鎘對不同發育階段中華稻蝗抗氧化性的影響
2017-01-05 10:59:52張曉明李麗君馬恩波張建珍郭亞平
山西農業科學 2016年2期
關鍵詞:差異
張曉明,李麗君,馬恩波,張建珍,郭亞平,3
(1.山西省農業科學院農業環境與資源研究所,山西太原030031;2.山西大學生物應用學研究所,山西太原030006;3.山西大學生命科學學院,山西太原030006)
鎘對不同發育階段中華稻蝗抗氧化性的影響
張曉明1,李麗君1,馬恩波2,張建珍2,郭亞平2,3
(1.山西省農業科學院農業環境與資源研究所,山西太原030031;2.山西大學生物應用學研究所,山西太原030006;3.山西大學生命科學學院,山西太原030006)
采用急性染毒方法研究Cd2+脅迫對不同發育階段中華稻蝗胸部、腹部谷胱甘肽過氧化物酶(GPx)活性、谷胱甘肽硫轉移酶(GST)活性和總抗氧化能力(T-AOC)的影響。結果表明,不同質量濃度的Cd2+作用于不同發育階段中華稻蝗體內時,其胸部和腹部的GPx,GST活性及T-AOC對Cd2+脅迫的應激反應程度不同。說明Cd2+可影響中華稻蝗的GPx,GST活性及T-AOC,中華稻蝗對Cd2+有一定的耐受性,且這種耐受性與中華稻蝗的齡期及性別有關。中華稻蝗在受到Cd2+脅迫時將調動其體內的抗氧化機制,維持H2O2產生與清除的平衡,有效地清除過量的ROS,保護機體免受外界不利因素的損害,進而避免機體DNA、蛋白質等功能的損傷。中華稻蝗可作為農業環境中Cd2+的指示生物。
中華稻蝗;鎘;總抗氧化能力;谷胱甘肽過氧化物酶;谷胱甘肽硫轉移酶
鎘(Cd)屬于毒性較強的重金屬元素之一,是生物體非必需元素。隨著經濟的飛速發展,鎘由工農業生產等途徑帶入農田環境,富集于作物體內,通過食物鏈危及人類健康[1-4]。據文獻報道,Cd可參與生物體內的氧化還原反應,產生大量的活性氧自由基(ROS),導致機體內DNA斷裂、脂質過氧化、酶失活等系列氧化應激。生物體為了減輕和防止ROS損傷,在長期的進化中,形成一套完整的抗氧化系統。
谷胱甘肽系統是生物體內主要的抗氧化防御體系,它包括還原型谷胱甘肽(GSH)、谷胱甘肽過氧化物酶(GPx)和谷胱甘肽硫轉移酶(GST)等。GPx在抗氧化系統中起著維持ROS產生與清除平衡的重要作用[5]。GPx具有清除生物體內H2O2的作用,在抗氧化機制中起著至關重要的作用[6]。GST是生物體內的重要解毒酶,可保護DNA及一些蛋白質免受不利因素的損傷。GST活性因外界不利因素如:化學污染物、脂質代謝和某些病理原因被誘導而發生變化[7]。抗氧化系統中各成分單獨或協同作用,降低ROS濃度,起到抗氧化的作用[6]。總抗氧化能力(T-AOC)表示生物體內酶促和非酶促2個體系抗氧化的總體水平,起著制約和清除機體過多ROS、保護細胞正常功能、維持機體正常代謝的作用,反映機體脂質過氧化水平以及ROS對機體損傷程度[8-10]。目前對未染毒、慢性染毒的中華稻蝗體內GPx活性、T-AOC有少量報道,但Cd2+急性脅迫對中華稻蝗不同發育階段GPx,GST活性及T-AOC的影響研究未見報道。
本試驗以中華稻蝗為材料,研究Cd2+對不同齡期中華稻蝗腹部、胸部GPx,GST活性及T-AOC的影響,探討不同齡期中華稻蝗對Cd2+脅迫的反應機制,豐富生態毒理學內容,旨在為研究昆蟲抗重金屬污染的適應機制提供基礎資料,為環境重金屬污染治理、提高環境安全性提供參考資料。
