安徽銅陵白姜姜瘟病病原鑒定
2017-01-12 08:29:01胡洪濤朱志剛閔勇楊自文黃
長江蔬菜·學術版 2016年12期
胡洪濤++朱志剛++閔勇++楊自文++黃大野++姚經(jīng)武++楊妮娜++龍同++程賢亮

摘 要:通過田間調查發(fā)現(xiàn),銅陵白姜姜瘟病在銅陵當?shù)乇憩F(xiàn)出木質部黃心和黑心2種癥狀,通過TTC平板分離、致病性測定、16S rDNA以及青枯菌特異性鞭毛基因(fliC)檢測表明,導致2種不同姜瘟病癥狀的病原菌均為青枯雷爾氏菌(Ralstonia solanacearum),但二者致病性存在明顯差異。
關鍵詞:銅陵;白姜;姜瘟病;青枯雷爾氏菌;病原鑒定
中圖分類號:S632.5 文獻標識碼:A 文章編號:1001-3547(2016)24-0075-04
由青枯雷爾氏菌(Ralstonia solanacearum,簡稱青枯菌)侵染造成的細菌性青枯病是世界性重要病害之一,該菌可侵染54個科的450多種植物[1],但以茄科作物,如番茄、馬鈴薯等,受害最為嚴重。生姜青枯病,因其發(fā)病快、死亡率高、無有效防治手段,所以俗稱姜瘟病[2]。安徽銅陵白姜種植歷史悠久,姜塊色白鮮嫩汁多、香味純正、品質優(yōu)良,并具藥用價值,宋朝時被列為朝廷貢品,被譽為銅陵“八寶”之一。銅陵白姜種植面積約為400 hm2,年產(chǎn)量約為4 000 t,年產(chǎn)值近億元,是當?shù)刂匾奶厣r(nóng)業(yè)產(chǎn)業(yè)[3]。銅陵市義安區(qū)是銅陵白姜主產(chǎn)區(qū),近年來該區(qū)姜瘟病的發(fā)生日益嚴重,連作田發(fā)病率50%以上,重病田甚至絕收,已經(jīng)嚴重阻礙了當?shù)厣a(chǎn)業(yè)的可持續(xù)發(fā)展[3]。盡管對姜瘟病的研究進行多年,然而對銅陵姜瘟病病原的研究甚少。為此,開展了銅陵白姜姜瘟病病原檢測的研究,以期為進一步有效防治該病提供科學依據(jù)。
1 材料與方法
1.1 樣品收集
2016年5~8月,在銅陵市義安區(qū)興化村等主要白姜產(chǎn)區(qū)分別挖取葉部呈現(xiàn)萎蔫狀的生姜姜塊,用清水將其表面泥土沖洗干凈,吸水紙吸干,冰盒保存帶回實驗室。
1.2 病原菌分離
病原菌分離均于采樣后48 h內在超凈工作臺(北京東聯(lián)哈爾儀器制造有限公司,型號A2)上完成。樣品先用無菌水沖洗3遍,晾干至表面無明顯水滴,在75%的酒精中浸泡2~3 min,再用無菌水沖洗3遍,晾干至表面無明顯水滴,用消毒刀片在姜塊中部表面切一道長2~4 cm、深約2 mm的切口,用消毒鑷子將其掰開,并用消毒牙簽蘸取姜塊中間部位汁液,在TTC平板上劃線分離[4]。將分離平板置
于28℃培養(yǎng)箱培養(yǎng)24~72 h,所得單菌落超低溫
(-80℃)保存于25%甘油管中。
1.3 致病性測定
病原菌制備:將病原分離物,接種于SPA液體培養(yǎng)基[5],搖床(北京東聯(lián)哈爾儀器制造有限公司,型號DLHR-Q200)發(fā)酵48 h(28℃,150 r/min)后,將分光光度計(上海光學儀器721型)OD560值調至1.0,測定菌液濃度約為1×108 CFU/mL,備用。
將溫室生長至5~8葉的銅陵白姜植株從營養(yǎng)缽中取出,用清水將根部沖洗干凈,用消毒針在莖基部刺3個深度和間隔均約為1 cm的針孔,而后將根系置入病原菌發(fā)酵液中浸泡20 min。無菌水處理作為對照。每處理5株,重復2次。處理后的植株立即種植于營養(yǎng)缽中,置于人工氣候箱(光周期
