結(jié)合、ATP結(jié)合、細(xì)胞凋亡等生物過程(P<0.01),如圖3。
2.4.3 mmu-miR-143預(yù)測靶基因信號轉(zhuǎn)導(dǎo)通路的富集分析 根據(jù)預(yù)測出的靶基因集合,采用DAVID數(shù)據(jù)庫對其進(jìn)行基于 KEGG 數(shù)據(jù)庫的生物通路富集分析,發(fā)現(xiàn)mmu-miR-143參與了多個Pathway的調(diào)節(jié),其靶基因顯著富集在血管內(nèi)皮生長因子(Vascular endothelial growth factor,VEGF)、T細(xì)胞受體(T cell receptor)、絲裂原活化蛋白激酶(Mitogen-activated protein kinase ,MAPK)、神經(jīng)營養(yǎng)因子(Neurotrophin)、腫瘤等信號通路中(P<0.01),如圖4。

圖4 miR-143潛在作用靶標(biāo)信號轉(zhuǎn)導(dǎo)通路的富集分析Fig.4 KEGG enrichment analysis of target genes of miR-143
3 討論
RA 是一種以關(guān)節(jié)滑膜組織慢性炎癥為特點的自身免疫性疾病,發(fā)病原因迄今尚不明確,其發(fā)生可能與遺傳、慢性炎癥、自身免疫功能紊亂等因素有關(guān),miRNA的發(fā)現(xiàn)為RA的研究帶來了新的契機(jī)。MiRNA與靶基因結(jié)合后可介導(dǎo) mRNA 降解或者抑制其翻譯,一個miRNA可調(diào)控生物體內(nèi)多條基因的表達(dá),進(jìn)而參與細(xì)胞發(fā)育、分化、增殖、凋亡等生物學(xué)過程[5]。多種miRNA(如miR-23b、miR-146a、miR-155、miR-223、miR-203等)通過影響多種前炎癥因子的生成[6,7],干預(yù)MAPK等信號分子的表達(dá)[6-8],影響滑膜成纖維細(xì)胞增殖、遷移和破骨細(xì)胞分化與骨吸收過程[9],miRNA調(diào)控網(wǎng)絡(luò)對RA的發(fā)展起著至關(guān)重要的作用[1]。本研究結(jié)果顯示:與空白對照小鼠比較,CIA小鼠具有差異表達(dá)的microRNA有221個,提示多種miRNA的異常表達(dá)參與CIA的病理過程。
miRNA芯片結(jié)果提示與CIA組比較QLT組差異表達(dá)的microRNA有169個,QLT可能有干預(yù)作用的有49個miRNAs。其中多種miRNA功能涉及生物體免疫、炎癥,細(xì)胞周期調(diào)控、細(xì)胞凋亡及分化等生理病理過程,亦可能參與RA的發(fā)展。如Stanczyk等[10,11]證實miRNA-203、miR-19a/b可通過核轉(zhuǎn)錄因子κB、調(diào)節(jié)Toll樣受體通路影響基質(zhì)金屬蛋白酶(Matrix Metallo Preteinases,MMP)-1、MMP-3和IL-6的產(chǎn)生,miR-181調(diào)節(jié)T細(xì)胞的發(fā)育和成熟,影響單核細(xì)胞和巨噬細(xì)胞的炎癥反應(yīng)等[12],在RA滑膜炎癥發(fā)生和關(guān)節(jié)破壞方面發(fā)揮了重要作用。因此,QLT可能通過干預(yù)多種miRNA調(diào)控miRNA網(wǎng)絡(luò),多環(huán)節(jié)、多途徑干預(yù)CIA的多種病理過程。
根據(jù)miRNA芯片結(jié)果結(jié)合文獻(xiàn)查閱,miR-143在腫瘤方面的研究顯示其可調(diào)控MMP-13、MMP-2、MMP-9等細(xì)胞因子[13,14],Toll樣受體、MAPK/COX2 等信號分子[15,16],進(jìn)而影響細(xì)胞的增殖、遷移、侵襲和血管生成等過程。而在RA中多種細(xì)胞釋放IL、MMPs等細(xì)胞因子,并與VEGF、MAPK等多種信號蛋白交互作用,促進(jìn)滑膜炎、血管翳與骨破壞的發(fā)生[17,18]。此外Tam等[19]發(fā)現(xiàn) miR-143在傷害性感受刺激傳入的主要神經(jīng)元上高表達(dá),且CFA 炎性疼痛能夠引起 miR-143表達(dá)顯著下調(diào),提示其可能與炎性疼痛相關(guān)。結(jié)合結(jié)果及文獻(xiàn)首先集中對miR-143進(jìn)行驗證及分析,PCR結(jié)果顯示:CIA中miR-143下調(diào)為明顯,而QLT干預(yù)可上調(diào)miR-143的表達(dá)水平,提示miR-143可能是清絡(luò)通痹方調(diào)控 miRNA網(wǎng)絡(luò)的重要靶位之一。