南京莫愁湖與紫霞湖浮游細(xì)菌群落結(jié)構(gòu)的季節(jié)性變化及其與環(huán)境因子的關(guān)系
2017-01-03 06:23:32曹新益徐慧敏王司辰何肖微余鐘波趙大勇
化學(xué)與生物工程 2016年12期
關(guān)鍵詞:差異結(jié)構(gòu)
曹新益,徐慧敏,王司辰,黃 睿,沈 烽,何肖微,余鐘波,趙大勇
(1.河海大學(xué) 水文水資源與水利工程科學(xué)國家重點實驗室,江蘇 南京 210098;2.河海大學(xué)水文水資源學(xué)院,江蘇 南京 210098)
南京莫愁湖與紫霞湖浮游細(xì)菌群落結(jié)構(gòu)的季節(jié)性變化及其與環(huán)境因子的關(guān)系
曹新益1,2,徐慧敏1,2,王司辰1,2,黃 睿1,2,沈 烽1,2,何肖微1,2,余鐘波1,2,趙大勇1,2
(1.河海大學(xué) 水文水資源與水利工程科學(xué)國家重點實驗室,江蘇 南京 210098;2.河海大學(xué)水文水資源學(xué)院,江蘇 南京 210098)
應(yīng)用16S rRNA基因末端限制性片段長度多態(tài)性(T-RFLP)技術(shù)和多元統(tǒng)計方法研究了南京莫愁湖和紫霞湖浮游細(xì)菌群落結(jié)構(gòu)的季節(jié)性變化及其與環(huán)境因子的關(guān)系。結(jié)果表明,莫愁湖和紫霞湖的浮游細(xì)菌群落alpha多樣性指數(shù)在夏季和秋季時最高,在春季和冬季時最低;紫霞湖冬季浮游細(xì)菌群落alpha多樣性指數(shù)顯著大于莫愁湖。非度量多維尺度(NMDS)分析結(jié)果顯示,莫愁湖和紫霞湖浮游細(xì)菌群落結(jié)構(gòu)存在明顯的季節(jié)性變化。Mantel檢驗、典范對應(yīng)分析(CCA)表明,影響莫愁湖和紫霞湖浮游細(xì)菌群落結(jié)構(gòu)最顯著的環(huán)境因子是溫度和pH值,營養(yǎng)物對莫愁湖和紫霞湖浮游細(xì)菌群落結(jié)構(gòu)的影響程度取決于湖泊的營養(yǎng)水平。為進一步研究不同程度富營養(yǎng)化水體中的浮游細(xì)菌群落結(jié)構(gòu)差異提供了幫助。
莫愁湖;紫霞湖;浮游細(xì)菌;群落結(jié)構(gòu);季節(jié)性變化;環(huán)境因子;多元統(tǒng)計;末端限制性片段長度多態(tài)性技術(shù)
莫愁湖是南京市一個重要的景觀類湖泊,近年隨著經(jīng)濟的發(fā)展,其生態(tài)環(huán)境遭到破壞,水質(zhì)屬劣Ⅴ類水平[1],已經(jīng)呈現(xiàn)較為嚴(yán)重的富營養(yǎng)化狀態(tài)。莫愁湖富營養(yǎng)化的主要限制因子為氮[2],近年來采取底泥疏浚、截污等措施,水質(zhì)有了明顯改善。紫霞湖是位于南京市中山陵風(fēng)景區(qū)的人工蓄水湖泊,外源營養(yǎng)物質(zhì)補給較少,為典型的低緯度貧營養(yǎng)型湖泊[2-3]。目前,對莫愁湖的富營養(yǎng)化、真核浮游生物、浮游植物、重金屬污染[2,4-6]和紫霞湖的浮游動物、浮游植物、重金屬污染[2,4,7]方面的研究報道較多,但對兩湖浮游細(xì)菌群落結(jié)構(gòu)的研究較少。
浮游生物作為水體生態(tài)系統(tǒng)中的優(yōu)勢種,是高等水生動植物賴以生存的重要物質(zhì)與能量基礎(chǔ),在生態(tài)系統(tǒng)中起著至關(guān)重要的作用[8]。浮游細(xì)菌是指浮游生物中的原核生物類群,主要包含古細(xì)菌、自養(yǎng)細(xì)菌和異養(yǎng)細(xì)菌[3]。浮游細(xì)菌對水體生態(tài)系統(tǒng)中食物鏈基本環(huán)節(jié)的發(fā)展有著決定性作用,浮游細(xì)菌在水體生態(tài)系統(tǒng)中的作用以及影響浮游細(xì)菌的環(huán)境因素受到研究者的廣泛關(guān)注[9]。區(qū)域特點和湖泊水體的營養(yǎng)水平均可以影響湖泊浮游細(xì)菌多樣性和群落結(jié)構(gòu)的組成,導(dǎo)致水體浮游細(xì)菌群落結(jié)構(gòu)的組成發(fā)生變化[10]。浮游細(xì)菌數(shù)量多,在污染物降解和氮磷等營養(yǎng)物循環(huán)過程中發(fā)揮著重要作用[11]。浮游細(xì)菌受外部環(huán)境的影響非常顯著[12],因此,浮游細(xì)菌群落結(jié)構(gòu)組成的改變可以在一定程度上反映該水體環(huán)境狀態(tài),研究表明浮游細(xì)菌在水質(zhì)調(diào)控中起著重要作用[13-14],目前對于浮游細(xì)菌群落在富營養(yǎng)化水體中的分布研究取得了很大進展[15-16]。
