子癇前期體外缺氧細胞中Nrf2和NQO1的表達變化
2017-01-06 03:14:09張展周潔宋婉玉李愛萍常愛民
山東醫藥 2016年45期
關鍵詞:氧化應激
張展,周潔,宋婉玉,李愛萍,常愛民
(鄭州大學第三附屬醫院,鄭州450052)
?
子癇前期體外缺氧細胞中Nrf2和NQO1的表達變化
張展,周潔,宋婉玉,李愛萍,常愛民
(鄭州大學第三附屬醫院,鄭州450052)
目的 探討子癇前期體外缺氧細胞中核因子E2相關因子2(Nrf2)和醌氧化還原酶1(NQO1)的表達水平及定位變化。方法 選取絨癌JAR細胞株,以含10%胎牛血清的RPMI1640培養基培養。取對數生長期細胞,貼壁后分組培養。對照組:按照之前的條件繼續培養24 h;馬來酸二乙酯(DEM)組用含50 μmol/L DEM的培養基在20% O2培養箱中繼續培養24 h;子癇前期體外模型組(缺氧組):將貼壁后的細胞置于1% O2培養箱中繼續培養24 h。采用實時熒光定量PCR法、蛋白質免疫印跡法分別檢測JAR細胞中Nrf2 mRNA及蛋白和NQO1 mRNA及蛋白的表達水平;采用細胞免疫熒光法檢測各組JAR細胞中Nrf2蛋白定位情況。結果 與對照組比較,DEM組Nrf2和NQO1 mRNA及蛋白表達升高,缺氧組Nrf2和NQO1 mRNA及蛋白表達降低(P<0.05或<0.01)。細胞免疫熒光結果顯示,DEM組中Nrf2蛋白在JAR細胞核中表達較密集,而缺氧組細胞核中只有少量表達。結論 子癇前期體外缺氧細胞中Nrf2及其目的基因NQO1表達水平降低,以細胞核中的表達降低為著。
子癇前期;缺氧;核因子E2相關因子2;醌氧化還原酶1
子癇前期(PE)是妊娠期20周后出現收縮壓≥140 mmHg和(或)舒張壓≥90 mmHg伴蛋白尿≥0.3 g/24 h,或隨機尿蛋白+[1],是圍產期婦女及新生兒死亡的主要原因。目前認為氧化應激參與了PE發病。已有研究證實,PE患者的胎盤組織和母體循環中均存在過度氧化應激,但具體機制仍不明確[2]。核因子E2相關因子2(Nrf2)是抵御氧化應激的重要轉錄因子。在正常(非應激)狀態下,Nrf2大多與Kelch樣環氧氯丙烷相關蛋白1(Keap1)結合于胞質中。當處于氧化應激狀態下,Nrf2與Keap1解離,在胞核中累積,激活其下游的抗氧化應激因子,其中包括醌氧化還原酶1(NQO1)[3]。本課題組已證實,在正常妊娠孕婦及PE患者胎盤組織中Nrf2和NQO1均有不同程度的表達[4]。另外,有文獻報道Nrf2下游因子HO-1、GCLC、GCLM在PE胎盤組織中的表達也有所降低[5]。2015年10月~2016年5月,本研究通過觀察正常或缺氧條件下以及加入馬來酸二乙酯(DEM)后Nrf2和NQO1在滋養細胞JAR中的表達水平以及Nrf2的表達定位情況,探討不同條件下JAR細胞中Nrf2和NQO1的表達變化及與PE的潛在關系。
1 材料與方法
1.1 材料 絨癌JAR細胞株購于美國ATCC公司;DEM購自上海阿拉丁公司;RNA提取試劑盒購自美國Invitrogen公司;cDNA合成試劑盒購自日本東洋紡公司;蛋白濃度BCA試劑盒購自北京康為公司;兔抗人Nrf2多克隆抗體購自美國Santa公司;鼠抗人NQO1多克隆抗體購自美國CST公司;鼠抗人β-actin單克隆抗體購自美國Abcam公司;引物序列設計與合成均由上海生工公司完成。
1.2 細胞培養及分組 JAR細胞以含10%胎牛血清的RPMI1640培養基在37 ℃、5% CO2、20% O2培養箱中培養。實驗時,取對數生長期細胞用胰酶消化,重懸后將2×105細胞接種在六孔板上,貼壁后對細胞進行以下分組培養:對照組:按照之前的條件繼續培養24 h;DEM組:用含50 μmol/L DEM的培養基在20% O2培養箱中繼續培養24 h;PE體外模型組(缺氧組):將貼壁后的細胞置于1% O2培養箱中繼續培養24 h。
1.3 JAR細胞中Nrf2、NQO1 mRNA相對表達的檢測 采用實時熒光定量PCR法。JAR細胞的總RNA按試劑盒方法提取,用Nano-2000分光光度計(A260/280)測定RNA濃度;按照cDNA合成試劑盒進行逆轉錄;熒光定量PCR總反應體系為15 μL;反應引物序列:Nrf2上游引物為5′-ACTACTCCCAGGTTGCCCA-3′,下游引物為5′-GAACAAGTGACTGAAACGTAGCC-3′;NQO1上游引物為5′-GAAAGGCTGGTTTGAGCGAG-3′,下游引物為5′-GTGGATCCCTTGCAGAGAGT-3′;β-actin mRNA作為內參基因,上游引物為5′-ATGGTGGGAATGGGTCAGAAG-3′,下游引物為5′-TCTCCATGTCGTCCCAGTTG-3′。反應條件:95 ℃預變性1 min,95 ℃變性15 s,60 ℃退火/延伸1 min,共40個循環,記錄熔解曲線及CT值,擴增產物4 ℃保存;采用2-ΔΔCT方法計算Nrf2、NQO1 mRNA的相對表達量。