1 材料和方法
1.1 供試昆蟲
試驗用中華稻蝗于2008年7—8月采自山西省太原市(E112°36′,N37°46′)晉陽湖附近的廢棄農田中。用網捕法采集中華稻蝗后,將其帶回實驗室,在室溫條件下飼喂1~2 d。按照齡期,將大小均勻一致的雌、雄中華稻蝗各30頭分籠喂養。
1.2 急性染毒試驗
將CdCl2·2.5H2O溶于三蒸水中配制50,100,150,200 mg/L梯度的溶液,對照為三蒸水。每個處理3次重復。急性染毒、蟲體凍存方法參考文獻[11]進行。
1.3 酶液制取
測定酶活性時,將中華稻蝗從-80℃冰箱中取出,用眼科手術剪刀迅速將冰凍蟲體的胸部及腹部剪下,放入勻漿液中,制備所需酶液。酶液制備按照南京建成生物工程研究所提供的試劑盒方法。
1.4 生化分析
按照南京建成生物工程研究所提供的分光光度法在酶標儀(Spectra MAX 190,美國)上測定GPx,GST活性及T-AOC、蛋白含量。每個處理組的GPx,GST活性及T-AOC、蛋白質含量均重復測定3次。
1.5 數據分析
利用SPSS 11.5[12]軟件系統,對中華稻蝗胸部、腹部GPx,GST活性及T-AOC數據分別進行方差分析與Duncan多重比較。
2 結果與分析
2.1 中華稻蝗GPx活性對Cd2+脅迫的應答
2.1.1 中華稻蝗胸部GPx活性對Cd2+脅迫的應答從圖1-A可以看出,隨著Cd2+質量濃度的不斷提高,雌性中華稻蝗3,5齡若蟲及成蟲GPx活性均表現為先升高后降低的趨勢;4齡若蟲表現為升高趨勢。當Cd2+為150 mg/L時,3齡若蟲的GPx活性達到最高。當Cd2+為200 mg/L時,4齡若蟲的GPx活性達到最高,與對照間差異顯著。當Cd2+為150 mg/L時,5齡若蟲GPx活性顯著高于對照。ANOVA顯示,各齡期不同處理間GPx活性差異達極顯著水平(P<0.01)。

從圖1-B可以看出,當Cd2+作用于雄性中華稻蝗時,隨著Cd2+質量濃度的不斷提高,3,4齡若蟲GPx活性均表現為先升高后降低的趨勢。當Cd2+為150 mg/L時,3,4齡若蟲的GPx活性均達到最高。對于5齡若蟲及成蟲,隨著Cd2+質量濃度的提高,GPx活性整體上表現為升高趨勢,且當Cd2+質量濃度為150,200 mg/L時,GPx活性顯著高于對照。ANOVA顯示,各齡期不同處理間GPx活性差異極顯著(P<0.01)。
2.1.2 中華稻蝗腹部GPx活性對Cd2+脅迫的應答
從圖2-A可以看出,當Cd2+作用于雌性中華稻蝗時,隨著Cd2+質量濃度的不斷提高,3,4齡若蟲GPx活性表現為先升高后降低的趨勢;5齡若蟲及成蟲GPx活性表現為先降低后升高的趨勢。當Cd2+為150 mg/L時,3齡若蟲的GPx活性達到最高,與對照間差異顯著。當Cd2+為100 mg/L時,4齡若蟲的GPx活性達到最高,但與對照間差異不顯著。當Cd2+為150,200 mg/L時,5齡若蟲GPx活性顯著高于對照。ANOVA顯示,3齡若蟲及成蟲不同處理間GPx活性差異顯著(P<0.05),4,5齡若蟲不同處理間GPx活性差異極顯著(P<0.01)。
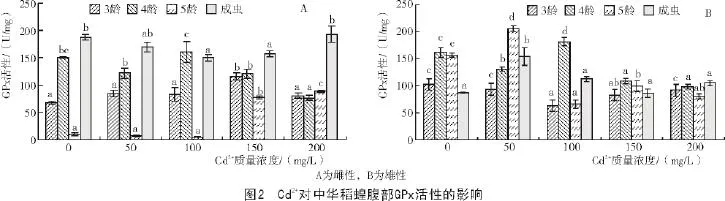
