植物木質素的合成與調控研究進展
2017-01-05 06:13:48丁霄曹彩榮李朋波吳翠翠曹美蓮楊六六
山西農業科學 2016年9期
關鍵詞:植物
丁霄,曹彩榮,李朋波,吳翠翠,曹美蓮,楊六六
(山西省農業科學院棉花研究所,棉花種質資源利用與分子設計育種山西省重點實驗室,山西運城044000)
植物木質素的合成與調控研究進展
丁霄,曹彩榮,李朋波,吳翠翠,曹美蓮,楊六六
(山西省農業科學院棉花研究所,棉花種質資源利用與分子設計育種山西省重點實驗室,山西運城044000)
木質素作為植物次生細胞壁的重要組分,分布于輸導組織和木質化組織細胞壁中,不僅具有提高細胞壁的隔水性和機械強度,而且在提高植物的抗病抗逆方面也發揮著重要作用。對植物木質素的種類、合成調控和利用基因工程從源頭調控植物木質素含量等方面的研究現狀進行了概述;隨著轉基因技術的發展,有望通過更多更有效的途徑來改變植物木質素的組成。
木質素;合成與調控;基因工程
木質素是植物經類苯丙酸途徑合成的單體經進一步分化形成的3個苯丙烷衍生物通過化學鍵聚合而成的高分子化合物[1]。其大量存在于植物的木質部纖維素纖維之間,通過形成交織網來硬化細胞壁,能夠使木質部維持極高的硬度,以形成植株的形態,承載整株植物的重量,并形成較強的抗壓能力,因此,木質素在維管植物的進化中發揮著重要作用[2]。在木本植物中,木質素占25%~30%;在草本植物中,木質素占16%;在自然界有機物中的含量僅次于纖維素,是植物轉化太陽能形成的重要有機物之一[3]。此外,木質素還密切參與植物的生長過程,如水分運輸、抗病及對逆境的適應能力[4]。
1 木質素結構的單體類型
組成木質素的單體類型主要有香豆醇、松柏醇和芥子醇3種木質醇,這3種木質醇是苯丙烷衍生物經不同的羥基化和甲基化進而形成的并通過不同化學鍵連接而成[5]。3種木質醇香豆醇、松柏醇與芥子醇進一步形成香豆醇殘基型(p-hydroxyphenyl H)木質素、松柏醇殘基型(guaiacyl G)木質素和芥子醇殘基型(syringyl S)木質素(圖1),形成的木質素殘基通過共價方式如醚鍵(-O-)與碳-碳鍵(C-C)2種方式連接起來[6]。
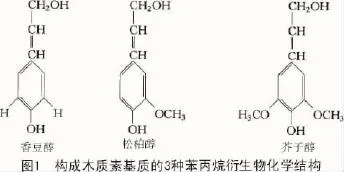
不同的植物類群中,木質素所占的比例及其3種不同類型成分的百分比也不盡相同。G型木質素主要由松柏醇聚合而成,該類植物存在于蕨類植物與裸子植物;G-S型木質素主要由松柏醇與芥子醇聚合而成,該類植物存在于雙子葉植物;H-G-S型木質素不同程度地包含了3種木質醇單體,該類植物主要是單子葉植物[7]。不僅不同物種間木質素的組成有差異,同一物種在不同時期不同部位其木質素的組成也是不同的,在同一物種間木質素的差異往往需要借助細胞水平上的檢測手段如組織細胞化學染色、紫外光譜學及利用細胞壁免疫抗體等手段進行檢測分析和相對測定[8]。除了上述常見的3種木質素單體外,自然界中還存在少量其他類型的木質素單體參與木質素的合成,如自由基偶聯產生的阿魏酸脫氫二聚體及乙酰化的木質素單體[9]。
2 木質素的功能