研究表明,水體生態(tài)系統(tǒng)中的營養(yǎng)鹽既可通過“上行效應(yīng)”直接對細(xì)菌群落的組成和分布造成影響[17-19],也可通過對水體細(xì)菌捕食者的組成和豐度的擾動間接影響水體細(xì)菌群落結(jié)構(gòu)和多樣性[20]。目前,越來越多的研究者開始關(guān)注浮游細(xì)菌與環(huán)境因子之間的關(guān)系[21-22],由于浮游細(xì)菌與環(huán)境因子的關(guān)系較為復(fù)雜,目前大部分研究仍在摸索階段[23]。Haukka等[24]和Wu等[25]研究發(fā)現(xiàn),在向淡水生態(tài)系統(tǒng)中添加營養(yǎng)物質(zhì)后,浮游細(xì)菌的群落結(jié)構(gòu)發(fā)生了變化,這種變化可能與水體營養(yǎng)水平的升高有關(guān),這表明淡水湖泊的營養(yǎng)狀態(tài)可能是影響浮游細(xì)菌群落結(jié)構(gòu)的重要因素。
作為降解者的微生物群落,浮游細(xì)菌個體結(jié)構(gòu)簡單,對環(huán)境變化敏感,其群落結(jié)構(gòu)組成和多樣性能夠在一定程度上反映水體環(huán)境質(zhì)量狀況[8],因此,調(diào)查年內(nèi)莫愁湖和紫霞湖中浮游細(xì)菌群落結(jié)構(gòu)組成的變化有助于更好地理解其生態(tài)系統(tǒng)。邊園琦等[3]對紫霞湖的浮游細(xì)菌群落結(jié)構(gòu)進行了研究,但并未研究紫霞湖浮游細(xì)菌群落結(jié)構(gòu)的季節(jié)性變化;沈烽等[26]應(yīng)用高通量測序技術(shù)對莫愁湖和紫霞湖的浮游細(xì)菌多樣性進行了研究,但并未探究浮游細(xì)菌群落結(jié)構(gòu)與環(huán)境因子之間的關(guān)系;Bai等[16]研究了高原湖泊滇池沉積物細(xì)菌多樣性,發(fā)現(xiàn)沉積物細(xì)菌多樣性的分布存在時空差異,并且受季節(jié)變化的影響;Ma等[27]研究粵東海水增養(yǎng)殖區(qū)浮游細(xì)菌數(shù)量變化時發(fā)現(xiàn),季節(jié)是影響浮游細(xì)菌數(shù)量的重要因素。季節(jié)變化會導(dǎo)致湖泊水體溫度、pH值等環(huán)境因子的變化,同時改變浮游植物光合作用強度,導(dǎo)致水體中有機物含量變化[28]。因此,研究湖泊浮游細(xì)菌群落結(jié)構(gòu)的季節(jié)性變化、探究導(dǎo)致這種變化的影響因子是必要的。
末端限制性片段長度多態(tài)性(terminal restriction fragment length polymorphism,T-RFLP)技術(shù)是一種新興的分子生物學(xué)技術(shù)[29-30],該技術(shù)不必進行細(xì)菌培養(yǎng)即可進行細(xì)菌群落分析[31],可以有效地應(yīng)用于微生物群落的對比分析和微生物多樣性分析[32]。自1997年Liu等[33]應(yīng)用T-RFLP技術(shù)研究微生物的多樣性以來,T-RFLP技術(shù)已被廣泛應(yīng)用于土壤[34-36]、植物根系[37]、污水處理[38]、湖泊沉積物[39]、淡水水體[40]等方面的研究。
作者應(yīng)用T-RFLP技術(shù)對南京莫愁湖和紫霞湖一年內(nèi)四個季節(jié)的浮游細(xì)菌群落結(jié)構(gòu)進行分析,研究2個湖泊浮游細(xì)菌群落結(jié)構(gòu)的季節(jié)差異,通過多元統(tǒng)計方法研究浮游細(xì)菌群落結(jié)構(gòu)與環(huán)境因子的關(guān)系,探討影響莫愁湖和紫霞湖浮游細(xì)菌群落結(jié)構(gòu)的關(guān)鍵環(huán)境因子,擬為莫愁湖和紫霞湖污染水體的生態(tài)修復(fù)提供理論依據(jù),豐富對城市湖泊水體浮游細(xì)菌多樣性、浮游細(xì)菌群落結(jié)構(gòu)與環(huán)境因子相互關(guān)系的認(rèn)識。
1 實驗
1.1 水樣采集