生物信息學(xué)分析發(fā)現(xiàn)miR-143 的靶基因中有VEGF、MAPKs、MMPs、TNF等,并參與了骨骼系統(tǒng)發(fā)育、蛋白質(zhì)氨基酸磷酸化、離子轉(zhuǎn)運結(jié)合、細(xì)胞凋亡等生物過程;信號通路顯著富集在VEGF、T細(xì)胞受體、MAPK、神經(jīng)營養(yǎng)因子信號通路等,均可能與RA炎癥、免疫、骨破壞等病理過程密切相關(guān)。QLT可能通過干預(yù)miR-143影響RA的免疫、炎癥、疼痛等多種病理過程。因此,miR-143可能作為復(fù)方治療RA的重要起始或中心環(huán)節(jié),與多種細(xì)胞因子和信號蛋白相關(guān),且具有網(wǎng)絡(luò)性調(diào)控的特點,因此可作為QLT復(fù)方機(jī)制研究的切入點。
綜上所述,多種miRNA的異常表達(dá)參與CIA的病理過程,QLT可能通過干預(yù)miRNA調(diào)控網(wǎng)絡(luò),多環(huán)節(jié)、多途徑干預(yù)關(guān)節(jié)炎免疫、炎癥、疼痛等多種病理過程,miR-143可能作為QLT治療RA的重要環(huán)節(jié)。本研究為進(jìn)一步認(rèn)識QLT干預(yù)膠原性關(guān)節(jié)炎機(jī)制提供新的線索,有助于深化對RA病理機(jī)制、治療策略的認(rèn)識。
[1] Salehi E,Eftekhari R,Oraei M,etal.MicroRNAs in rheumatoid arthritis[J].Clin Rheumatol,2015,34(4):615-628.
[2] Liu TY,Zhou LL,Zhou C,etal.Inhibition mechanism of Qingluo Tongbi Granule on osteoclast differentiation induced by synovial fibroblast and monocytes co-culture in adjuvant-induced arthritic rats[J].Chin J Integr Med,2015,4(4),291-298
[3] 周玲玲,柳璋璞,周 聰,等.清絡(luò)通痹顆粒通過MAPKs信號通路干預(yù)培養(yǎng)誘生的破骨樣細(xì)胞的分化[J].中國免疫學(xué)雜志,2013,29(12):1235-1239.
[4] Brand DD,Kang AH,Rosloniec EF.The mouse model of collagen-induced arthritis[M].Autoimmunity[J].Humana Press,2004:295-312.
[5] Carthew RW,Sontheimer EJ.Origins and mechanisms of miRNAs and siRNAs[J].Cell,2009,136(4):642-655.
[6] Niimoto T,Nakasa T,Ishikawa M,etal.MicroRNA-146a expresses in interleukin-17 producing T cells in rheumatoid arthritis patients[J].Bmc Musculoskeletal Disorders,2010,11(10):945-946.
[7] Kurowska-Stolarska M,Alivernini S,Ballantine LE,etal.MicroRNA-155 as a proinflammatory regulator in clinical and experimental arthritis[J].PNAS,2011,108(27):11193-11198.
[8] Murata K,Yoshitomi H,Furu M,etal.MicroRNA-451 down-regulates neutrophil chemotaxis via p38 MAPK[J].Arthritis Rheumatol,2014,66(3):549-559.
[9] Shibuya H,Nakasa T,Adachi N,etal.Overexpression of microRNA-223 in rheumatoid arthritis synovium controls osteoclast differentiation[J].Mod Rheumatol,2013,23:674-685.
[10] Stanczyk J,Ospelt C,Karouzakis E,etal.Altered expression of microRNA-203 in rheumatoid arthritis synovial fibroblasts and its role in fibroblast activation[J].Arthritis Rheumatol,2011,63(2):373-381.