1.4 JAR細胞Nrf2、NQO1蛋白相對表達的檢測 采用蛋白質免疫印跡法。用含有PMSF的細胞裂解液RIPA提取JAR細胞中總蛋白,蛋白濃度用BCA試劑盒檢測;采用5%濃縮膠和12%分離膠以進行SDS-PAGE凝膠電泳,蛋白上樣量為20 μg;使用濕式轉膜儀將凝膠中的蛋白轉至硝酸纖維素膜;使用含5%脫脂奶粉的TBST 液封閉后加入Nrf2、NQO1和內參β-actin的一抗4 ℃孵育過夜后,加入二抗孵育,洗脫二抗后用Odyssey雙色紅外激光成像系統掃描并分析蛋白條帶灰度值。
1.5 Nrf2蛋白細胞內定位檢測 采用細胞免疫熒光法。室溫下,4%的多聚甲醛固定爬好細胞的玻片15 min后用PBS洗滌3次;0.2% Triton X-100室溫通透10 min洗滌后用山羊血清室溫封閉,接著加入Nrf2的一抗放入濕盒內4 ℃孵育,過夜后加入二抗室溫孵育;滴加DAPI避光孵育染核后,使用熒光倒置顯微鏡觀察蛋白定位情況。
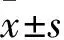
2 結果
2.1 3組Nrf2和NQO1mRNA及蛋白相對表達量的比較 與對照組比較,DEM組Nrf2、NQO1mRNA及蛋白升高,缺氧組降低(P<0.05或<0.01)。見表1。
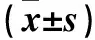
表1 3組Nrf2和NQO1 mRNA及蛋白的相對表達量比較
注:與對照組比較,*P<0.05,**P<0.01。
2.2 JAR細胞中Nrf2蛋白的定位情況 在相同的激發強度及條件下,Nrf2在各組JAR細胞中均呈陽性表達,且胞質、胞核中皆有表達。對照組中可見綠色熒光在JAR細胞質和胞核中均勻分布;DEM組中,胞質有表達,但是胞核中表達更密集;缺氧組中綠色熒光表達變弱,且胞核中只有少量表達。
3 討論
正常妊娠與子宮內環境的適應性改變有關,其根據胎兒必要的需求暫時改變了子宮內氧含量、代謝和免疫狀態。孕早期,胎兒在相對低氧的環境中發育,即氧分壓為17 mmHg(約2.4% O2)[6], 該環境能夠確保足夠的絨毛外滋養細胞侵襲入母體蛻膜以及適當的胎盤血管化和血管生成,以維持母體胎盤之間的正常血液循環。PE滋養細胞侵襲能力下降和子宮胎盤灌注不足導致胎盤氧化與抗氧化作用失衡,出現氧化應激。本次研究選用的JAR細胞來源于絨癌細胞,與胎盤滋養細胞有許多共同特征,1% O2條件下培養該細胞株作為PE的體外缺氧模型已經應用于多項研究[7,8]。
Nrf2是cap′n′collar(CNC)轉錄因子家族中一種強有力的轉錄激活因子,具有高度保守的堿性亮氨酸拉鏈結構,能與抗氧化反應元件(ARE)結合,在氧化應激和電應力誘導的多種細胞保護基因表達中起著重要作用。ARE是在多種Ⅱ相解毒酶及氧化-還原平衡蛋白的基因啟動子中發現的一種順式調節因子。正常情況下,Nrf2與其負性調節因子Keap1結合,后者可通過快速蛋白酶體降解而負性調節Nrf2。在氧化應激或親電體存在情況下,Nrf2停止降解。穩定的Nrf2在胞核積聚并與小maf蛋白偶聯,通過與ARE序列相互作用,激活目的基因,發揮保護細胞的作用[9]。NQO1即是其激活的目的基因之一。NQO1是一種專性雙電子還原酶,是兩個相互纏繞的殘基單體組成的同源二聚體。作為一種氧化代謝酶,NQO1不僅可以將內源性和外源性醌類成為氫醌類,避免有毒的半醌自由基中間體的形成,還可以激活泛醌和維生素e醌構成的抗氧化劑,對抗氧化應激。另外,NQO1也可通過清除過氧化物而發揮保護細胞的作用[10]。NQO1在上皮細胞、血管內皮細胞的胞質和人體多種組織中廣泛表達。本課題組已經證實重度PE患者胎盤組織中Nrf2和NQO1的表達水平低于正常胎盤組織,胎盤氧化應激增強,其原因可能是Nrf2功能被抑制,使得其目的基因NQO1活化降低,抗氧化能力減弱[4]。
本研究結果顯示,加入Nrf2特異性激動劑后,Nrf2和NQO1 mRNA和蛋白的表達水平均有顯著升高,表明與其他細胞和人體組織中相同,在JAR細胞中NQO1同樣受Nrf2調控;缺氧組中Nrf2的活化由于低氧而受到了抑制,該結果與本課題組在PE胎盤組織中得到的結果理論意義上相符,并與Chigusa等[5]眾多學者的研究結果一致。迄今為止,有關Nrf2的活化抑制因素所知甚少。已有研究證實,在JAR細胞中,促進Nrf2降解和蛋白表達的Keap1不受缺氧環境的影響,表明在缺氧環境下Keap1與Nrf2的抑制無關[5]。本研究結果顯示,與對照組相比,缺氧組中Nrf2在胞核中低表達,可能是缺氧抑制了Nrf2在胞核中的積聚使得與小maf蛋白偶聯的Nrf2減少,與ARE序列相互作用減弱,以至于不能正常激活其目的基因,發揮其保護細胞的作用。
綜上所述,與正常妊娠生理性缺氧相比,PE胎盤組織水平及體外細胞水平試驗均說明,缺氧可能抑制了Nfr2入核,使其無法正常激活下游的目的基因,抗氧化能力減弱,發生氧化應激。氧化應激可導致胎盤釋放包括炎癥細胞因子、抗血管生成因子、凋亡碎片在內的一系列因子,加劇母體炎癥反應和內皮細胞損傷,與PE的發病密切相關。因此,Nrf2的功能抑制可能參與了PE的發生發展,兩者也可能互為因果關系。然而,PE以及缺氧環境下滋養細胞Nrf2抑制表達的詳細機制還未明確,仍需進一步探索。