在2011年1月至2012年1月間,每個季節(jié)對莫愁湖和紫霞湖各采集一次水樣,每次采集3~4個平行樣本,采樣點均勻分布在湖區(qū)內(nèi)。
使用采水器采集距水面0.5 m處的水樣1 000 mL,分裝在滅菌藍蓋玻璃瓶中(期間現(xiàn)場測定水樣的溫度及pH值),密封,避光保存在4 ℃保溫箱中,盡快運回實驗室進行后續(xù)處理。
1.2 理化指標(biāo)分析
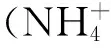
1.3 T-RFLP分析
取上述經(jīng)過0.22 μm濾膜過濾的水樣,參照文獻[39]進行DNA提取。PCR擴增反應(yīng)采用細(xì)菌通用引物對27f(5′-AGAGTTTGATCCTGGCTCAG-3′)和926r(5′-CCGTCAATTCCTTTGAGTTT-3′)。PCR反應(yīng)體系和循環(huán)條件參照文獻[41]。擴增體系為50 μL,包含 5×PCR buffer 10 μL、0.25 mmol·L-1dNTP、2.5 mmol·L-1鎂離子、0.2 mmol·L-1上下游引物、10 ng DNA模板、2 U Taq酶,加去離子水至50 μL。擴增程序為:95 ℃預(yù)變性3 min;94 ℃變性1 min,55 ℃退火1 min,72 ℃延伸1 min,循環(huán)30次;72 ℃延伸7 min。4 ℃保存,進行下游分析。
PCR擴增產(chǎn)物在2%瓊脂糖凝膠電泳檢測后使用核酸片段分析儀(CEQ8000型,Beckman Coulter,Fullerton,CA,USA)對酶切產(chǎn)物進行T-RFLP分析,分析時控制堿基長度介于60~600 bp、相對豐度超過1%的片段的末端限制性片段,即T-RFs。按T-RFs占各個樣品的豐度百分比進行標(biāo)準(zhǔn)化,再分別計算每個樣品浮游細(xì)菌群落的alpha多樣性指數(shù)(Simpson指數(shù)和Shannon指數(shù)),以研究水體中浮游細(xì)菌群落的多樣性和均一度。
對每組樣品的多樣性指數(shù)做anova檢驗,用a、b、c、d分別標(biāo)注多樣性指數(shù)有顯著差異的樣品(P<0.05)。
1.4 alpha多樣性指數(shù)分析
采用Shannon(H′)和Simpson指數(shù)(D)進行浮游細(xì)菌群落的多樣性評價[39]。
(1)
(2)
式中:S為物種總數(shù);Pi為第i個峰面積占總面積的比例,即T-RFs的相對豐度。
1.5 多元統(tǒng)計分析
利用R語言軟件(R programming language,version 3.2.3)中的Vegan軟件包完成如下分析:非度量多維尺度(non-metric multidimensional scaling,NMDS)分析、典范對應(yīng)分析(canonical correlation analysis,CCA)、envfit分析、Mantel及偏Mantel檢驗、方差分解分析(variation partitioning analysis,VPA)。其中,NMDS分析用于判定不同樣品間浮游細(xì)菌群落結(jié)構(gòu)的差異性,CCA用于研究環(huán)境因子對浮游細(xì)菌群落結(jié)構(gòu)的影響,Mantel及偏Mantel檢驗用于探究環(huán)境差異與浮游細(xì)菌群落結(jié)構(gòu)差異的相關(guān)性,VPA用于研究不同環(huán)境因子對浮游細(xì)菌群落差異的影響程度。
2 結(jié)果與討論
2.1 水樣理化指標(biāo)分析(表1)
表1 莫愁湖和紫霞湖水樣理化指標(biāo)分析
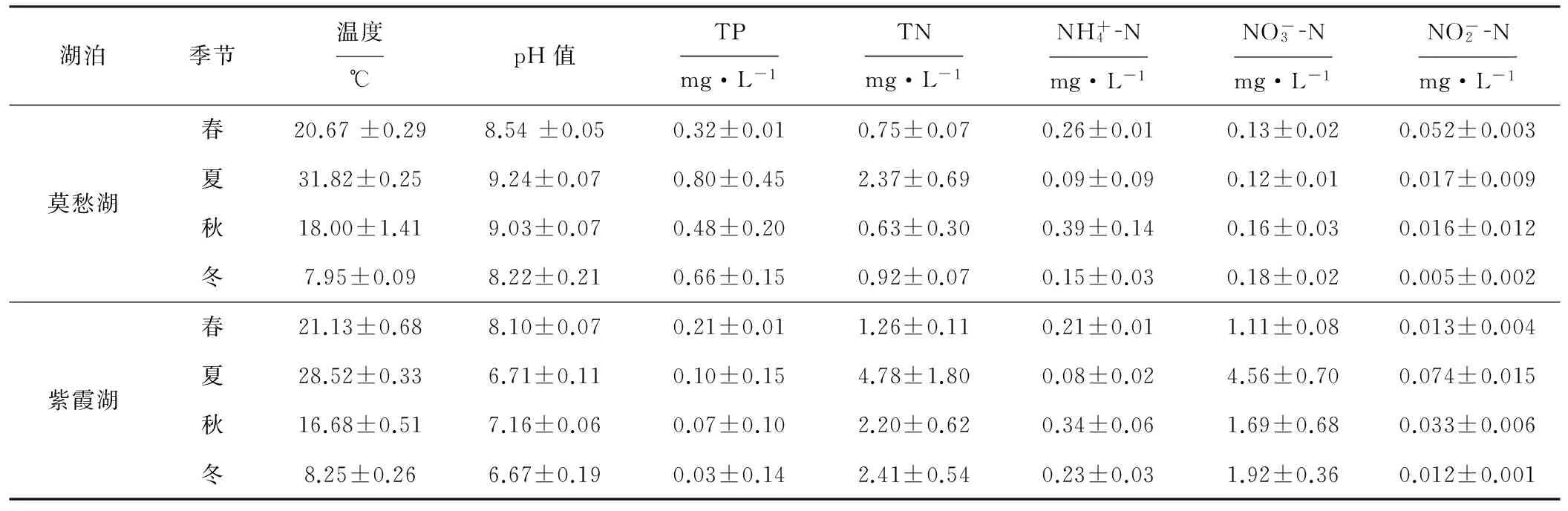
Tab.1 Physicochemical parameters of water samples from Mochou Lake and Zixia Lake
依Thomas標(biāo)準(zhǔn)[40]:“富”級:無機氮>1.5 mg·L-1,TP>0.1 mg·L-1;“中-富”級:無機氮0.5~1.5 mg·L-1,TP 0.03~0.1 mg·L-1。從表1可以看出,莫愁湖的TN在4個季節(jié)都達到了“中-富”級水平,TP都達到了“富”級水平,而紫霞湖的TN在4個季節(jié)都達到了“富”級水平,TP則為“中-富”級水平;2個湖泊的TP在夏季明顯升高(分別達到0.80 mg·L-1和0.10 mg·L-1),莫愁湖的TP整體上高于紫霞湖;2個湖泊的TN整體而言比較穩(wěn)定,均在夏季達到最高值(分別達到2.37 mg·L-1和4.78 mg·L-1),紫霞湖的TN整體上高于莫愁湖。這是因為,莫愁湖作為景觀湖泊,其主要污染源為生活污水和生活垃圾,污染程度更加嚴(yán)重;紫霞湖作為風(fēng)景區(qū)的人工蓄水湖泊,污染程度相對較輕。