從圖2-B可以看出,當Cd2+作用于雄性中華稻蝗時,隨著Cd2+質量濃度的不斷提高,3齡若蟲GPx活性均低于對照,4,5齡若蟲及成蟲GPx活性整體上均表現為先升高后降低的趨勢。對于3齡若蟲來講,對照的GPx活性最高,蟲體的GPx活性受到抑制;當Cd2+為100 mg/L時,4齡若蟲的GPx活性達到最高,與對照間差異顯著。當Cd2+為50 mg/L時,5齡若蟲及成蟲的GPx活性達到最高,且顯著高于對照。ANOVA顯示,3齡若蟲不同處理間GPx活性差異顯著(P<0.05),4,5齡若蟲及成蟲不同處理間GPx活性差異極顯著(P<0.01)。
2.2 中華稻蝗GST活性對Cd2+脅迫的應答
2.2.1 中華稻蝗胸部GST活性對Cd2+脅迫的應答
從圖3-A可以看出,當Cd2+作用于雌性中華稻蝗時,隨著Cd2+質量濃度的不斷提高,3齡若蟲GST活性均表現為先升高后降低的趨勢,4,5齡若蟲GST活性均表現為降低趨勢,成蟲表現為升高趨勢。當Cd2+為100 mg/L時,3齡若蟲的GST活性達到最高;且當Cd2+作用于3齡中華稻蝗時,對GST活性表現為抑制作用。當Cd2+為150,200 mg/L時,4齡若蟲的GST活性與對照間差異不顯著,5齡若蟲各處理的GST活性顯著低于對照。對于成蟲,隨著Cd2+質量濃度的提高,GST活性逐漸提高。Cd2+≥100 mg/L時,5齡若蟲的GST活性顯著低于對照。ANOVA顯示,3,5齡若蟲及成蟲不同處理間GST活性差異極顯著(P<0.01),4齡若蟲不同處理間GST活性差異顯著(P<0.05)。
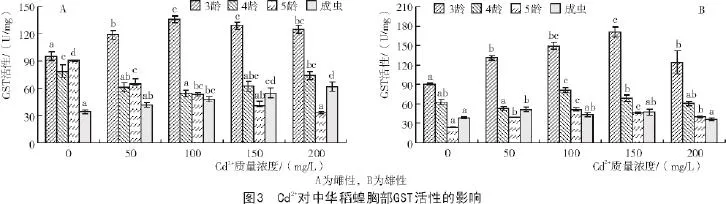
從圖3-B可以看出,隨著Cd2+質量濃度的不斷提高,雄性各齡期GST活性均表現為先升高后降低的趨勢。當Cd2+為150 mg/L時,3齡若蟲的GST活性達到最高。當Cd2+為100 mg/L時,4,5齡若蟲的GST活性達到最高,但與對照間差異顯著。當Cd2+為50 mg/L時,成蟲的GST活性達到最高,且顯著高于對照。ANOVA顯示,各齡期若蟲不同處理間GST活性差異極顯著(P<0.01),成蟲不同處理間GST活性差異顯著(P<0.05)。
2.2.2 中華稻蝗腹部GST活性對Cd2+脅迫的應答
從圖4-A可以看出,隨著Cd2+質量濃度的不斷提高,雌性3齡若蟲GST活性表現為先升高后降低的趨勢,4齡若蟲表現為降低趨勢,5齡若蟲及成蟲GST活性均表現為先降低后升高的趨勢。當Cd2+為 100 mg/L時,3齡若蟲的GST活性達到最高;當Cd2+為0 mg/L時,4齡若蟲與成蟲的GST活性達到最高。當Cd2+為150 mg/L時,5齡若蟲的GST活性達到最高。ANOVA顯示,各齡期不同處理間GST活性差異極顯著(P<0.01)。

從圖4-B可以看出,當Cd2+作用于雄性中華稻蝗時,隨著Cd2+質量濃度的不斷提高,3齡若蟲及成蟲的GST活性均表現為先降低后升高趨勢,4,5齡若蟲GST活性均表現為先升高后降低趨勢。當Cd2+為150 mg/L時,3齡若蟲的GST活性達到最高。當Cd2+為100 mg/L時,4,5齡若蟲及成蟲的GST活性均達到最高,4,5齡若蟲的GST活性與對照間差異顯著,而成蟲的GST活性與對照間差異不顯著。ANOVA顯示,各齡若蟲不同處理間GST活性差異極顯著(P<0.01),成蟲不同處理間GST活性差異顯著(P<0.05)。