[1] 謝幸,茍文麗.婦產科學[M].8版.北京:人民衛生出版社,2013:90-97.
[2] Jauniaux E, Burton GJ. The role of oxidative stress in placental-related diseases of pregnancy[J]. J Gynecol Obstet Biol Reprod (Paris), 2016,45(8):775-785.
[3] Baird L, Dinkova-Kostova AT. The cytoprotective role of the Keap1-Nrf2 pathway[J]. Arch Toxicol, 2011,85(4):241-272.
[4] 張展,趙小辰,馮楊,等.重度子癇前期患者胎盤組織中Nrf2和NQO1的表達[J].中國婦幼保健,2016,31(6):1142-1144.
[5] Chigusa Y, Kawasaki K, Kondoh E, et al. Simvastatin inhibits oxidative stress via the activation of nuclear factor erythroid 2-related factor 2 signaling in trophoblast cells[J]. J Obstet Gynaecol Res, 2016,42(1):36-43.
[6] Rodesch F, Simon P, Donner C, et al. Oxygen measurements in endometrial and trophoblastic tissues during early pregnancy[J]. Obstet Gynecol, 1992,80(2):283-285.
[7] Plosch T, Gellhaus A, van Straten EM, et al. The liver X receptor (LXR) and its target gene ABCA1 are regulated upon low oxygen in human trophoblast cells: a reason for alterations in preeclampsia[J]. Placenta, 2010,31(10):910-918.
[8] Valbuena-Diez AC, Blanco FJ, Oujo B, et al. Oxysterol-induced soluble endoglin release and its involvement in hypertension[J]. Circulation, 2012,126(22):2612-2624.
[9] Itoh K, Mochizuki M, Ishii Y, et al. Transcription factor Nrf2 regulates inflammation by mediating the effect of 15-deoxy-Delta(12,14)-prostaglandin j(2)[J]. Mol Cell Biol, 2004,24(1):36-45.
[10] Nioi P, Hayes JD. Contribution of NAD(P)H:quinone oxidoreductase 1 to protection against carcinogenesis, and regulation of its gene by the Nrf2 basic-region leucine zipper and the arylhydrocarbon receptor basic helix-loop-helix transcription factors[J]. Mutat Res, 2004,555(1-2):149-171.
10.3969/j.issn.1002-266X.2016.45.014
R714.2
A
1002-266X(2016)45-0045-03
2016-08-16)
猜你喜歡
中成藥(2021年5期)2021-07-21 08:39:04
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
中成藥(2018年6期)2018-07-11 03:01:24
中成藥(2018年5期)2018-06-06 03:11:43
天然產物研究與開發(2016年6期)2016-06-05 10:29:26
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:13
實用中西醫結合臨床(2015年7期)2015-02-28 16:30:23
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