2.2 alpha多樣性指數(shù)分析
根據(jù)T-RFLP圖譜結(jié)果,分別計算不同季節(jié)莫愁湖和紫霞湖浮游細(xì)菌群落的alpha多樣性指數(shù),結(jié)果如圖1所示。
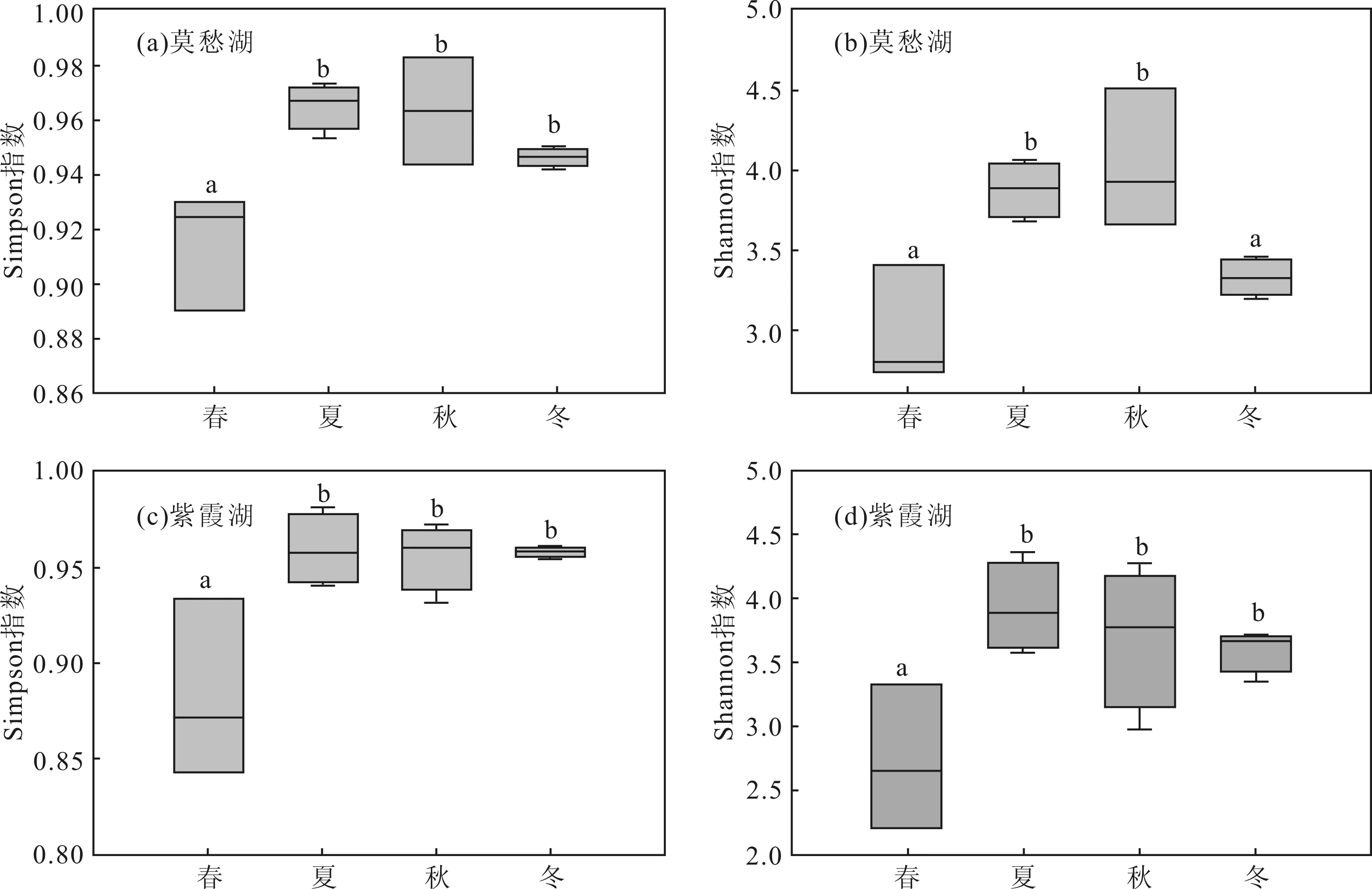
圖中不同的字母標(biāo)注表示2個組有顯著性差異(P<0.05,anova檢驗)
從圖1 中Simpson指數(shù)(圖1a、c)可以看出,春季莫愁湖和紫霞湖的浮游細(xì)菌群落的多樣性均要顯著低于其它3個季節(jié)。Shannon指數(shù)(圖1b、d)顯示,春季莫愁湖的浮游細(xì)菌群落的多樣性要顯著低于夏季和秋季,與冬季無顯著性差異;而紫霞湖浮游細(xì)菌群落多樣性要顯著低于其它3個季節(jié),這與沈烽等[26]的研究結(jié)果是一致的。
不同季節(jié)莫愁湖和紫霞湖浮游細(xì)菌群落的alpha多樣性指數(shù)對比如圖2所示。
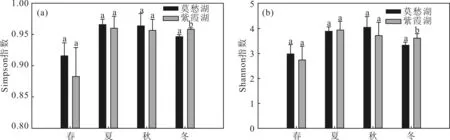
圖中不同的字母標(biāo)注表示2個組有顯著性差異(P<0.05,T檢驗)
從圖2中Simpson指數(shù)和Shannon指數(shù)可以看出,莫愁湖和紫霞湖在春季、夏季和秋季的浮游細(xì)菌群落多樣性無顯著性差異,冬季的浮游細(xì)菌群落多樣性有顯著性差異。這個結(jié)果可能與2個湖泊不同的營養(yǎng)類型有關(guān)[24]。
2.3 不同季節(jié)浮游細(xì)菌群落結(jié)構(gòu)差異
通過計算樣品間T-RFLP的距離(Bray-Curtis距離),采用NMDS分析方法探究不同季節(jié)莫愁湖和紫霞湖浮游細(xì)菌群落結(jié)構(gòu)的可能差異,結(jié)果如圖3所示。
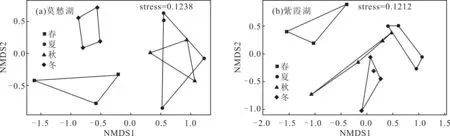
圖中每一個點表示一個樣品
圖3 不同季節(jié)莫愁湖和紫霞湖浮游細(xì)菌群落結(jié)構(gòu)的NMDS分析
Fig.3 NMDS analysis of bacterioplankton community structure of Mochou Lake and Zixia Lake in four seasons
從圖3a可以看出,莫愁湖春季、冬季和夏秋季的點重疊很小,夏季與秋季的點重疊較大,春季與冬季的點則無重疊,表明莫愁湖夏季、秋季的浮游細(xì)菌群落結(jié)構(gòu)比較相似,春季、夏秋季和冬季的浮游細(xì)菌群落結(jié)構(gòu)有比較顯著的差異。從圖3b可以看出,紫霞湖夏季、秋季和冬季的點相對接近,春季的點相對獨立,表明紫霞湖夏季、秋季和冬季的浮游細(xì)菌群落結(jié)構(gòu)比較相似,春季的浮游細(xì)菌群落結(jié)構(gòu)與其它3個季節(jié)有較明顯的差異。
2.4 浮游細(xì)菌群落結(jié)構(gòu)與環(huán)境因子的關(guān)系
2.4.1 CCA
莫愁湖與紫霞湖的數(shù)據(jù)CCA排序前四軸最大值分別為2.94和3.92,為了便于比較,因此均使用單峰模型[26]。使用CCA探究環(huán)境變量對浮游細(xì)菌群落結(jié)構(gòu)的影響,其中環(huán)境因子用帶箭頭的直線表示,直線越長則此環(huán)境因子與浮游細(xì)菌群落結(jié)構(gòu)的相關(guān)性越大,直線和排序軸的夾角越小則此環(huán)境因子與排序軸的相關(guān)性越大。如圖4所示。
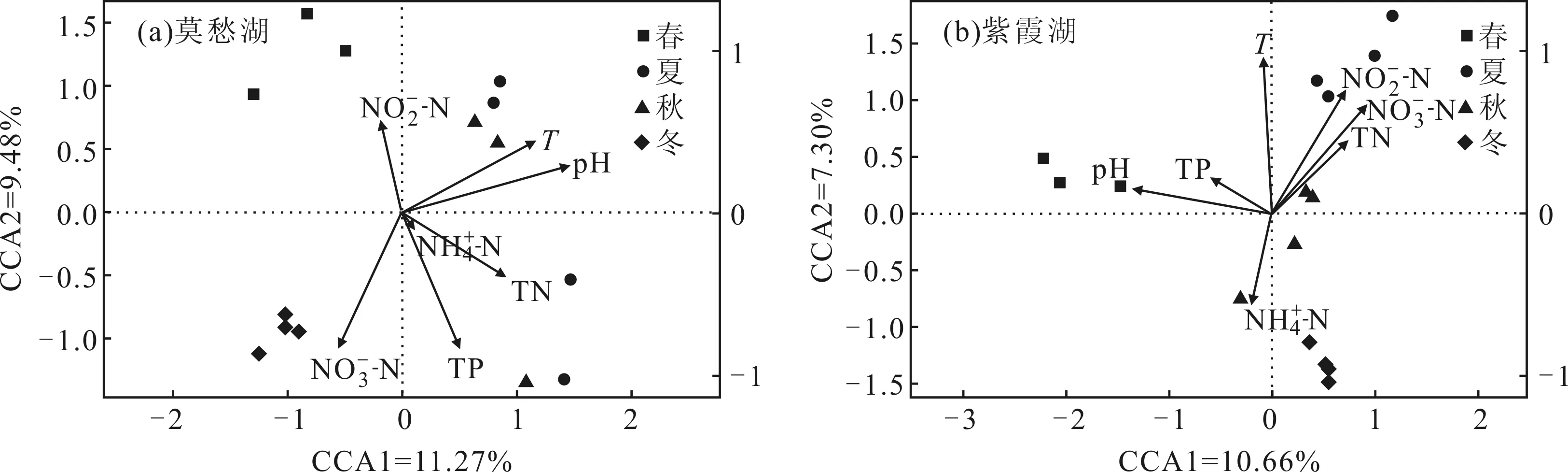
圖4 莫愁湖和紫霞湖浮游細(xì)菌群落結(jié)構(gòu)與環(huán)境因子的典范對應(yīng)分析
從圖4可看出,莫愁湖的排序軸1軸和2軸分別解釋了11.27%和9.48%的樣本變異,紫霞湖的排序軸1軸和2軸分別解釋了10.66%和7.30%的樣本變異。
2.4.2 envfit分析
使用envfit分析檢驗每個環(huán)境因子的顯著性,結(jié)果如表2所示。