[11] Philippe L,Alsaleh G,Suffert G,etal.TLR2 expression is regulated by microRNA miR-19 in rheumatoid fibroblast-like synoviocytes[J].J Immunol,2012,188(1):454-461.
[12] Xie W,Li M,Xu N,etal.MiR-181a regulates inflammation responses in monocytes and macrophages[J].Plos One,2013,8(3):e58639.
[13] Wu D,Huang P,Wang L,etal.MicroRNA-143 inhibits cell migration and invasion by targeting matrix metalloproteinase 13 in prostate cancer[J].Mol Med Rep,2013,8(2):626-630.
[14] Wang Q,Cai J,Wang J,etal.MiR-143 inhibits EGFR-signaling-dependent osteosarcoma invasion[J].Tumor Biol,2014,35(12):12743-12748.
[15] Guo H,Chen Y,Hu X,etal.The regulation of Toll-like receptor 2 by miR-143 suppresses the invasion and migration of a subset of human colorectal carcinoma cells[J].Molecular Cancer,2013,12(29):3477-3482.
[16] Hung P,Rodriguez CE,Graham W,etal.MiR-143 decreases COX-2 mRNA stability and expression in pancreatic cancer cells[J].Biochem Biophys Res Commun,2013,439(1):6-11.
[17] Schroeder M,Viezens L,Fuhrhop I,etal.Angiogenic growth factors in rheumatoid arthritis[J]. Rheumatol Int,2013,33(2):523-527.
[18] Criado G,Risco A,Alsina-Beauchamp D,etal.Alternative p38 mitogen-activated protein kinases are essential for collagen-induced arthritis[J].Arthritis Rheumatol,2014,66(5):1208-1217.
[19] Tam TS,Bastian I,Zhou XF,etal.MicroRNA-143 expression in dorsal root ganglion neurons[J].Cell Tissue Res,2011,346(2):163-173.
[收稿2015-07-24]
(編輯 許四平)
Study of Qingluo Tongbi Compound treating rheumatoid arthritis based on miRNA network
ZHU Ya-Mei,ZHOU Ling-Ling,PENG Xiao-Wu,GENG Shan,ZHOU Xue-Ping.Nanjing University of Chinese Medicine,Nanjing 210023,China
Objective:To study the mechanism of Qingluo Tongbi Compound(QLT) treating rheumatoid arthritis(RA)by observing miRNA Network of QLT on collagen-induced arthritis(CIA) mice.Methods: The model of RA was induced by collagenⅡ in DBA/1 mice and randomly divided into control group,CIA group,QLT group.Differently expressed miRNAs were detected by miRCURYTM LNA Array.Real-time PCR was applied to verify the reliability of miRNA array.Results: The bioinformatics software and database were applied to predict and analyze target genes.MiRNA array results showed that 221 miRNAs changed in CIA group compared with the control group,and 169 miRNAs changed in QLT group compared with CIA group.And the results of real-time PCR were consistent with the array.Compared with the control group,miR-143 was significantly reduced in CIA group,intervention of QLT obviously upregulated the expression of miR-143.The target genes of miR-143 were significantly stored in VEGF,T cell receptor,MAPK,signaling pathway.Conclusion: Multiple abnormal expression of miRNAs involved in the pathological process of CIA.QLT affected the expression of various miRNAs,which might be related to immunity,inflammation,pain pathological process of RA and miR-143 could be a potential target in the treatment.
microRNA;CIA;Qingluo Tongbi Compound(QLT)
10.3969/j.issn.1000-484X.2016.04.010
①本文受國家自然基金項目(81573869)、2014江蘇省普通高校研究生科研創(chuàng)新計劃(SJZZ-0122)資助和南京中醫(yī)藥大學(xué)國自然預(yù)研基金項目(14XYY01、14XYY10)。
朱亞梅(1989年-),女,在讀博士,主要從事中醫(yī)內(nèi)科學(xué)(風(fēng)濕病)研究,E-mail: zym-1220@163.com。
R285.5
A
1000-484X(2016)04-0495-05
②通訊作者,E-mail:zxp@njutcm.edu.cn。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學(xué)大眾(2021年6期)2021-07-20 07:42:44
科學(xué)(2020年3期)2020-11-26 08:18:30
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學(xué)學(xué)報(自然科學(xué)版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學(xué)院學(xué)報(2014年3期)2014-07-31 18:57:34