2.3 中華稻蝗T-AOC對Cd2+脅迫的應答
2.3.1 中華稻蝗胸部T-AOC對Cd2+脅迫的應答
從圖5-A可以看出,隨著Cd2+質量濃度的不斷提高,雌性3,4,5齡若蟲T-AOC均表現為先升高后降低的趨勢。當Cd2+為150 mg/L時,3齡若蟲的T-AOC達到最高。當Cd2+為50 mg/L時,4齡若蟲的T-AOC達到最高,但與對照間差異不顯著。當Cd2+為100 mg/L時,5齡若蟲T-AOC顯著高于對照。隨著Cd2+質量濃度的提高,成蟲的T-AOC逐漸提高。當Cd2+質量濃度為150,200 mg/L時,成蟲T-AOC顯著高于對照。ANOVA顯示,3齡若蟲及成蟲不同處理之間T-AOC差異極顯著(P<0.01),4,5齡若蟲不同處理之間T-AOC差異顯著(P<0.05)。

對于雄性中華稻蝗(圖5-B)來講,隨著Cd2+脅迫濃度的不斷提高,3齡若蟲T-AOC逐漸增加,當Cd2+為50 mg/L時,3齡若蟲T-AOC達到最高,為2.91 U/mg,隨后3齡若蟲的T-AOC表現出降低趨勢。當Cd2+為50,100 mg/L時,3齡若蟲的T-AOC顯著高于對照。隨著Cd2+質量濃度的提高,4,5齡若蟲T-AOC表現為升高趨勢。當Cd2+為200 mg/L時,4齡若蟲T-AOC達到最高,且顯著高于對照。當Cd2+質量濃度為150,200 mg/L時,5齡若蟲T-AOC顯著高于對照。當Cd2+為50 mg/L時,成蟲T-AOC顯著低于對照,其余Cd2+質量濃度對T-AOC未產生顯著影響。ANOVA顯示,3,4,5齡若蟲不同處理間T-AOC差異顯著(P<0.05),成蟲不同處理間T-AOC差異極顯著(P<0.01)。
2.3.2 中華稻蝗腹部T-AOC對Cd2+脅迫的應答
從圖6-A可以看出,雌性中華稻蝗受到Cd2+脅迫后,隨著Cd2+質量濃度的不斷提高,3,4齡若蟲及成蟲的T-AOC逐漸增加,隨后則表現出降低趨勢。當Cd2+為100 mg/L時,3,4齡若蟲T-AOC達到最高,且顯著高于對照。當Cd2+為50,200 mg/L時,成蟲T-AOC顯著高于對照。當5齡若蟲受到Cd2+脅迫時,各處理的T-AOC均低于對照。ANOVA顯示,3齡若蟲不同處理間T-AOC差異極顯著(P<0.01),4,5齡若蟲、成蟲不同處理間T-AOC差異顯著(P<0.05)。

從圖6-B可以看出,當Cd2+作用于不同齡期的雄性中華稻蝗時,T-AOC變化趨勢相同,均為隨著Cd2+質量濃度的不斷提高,T-AOC逐漸增加,而后表現出降低趨勢。對于3齡若蟲來講,當Cd2+為50,100 mg/L時,T-AOC顯著高于對照,且Cd2+為50 mg/L時T-AOC達到最高。當Cd2+為100 mg/L時,4齡若蟲的T-AOC達到最高,但與對照間差異不顯著。當Cd2+為100,150mg/L時,5齡若蟲T-AOC顯著高于對照。當Cd2+為50,100,150 mg/L時,成蟲的T-AOC均顯著高于對照,而Cd2+為200 mg/L的T-AOC顯著低于對照。ANOVA顯示,3,4齡若蟲、成蟲不同處理間T-AOC差異極顯著(P<0.01),5齡若蟲不同處理間T-AOC差異顯著(P<0.05)。