表2 環(huán)境因子的CCA排序軸的相關(guān)性分析
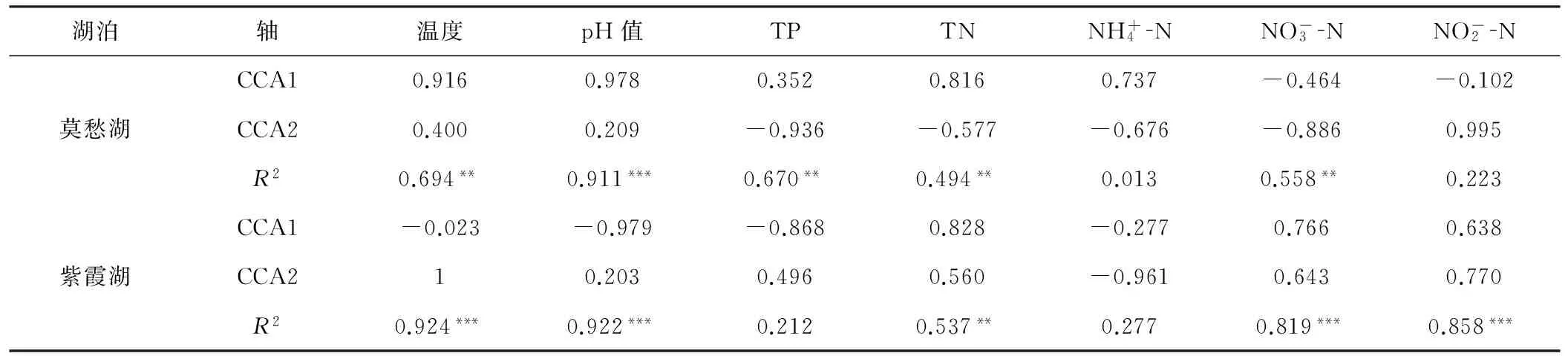
Tab.2 Correlation analysis of CCA ordination axises for environmental factors
注:顯著水平基于1000次置換;***表示P<0.001,極顯著;**表示P<0.01,較顯著;*表示P<0.05,顯著。下表同。
2.4.3 Mantel檢驗
使用Mantel檢驗分析樣品浮游細(xì)菌群落結(jié)構(gòu)差異與環(huán)境變量差異的相關(guān)性。樣品間浮游細(xì)菌群落結(jié)構(gòu)差異通過計算Bray-Curtis距離來確定,樣品間環(huán)境變量差異通過歐氏距離來確定,結(jié)果如表3所示。
表3 浮游細(xì)菌群落結(jié)構(gòu)和環(huán)境變量的Mantel檢驗
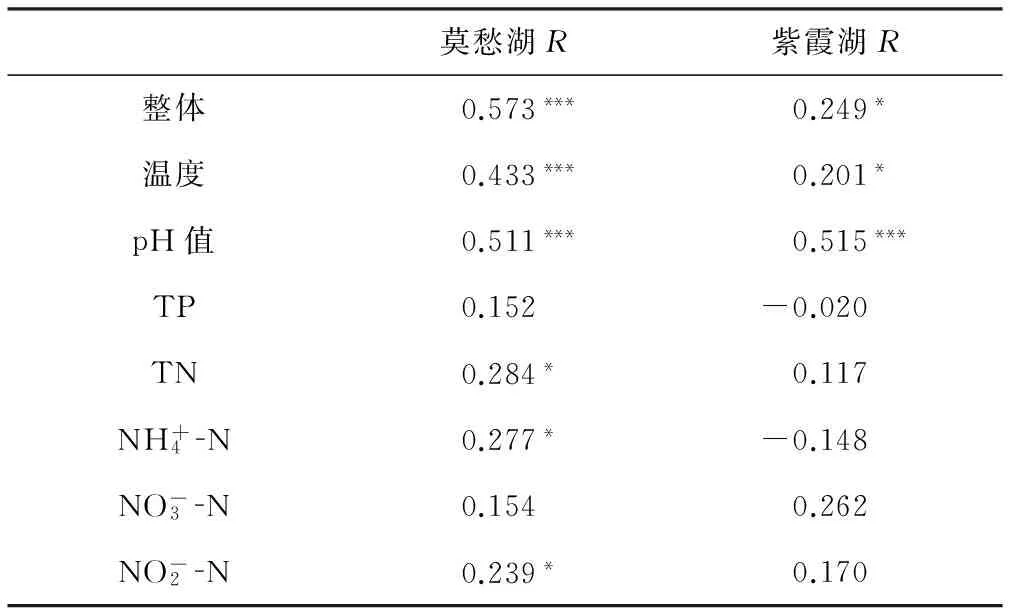
Tab.3 Mantel test for the bacterioplankton community structure and environmental variables
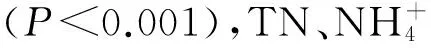
2.4.4 偏Mantel檢驗
偏Mantel檢驗是控制其它變量,分析一個變量與另一個變量的關(guān)系。浮游細(xì)菌群落結(jié)構(gòu)和環(huán)境變量的偏Mantel檢驗結(jié)果如表4所示。
2.5 討論
(1)通過T-RFLP技術(shù)研究了南京莫愁湖和紫霞湖浮游細(xì)菌群落結(jié)構(gòu)隨季節(jié)的變化。alpha多樣性指數(shù)分析表明,不同季節(jié)湖泊浮游細(xì)菌群落的多樣性指數(shù)是不同的。莫愁湖和紫霞湖的浮游細(xì)菌群落的Simpson指數(shù)在春季最低,而在夏季、秋季和冬季差異不大。這一方面是因為夏秋季節(jié)湖泊水體中的大型植物和浮游植物大量繁殖,光合作用產(chǎn)生的大量有機物為細(xì)菌自由生長繁殖提供了物質(zhì)基礎(chǔ);另一方面是因為冬春季水體溫度相對較低,抑制了細(xì)菌的新陳代謝和生長增殖速率,而夏秋季則相反[28]。2個湖泊春季、夏季和秋季的alpha多樣性指數(shù)沒有顯著性差異,紫霞湖冬季alpha多樣性指數(shù)要顯著高于莫愁湖,主要原因可能是:莫愁湖作為景觀湖泊受人類活動影響較大,一年四季均有污水排入[6],導(dǎo)致莫愁湖一年四季均處在較高的營養(yǎng)狀態(tài);紫霞湖作為人工蓄水湖泊受人類活動影響較小,1月份是枯水期,紫霞湖在此期間山間外來水流入湖泊較少,菌群未能富集,水體的浮游細(xì)菌多樣性相應(yīng)較高,而4月份處于豐水期,在此期間有大量外來水流入湖泊,菌群富集,水體的浮游細(xì)菌多樣性相應(yīng)降低[42]。劉蘭英等[42]對福州左海湖的研究,高程等[43]對北京城區(qū)湖泊浮游細(xì)菌的研究,邢鵬等[44]、秦伯強等[45]和Wu等[25]對太湖浮游細(xì)菌的研究也得到了類似的結(jié)論。
表4 浮游細(xì)菌群落結(jié)構(gòu)和環(huán)境變量的偏Mantel檢驗
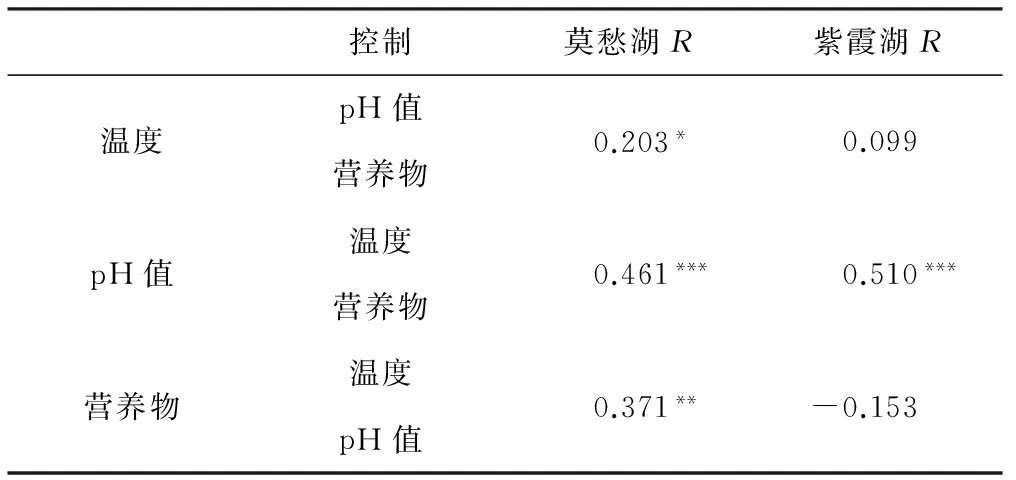
Tab.4 Partial Mantel test for the bacterioplankton community structure and environmental variables
(2)采用以Bray-Curtis距離為基礎(chǔ)的NMDS分析探究莫愁湖和紫霞湖浮游細(xì)菌群落結(jié)構(gòu)隨季節(jié)的變化。結(jié)果表明,莫愁湖浮游細(xì)菌群落結(jié)構(gòu)在夏季和秋季差異不大,而春季、冬季和夏秋季的浮游細(xì)菌群落結(jié)構(gòu)差異較大,這與沈烽等[26]對太湖浮游細(xì)菌群落結(jié)構(gòu)的研究結(jié)果是一致的;紫霞湖浮游細(xì)菌群落結(jié)構(gòu)在夏季、秋季和冬季差異不大,春季與其它季節(jié)差異較大。