作為植物次生細胞壁的重要組成成分之一,木質素在植物中具有抗倒伏、保證水分養分輸送的功能。木質素滲入到由纖維素、半纖維素等大分子構成的細胞壁骨架中并與之互相結合,從而增加了細胞機械強度,進而提高莖稈組織的抗倒伏能力[10]。通過抑制木質素合成途徑相關的基因,發現在雙子葉和單子葉植物細胞中的木質素含量均出現下降,伴隨木質素含量的下降,植物細胞壁的抗壓強度下降并出現細胞壁坍塌現象[11]。胡丹等[12]測定不同抗倒性甜蕎木質素含量發現,在不同時期木質素含量的多少與甜蕎的倒伏性狀均呈顯著相關。木質素作為高分子聚合物,其所具有的不溶于水的特性使得構成植物維管部的細胞壁具有疏水性,保證了植物中的水分、溶解其中的礦物質及有機物等能夠通過植物維管系統實現長距離輸送[13]。
木質素與植物的抗病性具有密切的關系[14]。木質素技揳入到由纖維素等物質構成的細胞壁骨架中形成物理屏障,進一步降低了病原菌及其多種變種的入侵,在細胞結構水平上提高了植物對病原菌及害蟲的防御能力[14-16]。吳立柱等[17]研究黃萎病對棉花導管的影響發現,抗病品種導管中木質素含量相較于感病品種高,因此抗病棉花品種在接種黃萎病菌后,輸送水分養分的導管未發生明顯堵塞。許勇等[18]對西瓜接種枯萎病病菌,發現在抗病西瓜品種中枯萎病病菌誘導西瓜的組織結構抗性,結果顯示,木質素在根莖的維管束病害抗性中具有重要地位。毛愛軍等[19]通過水楊酸處理研究辣椒抗疫病的生化機制,結果顯示,木質素含量與抗病性顯著相關,其中,相同辣椒品種經水楊酸處理相較于對照,木質素含量升高,抗病性增強。方東鵬等[20]對大豆的研究表明,在正常情況下,不同抗性的野生大豆品種木質素的含量差異不顯著,但分別接種大豆疫霉菌后,感病初期(12 h)抗病性強的野生大豆品種比感病品種木質素含量平均增加50%。在丹參的木質素合成研究中發現,接種病原菌后丹參木質素合成基因SmCCR2隨著時間的推移呈周期性波動,說明木質素參與到丹參的抗病過程[21]。
木質素參與植物對低溫脅迫的響應。研究發現,鴨兒芹經低溫處理后,木質素合成基因CjHCT顯著下降,木質素含量減少[22],同時對木質素合成基因CjC4H研究發現,該基因高溫條件下響應迅速,低溫條件下表達推遲,并且低溫條件下鴨兒芹表現出更加脆嫩的商品價值[23];通過1-MCP抑制枇杷果實中木質素的合成可有效減緩枇杷果實的冷害性狀[24],因此,抑制木質素的合成可以有效提高商品作物冷鏈儲藏時間。
在棉花中的研究顯示,調控木質素合成的NAC類轉錄因子GhNAC12,GhNAC20明顯受到低溫誘導[25];對比不同抗凍能力的楊樹品種,抗凍能力強的楊樹品種在低溫條件下木質素含量提升顯著,而較弱的楊樹品種在低溫條件下木質素含量差異不顯著[26],因此,通過提高木質素含量進而減少植物在低溫條件下電解液的滲出,或是通過提高木質素前體物質的合成,減少氧化物對植物的毒害來應對低溫脅迫都存在可能。
3 木質素合成過程的關鍵基因
前人的研究結果顯示,莽草酸途徑、類苯丙酸途徑和特異途徑是生物體中合成木質素的必經過程。第1步是植物通過光合作用形成的葡萄糖轉化為酪氨酸與色氨酸等芳香族氨基酸即莽草酸途徑[27];第2步將第一步所形成的化合物經過脫氨基、羥基化與甲基化等步驟,將芳香族氨基酸轉化為羥基肉桂酸類化合物(Hydroxycinnamic acids,HCAs)及羥基肉桂酸酯酞輔酶A類化合物(Hydroxycinnamic acid Coenzyme A,HCA-CoA)即類苯丙酸途徑;最后一步是將第2步形成的HCAs及HCA-CoA還原為木質醇單體即木質素合成的特異途徑,形成的單體再通過氧化聚合作用形成木質素[28]。在合成木質素的過程中,第2步和第3步起主要作用,因此,類苯丙酸途徑和特異途徑成為研究木質素合成的關鍵所在[29](圖2)。
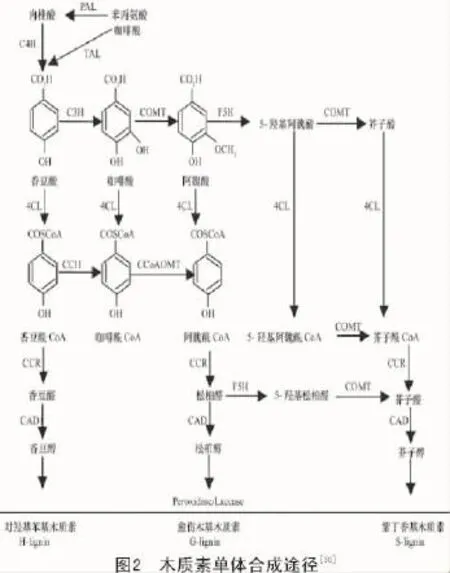