3 討論
中華稻蝗胸部是其運動中心,由大量肌肉和外骨骼組成,內部臟器包括前腸的食道部分、唾液腺及前胸腺等內分泌腺體;腹部是蟲體的代謝中心,大部分腸道、馬氏管、生殖腺、血淋巴和脂肪體等大量臟器存在其中,是重金屬累積的主要器官[13]。本研究結果顯示,Cd2+的急性脅迫對蟲體胸部及腹部產生不同程度的氧化脅迫。
生物體內存在完善的抗氧化系統,起著調節活性氧自由基(ROS)生成與清除的動態作用。在正常情況下,機體內ROS的產生、利用和清除保持動態平衡,ROS維持在較低水平。當環境中存在污染脅迫時,機體內的氧化還原反應將產生大量的ROS[14]。當ROS在機體內的累積超過抗氧化系統的清除能力時,則會產生氧化損傷,使機體處于氧化應激狀態[15]。谷胱甘肽系統是生物體主要的抗氧化防御體系,在維持細胞內環境穩態及機體的解毒代謝中起重要作用[16-17]。T-AOC是近年研究發現的反映機體抗氧化酶系統和非酶系統對外來刺激的代償能力以及機體自由基代謝的狀態綜合性指標,是機體頡頏ROS的主要體系,起著制約和清除機體產生過多ROS、保護細胞正常功能、維持機體正常代謝的作用[15]。
本研究結果表明,當不同質量濃度Cd2+作用于中華稻蝗時,不同齡期蟲體的GPx,GST活性及T-AOC均發生不同變化,因此,可將中華稻蝗作為農業環境中Cd2+的指示生物。同時,表明Cd2+作用導致蟲體內ROS濃度提高,蟲體則啟動其抗氧化系統清除過量的ROS,保護機體免受其損害,而T-AOC的變化是機體內各抗氧化酶活性改變共同作用的結果[18],這一結果與李麗君等[11,19]報道的急性Cd2+染毒可影響蟲體抗氧化酶活性一致。
本研究還表明,在Cd2+的作用下,不同齡期、性別蟲體的GPx,GST活性及T-AOC發生不同變化。隨著Cd2+質量濃度的提高,三者分別表現為先升高后降低的趨勢。表明低質量濃度Cd2+的存在激活了中華稻蝗體內GPx,GST的合成途徑,T-AOC隨之提高,GPx,GST可起到維持H2O2產生與清除平衡的作用,降低Cd2+毒性的作用,達到保護中華稻蝗的目的[20];但高質量濃度Cd2+導致GPx,GST活性呈下降趨勢,T-AOC受到抑制,表明中華稻蝗抗氧化系統對Cd2+的耐受性有一定的極限值。超過這一極限值時,隨著Cd2+質量濃度繼續提高,一些未被清除的自由基對細胞產生不可逆的傷害,導致酶活性逐漸降低,GST的解毒作用被抑制[5,7,21-22]。
隨著Cd2+質量濃度的提高,三者分別表現為升高趨勢,在Cd2+的脅迫下,中華稻蝗體內發生一系列氧化還原反應,產生大量的ROS,表現為GPx,GST活性及T-AOC的提高,提高中華稻蝗機體抗氧化能力,起到積極清除過量ROS的作用,使ROS對中華稻蝗的危害降到最小,保證機體進行正常的代謝活動[5,7,14]。
隨著Cd2+質量濃度的提高,三者分別表現為先降低后升高趨勢,表明低質量濃度Cd2+作用于中華稻蝗時,機體內未產生大量的ROS,Cd2+未對機體產生毒害作用;高質量濃度Cd2+可導致ROS濃度提高,因此,只有抗氧化酶系統和非酶系統中不同成分協同作用,提高T-AOC,使過量ROS的清除遠遠超出產生速度,進而降低機體的脂質過氧化水平以及ROS對機體的損害程度[10,14]。Cd2+的存在導致GPx,GST活性及T-AOC降低,表明低質量濃度的Cd2+即可對中華稻蝗GPx,GST合成途徑產生毒害作用,致使細胞內自由基累積,造成基因損傷,導致細胞凋亡和功能喪失[5,23],致使T-AOC表現為下降趨勢,Cd2+對機體造成一系列的損害[15]。