(3)運用CCA方法探究莫愁湖和紫霞湖浮游細(xì)菌群落結(jié)構(gòu)與環(huán)境因子的關(guān)系。結(jié)果表明,莫愁湖和紫霞湖的溫度、pH值對浮游細(xì)菌群落結(jié)構(gòu)均有著極其重要的影響,營養(yǎng)物有一定影響但不如前兩者,尤其是對于紫霞湖的影響。研究表明,浮游細(xì)菌群落結(jié)構(gòu)和多樣性與空間[46]、時間[42-43]、環(huán)境變量(如溫度、pH值和營養(yǎng)物)有關(guān)[47],這與本研究結(jié)果是一致的。Mantel檢驗結(jié)果表明,莫愁湖的溫度差異和pH值差異對浮游細(xì)菌群落結(jié)構(gòu)的差異有著極顯著的影響(P<0.001),莫愁湖的營養(yǎng)物差異的影響顯著(P<0.05),但紫霞湖的營養(yǎng)物差異的影響并不顯著,這可能與2個湖泊的營養(yǎng)水平不同有關(guān)[48]。
研究表明,①在營養(yǎng)水平較低的湖泊中,營養(yǎng)物會通過“上行效應(yīng)”直接影響水體浮游細(xì)菌群落的生長[18-19,49]。②pH值會顯著影響湖泊水體浮游細(xì)菌群落結(jié)構(gòu)和多樣性[49-50],一方面,不同梯度的pH值會不同程度地影響各類細(xì)菌的生長狀況(包括絕滅、繁衍、種的形成等[51]),從而直接影響浮游細(xì)菌群落結(jié)構(gòu)和多樣性;另一方面,不同的pH值梯度會不同程度地影響湖泊生態(tài)系統(tǒng)中的其它環(huán)境因子,從而間接影響水體浮游細(xì)菌群落結(jié)構(gòu)和多樣性。③溫度對水體浮游細(xì)菌群落結(jié)構(gòu)具有選擇作用,是影響湖泊浮游細(xì)菌群落結(jié)構(gòu)的關(guān)鍵因子之一[52-54]。
本研究中,各采樣點屬于弱堿性環(huán)境,適合大部分細(xì)菌生長,同時也存在一定的pH值梯度,而溫度在一年內(nèi)的變化十分明顯,因此會對浮游細(xì)菌群落結(jié)構(gòu)有一定的選擇性。
(4)T-RFLP技術(shù)具有分析精細(xì)、分析能力強、易于自動化、分辨率高和可重復(fù)性等優(yōu)點[55],被廣泛應(yīng)用于大范圍長時間尺度的微生物群落結(jié)構(gòu)研究[26]。但若不注意其諸多過程中的技術(shù)細(xì)節(jié),有可能引起分析誤差,導(dǎo)致無法完全準(zhǔn)確地反映環(huán)境樣品中細(xì)菌群落的結(jié)構(gòu)和多樣性信息。因而為了確保分析結(jié)果的真實可靠,在實驗設(shè)計時需采用多個具有代表性的重復(fù)樣品,使用相同方法重復(fù)提取各樣品總DNA,提高分析結(jié)果的可靠性[56]。
3 結(jié)論
應(yīng)用16S rRNA基因末端限制性片段長度多態(tài)性(T-RFLP)技術(shù)和多元統(tǒng)計方法研究了南京莫愁湖和紫霞湖浮游細(xì)菌群落結(jié)構(gòu)的季節(jié)性變化及其與環(huán)境因子的關(guān)系。結(jié)果表明,莫愁湖和紫霞湖的alpha多樣性指數(shù)在夏季和秋季時最高,在春季和冬季時最低;紫霞湖冬季浮游細(xì)菌群落的alpha多樣性指數(shù)顯著大于莫愁湖。非度量多維尺度(NMDS)分析結(jié)果顯示,莫愁湖和紫霞湖浮游細(xì)菌群落結(jié)構(gòu)存在明顯的季節(jié)性變化。Mantel檢驗、典范對應(yīng)分析(CCA)表明,影響莫愁湖和紫霞湖浮游細(xì)菌群落結(jié)構(gòu)最顯著的環(huán)境因子是溫度和pH值,營養(yǎng)物對莫愁湖和紫霞湖浮游細(xì)菌群落結(jié)構(gòu)的影響程度取決于湖泊的營養(yǎng)水平。為進一步研究不同程度富營養(yǎng)化水體中的浮游細(xì)菌群落結(jié)構(gòu)差異提供了幫助。
[1] 趙大勇,王猛,曾巾,等.南京莫愁湖沉積物中重金屬污染調(diào)查[J].河海大學(xué)學(xué)報(自然科學(xué)版),2012,40(5):520-524.
[2] 唐雅萍,張哲海,梅卓華,等.南京市主要湖泊浮游植物群落結(jié)構(gòu)分析[J].環(huán)境科技,2008,21(5):14-16.
[3] 邊園琦,邢鵬,李化炳,等.貧營養(yǎng)湖泊花神湖和紫霞湖浮游細(xì)菌群落分析[J].生態(tài)科學(xué),2010,29(2):109-115.
[4] 趙璧影,陳美軍,孫穎,等.南京8個湖泊超微真核浮游生物遺傳多樣性的研究[J].環(huán)境科學(xué),2010,31(5):1293-1298.