最近幾年的研究表明,木質素合成的總量與苯丙氨酸解氨酶(phenylalanine ammonialyase,PAL)、肉桂酸-4-羥基化酶(cinnamate-4-hydroxylase,C4H)和4-香豆酸輔酶A連接酶(4-Coumarate: CoA ligase,4CL)生成量及其活性的水平密切相關[12];而木質素的特異性與阿魏酸5-羥基化酶(ferulate 5-hydroxylase,F5H)、咖啡酸/5-羥基阿魏酸O-甲基轉移酶(caffeicacid O-methyltransfenase,COMT)和咖啡酰輔酶A/5-羥基阿魏酰輔酶A O-甲基轉移酶(caffeoyl-CoA O-methyltransfenase,CCoAOMT)這3種酶緊密相關,決定了3種不同的單體在木質素結構中所占比例[31]。
3.1 苯丙氨酸解氨酶(PAL)
PAL位于類苯丙酸途徑的源頭(圖2),除了參與木質素合成途徑外,同時還參與香豆酸酯類和類黃酮等次生物質的合成,因此,PAL同時也是合成其他酚類物質的必經途徑,并非只是單一控制植物木質素的合成。盡管PAL作為木質素合成的限速步驟之一,但對PAL的抑制研究顯示,轉基因植物在木質素合成受到抑制的同時也破壞了植物對其所處逆境的應答響應機制,因此,通過基因工程改變PAL進而改善作物性狀的成果較少[32]。但通過外源激素處理手段增強PAL的表達進而提高木質素含量,最終實現竹根姜抗病性提高[33]。此外,Xu等[34]通過轉錄組測序技術發現,在侵染了黃萎病病菌后,棉花感、抗品種間PAL表達差異十分顯著,因此,通過啟動子改造不失為一種更有效的途徑。
3.2 肉桂酸-4-羥基化酶(C4H)
C4H是一類單加氧酶,在木質素合成途徑中位于PAL之后,催化肉桂酸轉化為香豆酸即木質素單體的前體,PAL和C4H被認為是苯丙氨酸轉為香豆酸的唯一途徑,但與PAL一樣,C4H除了參與木質素的代謝合成也參與植物其他酚類物質的合成。對C4H的轉基因植物研究顯示,抑制C4H并未出現植物生長異常,但改變了木質素單體S/G的值,尤其是降低了木質素單體S的含量,推測木質素單體G很可能存在其他的通路來進行補償或者C4H與其他酶形成復合酶進行木質素單體S的合成[35]。
3.34 -香豆酸輔酶A連接酶(4CL)
4CL催化不同底物如香豆酸、咖啡酸、阿魏酸和肉桂酸等的羥基苯乙酸形成相應的CoA酯,位于類苯丙烷代謝途徑中控制形成不同木質素單體類型的轉折點,是調控木質素單體合成的關鍵酶[36]。目前,已在多種植物中發現并克隆驗證的了4CL的功能,抑制4CL的表達可明顯降低轉基因植物中木質素的含量且轉基因材料表現出明顯的生長加快現象,可能與木質素含量降低生物量不變導致纖維素含量增加有關[37]。同時,轉基因植物細胞壁出現明顯的顏色變化,說明其分子結構已發生明顯的變化并呈褐色,因此,可以利用4CL基因改變莖稈木質部顏色的特征挖掘其作為植物安全篩選標記的應用潛力。
3.4 阿魏酸5-羥基化酶(F5H)
F5H為S型木質素合成的唯一環節。Marita等通過研究擬南芥f5h缺失突變體發現,其木質素主要由G型木質素組成,過表達F5H將極大地提高S型木質素的含量并提高S/G的比值,因此,F5H為合成S型木質素的關鍵限速酶之一[38]。對F5H的表達研究發現,F5H存在于植物莖稈的微粒體中,在莖稈的木質部特別是厚壁組織中活性最高[32]。