4 結論
不同質量濃度的Cd2+作用于不同發育階段中華稻蝗體內時,其胸部和腹部的GPx,GST活性及T-AOC對Cd2+脅迫的表現應激反應程度不同,中華稻蝗可作為農業環境中Cd2+的指示生物。中華稻蝗在受到Cd2+脅迫時將調動其體內的抗氧化機制,維持H2O2產生與清除的平衡,有效地清除過量的ROS,保護機體免受外界不利因素的損害,進而避免機體DNA、蛋白質等功能的損傷。
[1]何俊瑜,任艷芳,朱誠期,等.鎘脅迫對鎘敏感水稻突變體活性氧代謝及抗氧化酶活性的影響[J].生態環境,2008,17(3):1004-1008.
[2]侯青葉,楊忠芳,楊曉燕,等.成都平原區水稻土成土剖面Cd形態分布特征及影響因素研究[J].地學前緣,2008,15(5):37-46.
[3]王新,周啟星.土壤重金屬污染生態過程、效應及修復[J].生態科學,2004,23(3):278-281.
[4]顧繼光,周啟星,王新.土壤重金屬污染的治理途徑及其研究進展[J].應用基礎與工程科學學報,2003,11(2):143-151.
[5]劉娜,閆博,李涌泉,等.Cd2+對長江華溪蟹谷胱甘肽系統的影響[J].環境科學,2008,29(8):2302-2307.
[6]李麗君,郭亞平,劉平,等.鎘長期暴露對中華稻蝗抗氧化機制的影響[J].農業環境科學學報,2011,30(3):443-448.
[7]田志環,申保忠.廢舊干電池污染液對泥鰍谷胱甘肽硫轉移酶活性的影響[J].濱州學院學報,2007,23(3):47-50.
[8]劉汝祥,侯明海,李彥芹,等.不同維生素A水平對荷斯坦種公牛血液抗氧化指標的影響[J].西南農業學報,2008,21(3):798-801.
[9]馮望寶,王安,艾濤.不同鋅水平對籠養育成蛋鴨生長性能及總抗氧化能力的影響[J].東北農業大學學報,2007,38(5):654-659.
[10]吳偉,聶鳳琴,瞿建宏,等.2,2′,4,4′-四溴聯苯醚對鯽魚離體肝臟組織的氧化脅迫[J].農業環境科學學報,2009,28(5):1005-1009.
[11]Li L,Liu X,GuoY,et al.Activity of the enzymes of the antioxidative system in cadmium-treated Oxya chinensis(Orthoptera Acridoidae)[J].Environmental Toxicology and Pharmacology,2005,20:412-416.
[12]洪楠,林愛花,李志輝,等.統計分析教程[M].北京:電子工業出版社,2000:134-148.
[13]孫鴿,吳海花,席玉英,等.長期取食染鎘小麥后中華稻蝗體內鎘的累積分布[J].農業環境科學學報,2009,28(9):1812-1817.
[14]孟凡平,王偉,李瑩,等.苯并芘和VE對紅鰭東方鲀抗氧化酶影響研究[J].水產科學,2012,31(5):270-274.