[5] 吳剛剛,吳曉霞,陳昌云,等.莫愁湖底泥重金屬污染評價[J].中國教育技術(shù)裝備,2011(9):104-105.
[6] 顧進偉,錢誼,黃輝,等.莫愁湖富營養(yǎng)化限制性因素確定的AGP實驗研究[J].南京師大學(xué)報(自然科學(xué)版),2014,37(3):111-115.
[7] 呂志均,戴曦,孫穎,等.紫霞湖浮游甲殼動物晝夜垂直遷移及其影響因子分析[J].水生態(tài)學(xué)雜志,2011,32(3):43-47.
[8] 劉樂冕,楊軍,余小青,等.廈門后溪流域沿城鄉(xiāng)梯度浮游細(xì)菌多樣性及其與環(huán)境因子的關(guān)系[J].應(yīng)用與環(huán)境生物學(xué)報,2012,18(4):591-598.
[9] RAYMOND P A,BAUER J E.Bacterial consumption of DOC during transport through a temperate estuary[J].Aquatic Microbial Ecology,2000,22(1):1-12.
[10] ZWART G,GRUMP B C,AGTERVELD M P K V,et al.Typical freshwater bacteria :an analysis of available 16S rRNA gene sequences from plankton of lakes and rivers[J].Aquatic Microbial Ecology,2002,28(2):141-155.
[11] MUYLAERT K,van der GUCHT K,VLOEMANS N,et al.Relationship between bacterial community composition and bottom-up versus top-down variables in four eutrophic shallow lakes[J].Applied and Environmental Microbiology,2000,68(10):4740-4750.
[12] PAER H W,DYBLE J,MOISANDER P H,et al.Microbial indicators of aquatic ecosystem change:current applications to eutrophication studies[J].FEMS Microbiological Ecology,2003,46(3):233-246.
[13] HAHN M W.The microbial diversity of inland waters[J].Current Opinion in Biotechnology,2006,17(3):256-261.
[14] 周純,宋春雷,曹秀云,等.太湖不同解有機磷菌株胞外堿性磷酸酶活性對藍藻碎屑的響應(yīng)[J].水生生物學(xué)報,2012,36(1):119-125.
[15] DUFOUR P,TORRETON J P,COLON M.Advantages of distinguishing the active fraction in baeterioplankton assemblages: some examples[J].Hydrobiologia,1990,207(1):295-301.
[16] BAI Y,SHI Q,WEN D,et al.Bacterial communities in the sediments of Dianchi Lake,a partitioned eutrophic waterbody in China[J].PLoS One,2012,7(5):e37796.
[17] CRUMP B C,KLING G W,BAHR M,et al.Bacterioplankton community shifts in an arctic lake correlate with seasonal changes in organic matter source[J].Applied and Environmental Microbiology,2003,69(4):2253-2268.
[18] PEREZ M T,SOMMARUGA R.Differential effect of algal- and soil-derived dissolved organic matter on alpine lake bacterial community composition and activity[J].Limnology and Oceanography,2006,51(6):2527-2537.
[19] BOUVY M,BETTAREl Y,BOUVIER C,et al.Trophic interactions between viruses,bacteria and nanoflagellates under various nutrient conditions and simulated climate change[J].Environmental Microbiology,2011,13(7):1842-1857.
[20] NAKANO S I,ISHII N,MANAGE P M,et al.Trophic roles of heterotrophic nanoflagellates and ciliates among planktonic organisms in a hypereutrophic pond[J].Aquatic Microbial Ecology,1998,16(2):153-161.
[21] DONNER G,SCHWARZ K,HOPPE H G,et al.Profiling the succession of bacterial populations in pelagic chemoclines[J].Arch Hydrobiol Spec Issues Advance Limnol,1996,48:7-14.
[22] LIU Y Q,YAO T D,ZHU L P,et al.Bacterial diversity of freshwater alpine Lake Puma Yumco on the Tibetan Plateau[J].Journal of Genomicrobiology,2009,26(2):131-165.
[23] VIEIRA R P,GINZALEZ A M,CARDOSO A M,et al.Relationships between bacterial diversity and environmental variables in a tropical marine environment[J].Environmental Microbiology,2008,10(1):189-199.
[24] HAUKKA K,KOLMONEN E,HYDER R,et al.Effect of nutrient loading on bacterioplankton community composition in lake mesocosms[J].Microbial Ecology,2006,51(2):137-146.
[25] WU Q L,CHEN Y W,XU K D,et al.Intra-habitat heterogeneity of microbial food web structure under the regime of eutrophication and sediment resuspension in the large subtrophical shallow Lake Taihu,China[J].Hydrobiologia,2007,581(1):241-254.
[26] 沈烽,黃睿,王司辰,等.不同OTUs劃分閾值對湖泊浮游細(xì)菌多樣性的影響[J].化學(xué)與生物工程,2016,33(2):25-30.
[27] 馬繼波,董巧香,黃長江.粵東大規(guī)模海水增養(yǎng)殖區(qū)柘林灣浮游細(xì)菌的時空分布[J].生態(tài)學(xué)報,2007,27(2):477-485.
[28] 馮勝,高光,秦伯強,等.太湖北部湖區(qū)水體中浮游細(xì)菌的動態(tài)變化[J].湖泊科學(xué),2006,18(6):636-642.
[29] HORTON T R,BRUNS T D.The molecular revolution in ectomycorrhizal ecology:peeking into the black-box[J].Molecular Ecology,2001,10(8):1855-1871.
[30] BRIDGE P,SPOONER B.Soil fungi:diversity and detection[J].Plant and Soil,2001,232(1):147-154.
[31] 余素林,吳曉磊,錢易.環(huán)境微生物群落分析的T-RFLP技術(shù)及其優(yōu)化措施[J].應(yīng)用與環(huán)境生物學(xué)報,2006,12(6):861-868.
[32] 王洪媛,管華詩,江曉路.微生物生態(tài)學(xué)中分子生物學(xué)方法及T-RFLP技術(shù)研究[J].中國生物工程雜志,2004,24(8):42-47.
[33] LIU W T,MARSH T L,CHENG H,et al.Characterization of microbial diversity by determining terminal restriction fragment length polymorphisms of genes encoding 16S rRNA[J].Applied and Environmental Microbiology,1997,63(11):4516-4522.
[34] BUCHAN A,NEWELL S Y,BULTER M,et al.Dynamics of bacterial and fungal communities on decaying salt marsh grass[J].Applied and Environmental Microbiology,2003,69(11):6676-6687.
[35] TOM-PETERSEN A,LESER T D,MARSH T L,et al.Effects of copper amendment on the bacterial community in agricultural soil analyzed by the T-RFLP technique[J].FEMS Microbiology Ecology,2003,46(1):53-62.
[36] 尹承苗,王功帥,李園園,等.連作蘋果園土壤真菌的T-RFLP分析[J].生態(tài)學(xué)報,2014,34(4):837-846.
[37] 馬棟山,熊薇,張瓊瓊,等.基于T-RFLP和因子分析的香蒲根際細(xì)菌群落研究[J].中國環(huán)境科學(xué),2014,34(10):2684-2691.