3.5 咖啡酸/5-羥基阿魏酸-O-甲基轉移酶(COMT)
COMT可以催化3類底物即咖啡酸、5-羥基松柏醛、5-羥基松柏醇甲基,其催化產物分別形成阿魏酸、芥子醛和芥子醇(圖2)。將綠竹中的COMT通過農桿菌侵染煙草反義抑制煙草中的COMT表達,轉基因煙草中出現木質素含量下降、莖稈變紅以及植株矮化現象[39]。抑制玉米中的COMT表達后玉米木質素含量尤其是S型木質素含量顯著降低,進而改善玉米的飼用價值[40]。Rastogi等[41]通過將白楊中得到的COMT抑制銀合歡中的COMT表達,發現在轉基因銀合歡中木質素含量下降并出現纖維素增加的性狀,其中木質素的減少主要是由S型木質素含量的下降造成的。通過RNAi技術干擾丹參中COMT的表達,結果顯示,轉基因丹參中木質素和S型木質素含量均出現下降[42]。植物中的COMT活性將直接影響轉基因植物中的S型木質素的合成,證明COMT通過對S型木質素前體的合成進而實現對S型木質素的控制。同時,COMT的表達也具有時空特異性,在植物木質部中大量表達,在葉中幾乎檢測不到COMT。
3.6 咖啡酰輔酶A/5-羥基阿魏酰輔酶A O-甲基轉移酶(CCoAOMT)
CCoAOMT通過控制咖啡酰CoA轉化成阿魏酰CoA來調控植物木質素的合成及類型。對CCoAOMT轉基因植物研究顯示,抑制CCoAOMT合成的同時對S型木質素和G型木質素的合成形成抑制,但對G型木質素的抑制尤為明顯,進而提高了S/G的值。Li等[43]通過RNA干擾技術抑制玉米中的CCoAOMT,結果顯示,轉基因玉米中木質素含量下降22.4%,纖維素含量提高23.3%,S/G的值提高57.08%。研究新西蘭松ccoaomt缺失突變體和RNAi干擾材料證明,CCoAOMT對木質素合成具有調控的作用[44],利用竺麻CCoAOMT構建植物干擾表達載體分別對煙草和亞麻進行轉化,轉基因煙草和亞麻木質素含量均出現下降[29]。因此,CCoAOMT主要是通過對G型木質素的合成控制進而改變木質素的含量。CCoAOMT在棉花中各個部位均有表達,以莖稈中的表達量為最高[45]。
4 調控木質素合成的轉錄因子
在木質素的合成調控網絡中存在許多對木質素合成相關基因進行調控的轉錄因子,其中主要有MYB類轉錄因子和NAC類轉錄因子。
4.1 MYB類轉錄因子
MYB類轉錄因子的特點是在其N端有一處串聯不同的MYB結構域(R1,R2,R3)并根據不同的串聯分為3個亞類。MYB結構域通過Helix-Turn-Helix(HTH)與DNA中的大溝結合。同時在其C端有一處由酸性氨基酸組成通過折疊成雙親的α-螺旋形成的轉錄激活區。
MYB類轉錄因子已在擬南芥中證明參與了植物木質素的調控,如R2R3型和LIM[46]等通過結合參與苯丙烷途徑的基因PAL,C4H,4CL,C3H,CCoAOMT,CCR和CAD啟動子區中存在的AC元件發揮調控作用,上述基因的啟動子區均含有包括AC-Ⅰ(ACCTACC),AC-Ⅱ(ACCAACC)或AC-Ⅲ(ACCTAAC)中一個或多個AC元件[47],這種轉錄因子通過與苯丙烷途徑基因的啟動子結合使得整個苯丙烷代謝受到統一的調控[48]。