[15]文靜,林志偉,周緒霞,等.枯草芽孢桿菌B10對飼喂高脂日糧小鼠抗氧化功能的影響[J].中國獸醫學報,2012,32(1):111-114.
[16]廖一驍,胡振英,程富勝,等.酵母硒對肉雞血清中GSH-Px、GSTs活性的影響[J].湖北農業科學,2010,49(8):1918-1920.
[17]馬恩波,葛娉婷,張建琴,等.飛蝗兩種群羧酸酯酶及谷胱甘肽S-轉移酶活性與馬拉硫磷敏感性關系的研究[J].山西大學學報:自然科學版,2012,35(2):383-387.
[18]高春生,王春秀,張書松.水體銅對黃河鯉肝胰臟抗氧化酶活性和總抗氧化能力的影響[J].農業環境科學學報,2008,27(3):1157-1162.
[19]李麗君,郭亞平,武文麗,等.鎘對中華稻蝗保護酶系統的毒性效應[J].農業環境科學學報,2010,29(8):1448-1454.
[20]韓雨哲,姜志強,任同軍,等.氧化魚油與棕櫚油對花鱸肝臟抗氧化酶及組織結構的影響[J].中國水產科學,2010,17(4):798-806.
[21]張彬彬.乙草胺對泥鰍肝臟乙酰膽堿酯酶和谷胱甘肽硫轉移酶的影響[J].淡水漁業,2008,38(4):73-75.
[22]王蕓,李健,李吉濤,等.pH脅迫對中國明對蝦抗氧化系統酶活力及基因表達的影響[J].中國水產科學,2011,18(3):556-564.
[23]王麗麗,夏斌,陳碧鵑,等.鎘脅迫對蝦夷扇貝抗氧化防御系統的影響[J].海洋環境科學,2012,31(1):39-42.
Effects of Cd2+on Anti-oxidative Capacity in Different Developing Stage of Oxya chinensis(Orthoptera Acridoidae)
ZHANG Xiao-ming1,LI Li-jun1,MA En-bo2,ZHANG Jian-zhen2,GUO Ya-ping2,3
(1.Institute of Agricultural Environment and Resource,Shanxi Academy of Agricultural Sciences,Taiyuan 030031,China;2.Institute of
Applied Biology,Shanxi University,Taiyuan 030006,China;3.College of Life Sciences,Shanxi University,Taiyuan 030006,China)
To explore and clarify the responses of Oxya chinensis to Cd2+stress,provide the scientific evaluation of Cd2+tolerance, the toxic effect of Cd2+on activities of glutathione peroxidase(GPx)and glutathione S-transferase(GST),and the total anti-oxidative capacity(T-AOC)in different developing stage of O.chinensis injected with Cd2+were determined.Different developing stages O. chinensis were injected with Cd2+at different concentrations(0,50,100,150,200 mg/L),and they were collected after 24 h.The results showed that the effect of Cd2+on GPx and GST activities and T-AOC were different within the experimental gradient.The GPx and GST activities,and T-AOC depended on the content of oxygen species(ROS)under Cd2+stress in the experiment,and Cd2+could induce damage of antioxidant defense system in O.chinensis,but O.chinensis kept its GPx and GST activities,and T-AOC increasing from Cd2+damage in some Cd2+contents.The tolerance of Cd2+changed in different developing stages and male or female O.chinensis.Variety of GPx and GST activities,and T-AOC in different developing stages and male or female O.chinensis might act as indicative functions for Cd2+contamination.
Oxya chinensis;cadmium(Cd);total anti-oxidative capacity(T-AOC);glutathione S-transferase(GST);glutathione peroxidase(GPx)
X174
A
1002-2481(2016)02-0152-06
10.3969/j.issn.1002-2481.2016.02.06
2015-08-28
國家自然科學基金項目(31071980);高等學校博士點基金項目(20111401110006);山西省回國留學人員科研資助項目(2008114);山西省農業科學院博士基金項目(YBSJJ0702)
張曉明(1958-),男,山西沁縣人,助理研究員,主要從事昆蟲毒理學研究工作。郭亞平為通信作者。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