[38] 王曉慧,文湘華,丁鹍,等.T-RFLP方法分析城市污水處理廠中細(xì)菌群落的動態(tài)變化[J].環(huán)境科學(xué),2010,31(5):1307-1312.
[39] 趙大勇,孫一萌,方超,等.太湖梅梁灣不同深度沉積物中細(xì)菌群落結(jié)構(gòu)組成[J].河海大學(xué)學(xué)報(自然科學(xué)版),2013,41(4):283-287.
[40] CHIN K J,LUEDERS T,FRIEDRICH M W,et al.Archaeal community structure and pathway of methane formation on rice roots[J].Microbial Ecology,2004,47(1):59-67.
[41] ZENG J,YANG L Y,LIANG Y,et al.Spatial distribution of bacterial communities in sediment of a eutrophic lake revealed by denaturing gradient gel electrophoresis and multivariate analysis[J].Canadian Journal of Microbiology,2008,54(12):1053-1063.
[42] 劉蘭英,呂新,李巍,等.福州左海湖冬春季浮游細(xì)菌群落16S rRNA多樣性[J].微生物學(xué)報,2014,54(2):200-210.
[43] 高程,黃滿榮,陶爽,等.北京城區(qū)不同水質(zhì)水體可培養(yǎng)細(xì)菌數(shù)量的季節(jié)動態(tài)變化[J].生態(tài)學(xué)報,2011,31(4):1157-1163.
[44] 邢鵬,孔繁翔,高光.太湖浮游細(xì)菌種群基因多樣性及其季節(jié)變化規(guī)律[J].湖泊科學(xué),2007,19(4):373-381.
[45] 秦伯強,胡維平,陳偉民,等.太湖水環(huán)境演化過程與機理[M].北京:科學(xué)出版社,2004:248-313.
[46] LINDSTR?N E S,EILER A,LANGENHEDER S,et al.Does ecosystem size determine aquatic bacterial richness?[J].Ecology,2007,88(1):252-253.
[47] 任麗娟,何聃,邢鵬,等.湖泊水體細(xì)菌多樣性及其生態(tài)功能研究進展[J].生物多樣性,2013,21(4):421-432.
[48] CHRZANOWSKI T H,STERNER R W,ELSER J J.Nutrient enrichment and nutrient regeneration stimulate bacterioplankton growth[J].Microbial Ecology,1995,29(3):221-230.
[49] LINDSTROM E S,KAMST-van AGTERVELD M P,ZWART G,et al.Distribution of typical freshwater bacterial groups is associated with pH,temperature,and lake water retention time[J].Applied and Environmental Microbiology,2005,71(12):8201-8206.
[50] YANNARELL A C,TRIPLETT E W.Geographic and environmental sources of variation in lake bacterial community composition[J].Applied and Environmental Microbiology,2005,71(2):227-239.
[51] LANGENHEDER S,LINDSTROM E S,TRANVIK L J.Structure and function of bacterial communities emerging from different sources under identical conditions[J].Applied and Environ-mental Microbiology,2006,72:212-220.
[52] SIMON M,WINSCH C.Temperature control of bacterioplankton growth in a temperate large lake[J].Aquatic Microbial Ecology,1998,16(2):119-130.
[53] ADAMS H E,CRUMP B C,KLING G W.Temperature controls on aquatic bacterial production and community dynamics in arctic lakes and streams[J].Environmental Microbiology,2010,12(5):1319-1333.
[54] DZIALLAS C,GROSSART H P.Temperature and biotic factors influence bacterial communities associated with the cyanobacteriumMicrocystissp.[J].Environmental Microbiology,2011,13(6):1632-1641.
[55] OSBORN A M,MOORE E R B,TIMMIS K N.An evaluation of terminal- restriction fragment length polymorphism (T-RFLP) analysis for the study of microbial community structure and dynamics[J].Environmental Microbiology,2000,2(1):39-50.
[56] 李獻梅,王小芬,崔宗均.末端限制性片段長度多態(tài)性技術(shù)(T-RFLP)在微生物群體分析上的應(yīng)用與技術(shù)優(yōu)化[J].中國農(nóng)業(yè)大學(xué)學(xué)報,2009,14(4):1-9.
Seasonal Variation of Bacterioplankton Community Structure and Its Relationship with Environmental Factors of Mochou Lake and Zixia Lake in Nanjing
CAO Xin-yi1,2,XU Hui-min1,2,WANG Si-chen1,2,HUANG Rui1,2,SHEN Feng1,2,HE Xiao-wei1,2,YU Zhong-bo1,2,ZHAO Da-yong1,2
(1.StateKeyLaboratoryofHydrology-WaterResourcesandHydraulicEngineering,HohaiUniversity,Nanjing210098,China;2.CollegeofHydrologyandWaterResources,HohaiUniversity,Nanjing210098,China)
Seasonal variation of bacterioplankton community structure and its relationship with environmental factors of Mochou Lake and Zixia Lake in Nanjing were studied by 16S rRNA terminal restriction fragment length polymorphism(T-RFLP) technology and multivariate statistical methods.The results showed that,alpha-diversity indexes of bacterioplankton community from Mochou Lake and Zixia Lake were the highest in summer and autumn,and the lowest in spring and winter.The alpha-diversity index of bacterioplankton community from Zixia Lake was significantly greater than that from Mochou Lake in winter.Non-metric multidimensional scaling(NMDS) analysis showed that,bacterioplankton community structure of Mochou Lake and Zixia Lake had a seasonal variation.Mantel test,canonical correlation analysis(CCA) showed the most significant environmental factors which affected the bacterioplankton community structure of Mochou Lake and Zixia Lake were temperature and pH value.Effects of nutrient on bacterioplankton community structure of Mochou Lake and Zixia Lake were depended on the nutrient levels in lake.Overall,this study provides a help to research difference of bacterioplankton community structure in various eutrophic waters.
Mochou Lake;Zixia Lake;bacterioplankton;community structure;seasonal variation;environme-ntal factor;multivariate statistics;terminal restriction fragment length polymorphism(T-RFLP) technology
國家自然科學(xué)基金資助項目(41371098),水文水資源與水利工程科學(xué)國家重點實驗室專項研究經(jīng)費資助項目(20145027312,20155019012),中央高校基本科研業(yè)務(wù)費資助項目(2015B14214)
2016-07-07
曹新益(1993-),男,江蘇邳州人,碩士研究生,研究方向:水生態(tài)環(huán)境保護與修復(fù),E-mail:76565886@qq.com;通訊作者:趙大勇,教授,E-mail:dyzhao@hhu.edu.cn。
10.3969/j.issn.1672-5425.2016.12.004
曹新益,徐慧敏,王司辰,等.南京莫愁湖與紫霞湖浮游細(xì)菌群落結(jié)構(gòu)的季節(jié)性變化及其與環(huán)境因子的關(guān)系[J].化學(xué)與生物工程,2016,33(12):19-26,30.
Q 178.1 X 172
A
1672-5425(2016)12-0019-08
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
哲學(xué)評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
模具制造(2019年3期)2019-06-06 02:10:54
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
現(xiàn)代企業(yè)(2015年9期)2015-02-28 18:56:50
鄭州大學(xué)學(xué)報(醫(yī)學(xué)版)(2015年2期)2015-02-27 14:50:46