已有的研究結果也證明了上述結論,如在擬南芥中2個myb103突變體中,F5H的表達受到抑制,導致了轉基因擬南芥中S型木質素含量下降,G型木質素含量增加,轉錄因子MYB103通過對F5H的調控被證明是木質素合成所必需的;在myb32突變體中,COMT的表達量上升,轉錄因子MYB32通過抑制COMT的表達實現對木質素合成的調控;Omer等[49]將銀合歡中的轉錄因子LlMYB1轉入煙草中過表達,發現轉基因煙草中的木質素含量顯著降低;在楊樹的次生細胞壁形成區域轉錄因子Ptt MYB21a表達量較高,通過轉基因抑制楊樹轉錄因子PttMYB21a的生成使得轉基因材料中CCoAOMT表達量升高,證明轉錄因子PttMYB21a抑制CCoAOMT的轉錄[50];在轉基因煙草中過表達MYB308和MYB330發現,4CL1,C4H和CAD的表達量下降,進而導致轉基因煙草中木質素含量下降,證明轉錄因子MYB308和MYB330是4CL1,C4H和CAD的負調控因子[51];在石榴種皮木質素含量的研究中發現,PgMYB表達量與木質素含量及籽粒硬度呈顯著負相關,表明PgMYB對石榴中的木質素呈負調控[52]。
4.2 NAC類轉錄因子
NAC類轉錄因子在植物體中存在多種功能,對木質素的調控只是其中的一類,其結構特點是在N端有150個氨基酸組成的保守性NAC結構域,其C端是差異較大的轉錄激活區。NAC類轉錄因子通過與調控木質素合成的MYB類轉錄因子的結合進而間接對木質素生物合成進行調控,由于NAC位于木質素調控網絡的上游,因此,通過NAC調控還存在很多不確定性,目前已在模式植物擬南芥中克隆驗證了部分NAC類轉錄因子參與木質素合成的調控[53]。NST1(NAC secondary wall thickening promoting factor1,NST1)是在擬南芥中發現的參與植物木質素合成調控的NAC類轉錄因子,通過在擬南芥中過表達NST1發現轉基因材料中的木質素含量提高[54];SND1(secondary wall-associated nac domain protein,SND1)主要是控制纖維素的,通過轉基因過表達NAC類轉錄因子SND1發現其也影響木質素在細胞壁中的沉積[55]。
除了上述兩大類轉錄因子外,還有其他的轉錄因子參與植物木質素的調控。如WRKY類轉錄因子與MYB類轉錄因子相似,與NST2的啟動子區結合,通過對髓細胞中木質素的沉積進行負調控[16],實現對植物木質素的調控作用[56];bHLH(Basic helix-loop-helix)類轉錄因子在擬南芥中過表達,表現出MYB83,MYB46和MYB63等轉錄水平升高,進而降低4CL1,HCT,COMT,PAL1和CCR1的表達量,使得轉基因植株的木質素含量顯著降低,因而,SbbHLH1在調控木質素代謝中的地位大于MYB[56]。
5 展望
由于調控木質素合成途徑的很多關鍵酶如CCoAOMT,F5H,COMT以及4CL在許多植物中往往是多拷貝,利用轉基因過表達木質素調控相關基因往往由于同源抑制無法獲得提高木質素含量或改變木質素結構組成的預期效果,結果是過表達與反義核酸表現一致,因此,采用共抑制、反義抑制技術在實現對木質素合成的分子調控更有效[57]。但木質素的合成調控是一個很大的代謝網絡,具有很強的可塑性和替補性,當其合成代謝發生異常時,往往會存在其他的補償機制以保證木質素在植物體內的生理功能。隨著多個物種基因序列的公布,可以實現對木質素合成調控相關基因的精確定位,并進行序列差異性、多態性、內含子剪切及啟動子方面的分析[58-59],并利用基因組編輯技術[60]進行定向誘變,避免多倍體植物中存在的同源抑制現象,以期獲得抗病抗倒伏植物材料。
[1]程華.銀杏黃酮和木質素代謝相關基因功能分析[D].南京:南京林業大學,2012.
[2]谷振軍.赤桉木質素單體合成途徑的基因家族研究[D].長沙:中南林業科技大學,2014.
[3]曾育才,張學先,劉小玲.造紙黑液木質素及其綜合利用[J].廣東化工,2005,32(10):17-20,81.
[4]李曉榮.東方百合木質素合成相關基因的克隆及功能分析[D].南京:南京林業大學,2009.
[5]Ralph J,Mackay J J,Hatfield R D,et al.Abnormal lignin in a loblollypine mutant[J].Science,1997,277:235-239.
[6]藺占兵.小麥肉桂酰輔酶A還原酶(CCR)基因的分離和功能分析[D].北京:中國科學院研究生院(植物研究所),2003.
[7]Faix O.Classification of lignins from different botanical origins by ft-ir spectroscopy[J].Holzforschung-international Journal of the Biology,Chemistry,Physics and Technology of Wood,1991,45(S1):21-28.
[8]石堅.OsSHN1和OsMYB48參與水稻木質素合成調控的研究[D].武漢:華中農業大學,2014.
[9]趙華燕.利用反義RNA技術進行木質素生物合成調控的研究[D].北京:中國科學院研究生院(植物研究所),2004.
[10]陳曉光,史春余,尹燕枰,等.小麥莖稈木質素代謝及其與抗倒性的關系[J].作物學報,2011,37(9):1616-1622.
[11]Li X,Chapple C.Understanding lignification:challenges beyond monolignol biosynthesis[J].Plant Physiology,2010,154(2):449-452.
[12]胡丹,劉星貝,汪燦,等.不同抗倒性甜蕎莖稈木質素合成關鍵酶基因的表達分析[J].中國農業科學,2015,48(9):1864-1872.
[13]師竹娟.甘藍型油菜木質素單體合成基因COMT和F5H的克隆及表達調控[D].北京:中國農業科學院,2006.
[14]武青山,武峻新,申瓊.不同基因型西葫蘆感病毒病前后幾種酶活性的變化[J].山西農業科學,2013,41(12):1368-1371.
[15]李堯臣,戚存扣.抗倒伏甘藍型油菜(Brassica napus L.)木質素含量及木質素合成關鍵基因的表達[J].江蘇農業學報,2011,27(3):481-487.
[16]郭光艷,柏峰,劉偉,等.轉錄因子對木質素生物合成調控的研究進展[J].中國農業科學,2015,48(7):1277-1287.
[17]吳立柱,王省芬,張艷,等.酸不可溶性木質素和漆酶在棉花抗黃萎病中的作用[J].作物學報,2014,40(7):1157-1163.
[18]許勇,王永健,葛秀春,等.枯萎病菌誘導的結構抗性和相關酶活性的變化與西瓜枯萎病抗性的關系[J].果樹科學,2000,17(2):123-127.
[19]毛愛軍,王永健,馮蘭香,等.水楊酸誘導辣椒抗疫病生化機制的研究[J].中國農學通報,2005,21(5):219-222,468.
[20]方東鵬,靳立梅,董利東,等.野生大豆接種大豆疫霉菌后木質素含量的變化[J].大豆科學,2015,34(1):99-102.
[21]陳塵,王政軍,曹鑫林,等.丹參肉桂酰輔酶A還原酶基因克隆與生物信息學分析[J].西北植物學報,2011,31(10):1963-1968.
[22]吳雪君,譚國飛,徐志勝,等.鴨兒芹羥基肉桂酸轉移酶基因的克隆及其對不同溫度的響應[J].西北植物學報,2015,35(7):1289-1296.
[23]譚國飛,王楓,王廣龍,等.鴨兒芹肉桂酸4-羥化酶基因的克隆與不同溫度下的表達分析[J].西北植物學報,2014,34(7):1298-1304.
[24]孫旭科.枇杷果實冷藏過程中細胞壁代謝機理及生物保鮮劑的研究[D].金華:浙江師范大學,2013.
[25]李鵬.轉錄因子NAC1在棉花纖維中的表達及在次生壁發育中的功能研究[D].武漢:華中師范大學,2011.
[26]張靜波.楊樹抗凍性生理學基礎及低溫誘導對爛皮病發生的影響[D].哈爾濱:東北林業大學,2009.
[27]Tsai C J,Kayal W E,Harding S A.Populus,the newmodel system for investigatingphenylpropanoid complexity[J].International Journal ofApplied Science and Engineering,2006,4(3):221-233.
[28]Boerjan W,Ralph J,Baucher M.Lignin biosynthesis[J].Annual ReviewofPlant Biology,2003,54(1):519-546.
[29]鄧晶.苧麻CCoAOMT基因干擾表達載體構建及其遺傳轉化[D].長沙:湖南農業大學,2008.
[30]李魏,譚曉風,陳鴻鵬.植物肉桂酰輔酶A還原酶基因的結構功能及應用潛力[J].經濟林研究,2009,27(1):7-12.
[31]Humphreys J M,Hemm M R,Chapple C.New routes for lignin biosynthesis defined by biochemical cCharacterization of recombinantferulate5-hydroxylase,amultifunctionalcytochrome P450-dependent monooxygenase[J].Proceedings of the National AcademyofSciences,1999,96(18):10045-10050.
[32]金順玉.毛竹四個木質素合成相關酶基因的克隆及組織特異性表達分析[D].北京:中國林業科學研究院,2009.
[33]周大祥,熊書.外源茉莉酸甲酯誘導竹根姜對青枯菌的抗性[J].西北植物學報,2015,35(7):1415-1420.
[34]Xu L,Zhu L,Tu L,et al.Lignin metabolism has a central role in the resistance ofcotton tothe wilt fungus verticilliumdahliae as revealed by rna-seq-dependent transcriptional analysis and histochemistry[J].Journal of Experimental Botany,2011,62(15):5607-5621.
[35]陳安和.甘藍型油菜及其親本物種C4H基因家族克隆及比較基因組學研究[D].重慶:西南大學,2006.
[36]倪志勇,王娟,呂萌,等.棉花4-香豆酸輔酶A連接酶基因克隆及原核表達[J].西北植物學報,2010,30(3):429-436.
[37]龍松華,李翔,陳信波,等.亞麻4CL基因克隆及RNAi遺傳轉化[J].西北植物學報,2014,34(12):2405-2411.
[38]Marita J M,Ralph J,Hatfield R D,et al.Nmr characterization of lignins in arabidopsis altered in the activity of ferulate 5-hydroxylase[J].Proceedings of the National Academyof Sciences,1999,96(22):12328-12332.
[39]李雪平,彭鎮華,高志民,等.抑制COMT基因表達對轉基因煙草木質素合成的影響[J].分子植物育種,2012,10(6):689-692.
[40]Pichon M,Deswartes C,Gerentes D,et al.Variation in lignin and cell wall digestibility in caffeic acid O-methyltransferase downregulated maize half-sib progenies in field experiments[J].Molecular Breeding,2006,18(3):253-261.
[41]Rastogi S,Dwivedi U N.Down regulation of lignin biosynthesis in transgenic leucaena leucocephala harboring O methyltransferase gene[J].BiotechnologyProgress,2006,22(3):609-616.
[42]劉曉晶.RNA干擾抑制的肉桂酰輔酶A還原酶和咖啡酸轉移酶還原酶對丹參木質素沉積及水溶性次生代謝物影響[D].西安:陜西師范大學,2010.
[43]Li X,Chen W,Zhao Y,et al.Downregulation of caffeoyl-coa O-methyltransferase(ccoaomt)by rna interference leads to reduced lignin production in maize straw[J].Genetics and Molecular Biology,2013,36(4):540-546.
[44]Wagner A,Tobimatsu Y,Phillips L,et al.CCoAOMT suppression modifies lignin composition in pinus radiata[J].The Plant Journal,2011,67(1):119-129.
[45]倪志勇,呂萌,馬文靜,等.棉花咖啡酰輔酶A-O-甲基轉移酶基因的克隆及表達[J].西北植物學報,2009,29(10):1946-1953.
[46]Rogers L A,Dubos C,Surman C,et al.Comparison of lignin deposition in three ectopic lignification mutants[J].New Phytologist,2005,168(1):123.
[47]M M,D O,R S.Isolation of a cdna and a genomic clone encoding cinnamate 4-hydroxylase fromarabidopsis and its expression manner in planta[J].Plant Physiology,1997,113(3):755-763.
[48]Sablowski R W,Moyano E,Culianez-macia F A,et al.A flower-specific MYBprotein activates transcription ofphenylpropanoid biosynthetic genes[J].EmboJournal,1994,13(1):37-128.
[49]Omer S,Kumar S,Khan B M.Over-expression of a subgroup 4 R2R3 Type MYB transcription factor gene from leucaena leucocephala reduces lignin content in transgenic tobacco[J].Plant Cell Reports,2013,32(1):161-171.
[50]Karpinska B,Karlsson M,Srivastava M,et al.MYB transcription factors are differentially expressed and regulated during secondary vascular tissue development in hybrid aspen[J].Plant Molecular Biology,2004,56(2):255-270.
[51]Tamagnone L,Merida A,Parr A,et al.The Am MYB308 and Am-MYB330 transcription factors from antirrhinum regulate phenylpropanoid and lignin biosynthesis in transgenic tobacco[J].The Plant Cell,1998,10(2):135-154.
[52]曹丹琴,楊健,關曉彎,等.石榴種皮木質素合成相關轉錄因子基因PgMYB的克隆與表達[J].西北植物學報,2015,35(1):23-29.
[53]邢國芳,張雁明,張魏斌,等.植物NAC轉錄因子的研究進展[J].山西農業科學,2012,40(4):409-411,423.
[54]Mitsuda N,Seki M,Shinozaki K,et al.The NAC transcription factors NST1 and NST2 of arabidopsis regulate secondary wall thickenings and are required for anther dehiscence[J].The Plant Cell,2005,17(11):2993-3006.
[55]Zhong R,Lee C,Ye Z.Functional characterization of poplar wood-associated NACdomain transcription factors[J].Plant Physiology,2010,152(2):1044-1055.
[56]Yan L,Xu C,Kang Y,et al.The heterologous expression in Arabidopsis thaliana of sorghum transcription factor SbbHLH1 downregulates lignin synthesis[J].Journal of Experimental Botany,2013,64(10):3021-3032.
[57]王進.亞麻(Linum usitatissimum)木質素合成關鍵酶基因的克隆及表達分析[D].北京:中國農業科學院,2009.
[58]Hang J,Wan R,Yan C,et al.Structural basis of pre-mRNA splicing[J].Science,2015,349:1191-1198.
[59]Yan C,Hang J,Wan R,et al.Structure of a yeast spliceosome at 3.6-angstromresolution[J].Science,2015,349:1182-1191.
[60]孟志剛,王艷玲,孟釗紅,等.利用CRISPR-Cas9系統定向誘變棉花精氨酸酶基因[C]//2015年棉花學會論文集.昌吉:中國棉花學會,2015.
Research Progress on Synthesis and Regulation of Plant Lignin
DINGXiao,CAOCairong,LI Pengbo,WUCuicui,CAOMeilian,YANGLiuliu
(Shanxi KeyLaboratoryofCotton GermplasmResources Utilization and Molecular Design Breeding,Institute ofCotton,Shanxi AcademyofAgricultural Sciences,Yuncheng044000,China)
Lignin is an important component of secondary cell wall in vascular plants and has important biological functions.The lignin distributes in machinery organization and conducting tissue in plant cell wall,which not only increases cell wall strength,improves the cell wall impermeability and the mechanical strength of the stalk,but also plays an important role in terms of resistant cells.The advanced researches on types,synthetic regulation and control plant lignin from the source by transgenic technology were reviewed.With the development of transgenic technology,it was expected to change the composition of plant lignin by more and more effective way.
lignin;synthesis and regulation;genetic engineering
Q943.2
A
1002-2481(2016)09-1406-06
10.3969/j.issn.1002-2481.2016.09.43
2016-03-16
山西省農業科學院院育種項目(Yyzjc1502);山西省財政支農項目(2015ZYZX-06);山西省農業科學院重點攻關項目(YGG1635)
丁霄(1986-),男,山西運城人,助理研究員,碩士,主要從事棉花分子育種研究工作。李朋波為通信作者。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