12 h/8 h,溫度30℃,濕度85%)內培養(yǎng)7~10 d。
1.4 16S rDNA檢測
純化后的病原分離物直接用PCR擴增,采用16S rDNA通用引物對(F:5'-AGAGTTTGATCCTGGCTCAG-3',R:5'-GGTTACCTTGTTACGACTT-3'),PCR反應體系(50 μL):2×ES Taq Mix 25μL,正反引物各1 μL,模板1 μL,ddH2O 22 μL。反應條件:95℃ 1 min,95℃ 30 s,58℃ 1 min,72℃ 1.5 min,30個循環(huán),72℃ 5 min。引物合成和PCR產(chǎn)物測序均由生工生物工程(上海)股份有限公司提供。
1.5 青枯菌鞭毛基因(fliC)基因檢測
根據(jù)NCBI中青枯菌鞭毛基因(fliC)序列設計引物對(F:5'-GAACGCCAACGGTGCGAACT-3',R:5'-GGCGGCCTTCAGGGAGGTC-3')。反應體系和條件同1.4。
1.6 進化樹分析
進化樹分析在MEGA 7.0中完成。首先,采用Clustalw進行DNA序列比對(DNA Weight Matrix:IUB,其他采用默認參數(shù))。進化樹的構建采用Maxium Likehood方法、Tamura-Nei模式。
2 結果與分析
2.1 姜瘟病田間癥狀
在銅陵義安村田間調查表明,感染植株在患病初期,葉片褪綠,葉尖和葉緣先黃化,逐漸萎縮反卷下垂(圖1A、B),病葉逐步從基部向上發(fā)展,至全株枯死。病株莖基部逐漸變軟,維管束變色褐化、呈水漬狀,擠壓時,有污白色菌膿流出。患病姜塊變軟、維管束變色,根據(jù)維管束顏色,可分為黃色和黑色2種,俗稱為黃心(圖1C-a)和黑心(圖1C-b)。黑心和黃心患病植株的姜塊木質部分別呈現(xiàn)黃色、黑色,而地上部癥狀無明顯差異,但相對于黃心癥狀,黑心癥狀病株萎蔫更快,病程更短。
2.2 致病性測試
致病性測試結果顯示,在接種后12 d后,植株葉片呈現(xiàn)萎蔫狀、卷縮,葉尖和葉緣失綠黃化,與田間病株癥狀完全一致(圖2),表明所分離的病原物為姜瘟病病原。然而,接種黑心癥狀病原的植株萎蔫程度更甚、失綠更為嚴重,提示黑心癥狀病原的致病性可能更強。
2.3 病原形態(tài)鑒定
導致白姜黑心和黃心癥狀的病原分離物在TTC培養(yǎng)基菌落均為近圓形或梭形,中間略微凸起、中間粉紅色至紅色、周圍為白色。病原菌均為單胞、桿狀,大小為(0.5~0.7)μm×(1.5~2.0)μm,革蘭氏染色陰性,而結晶紫染色顯示兩極著色深,與前人報道一致[6,7]。
2.4 16S rDNA測序
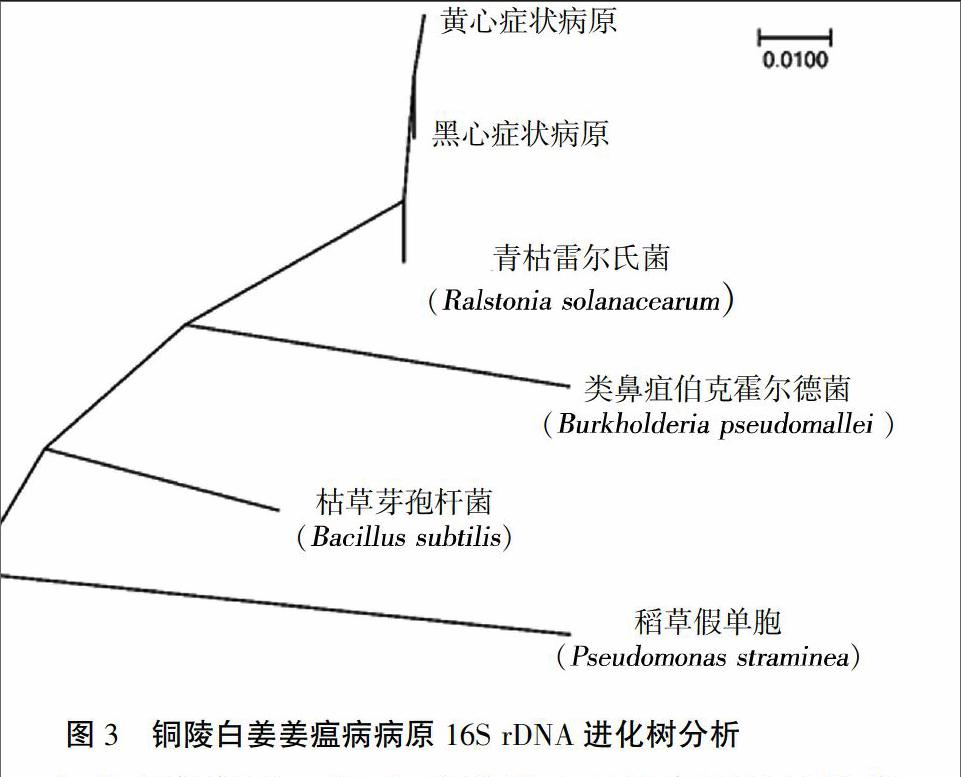
采用16S rDNA通用引物對擴增和測序,結果表明,2種癥狀病原分離物的16S rDNA產(chǎn)物長度均為1 439 bp(NCBI序列號:KX785159.1和KX785160.1)。黃心和黑心癥狀病原的16S rDNA序列與NCBI里保存的青枯雷爾氏菌有高度相似性(>99%)(圖3),而與其他菌如Bukholderia、Bacillus等,親緣關系相對較遠,表明無論是黃心還是黑心癥狀病原菌均為青枯雷爾氏菌。
2.4 青枯菌特異性鞭毛基因(fliC)檢測
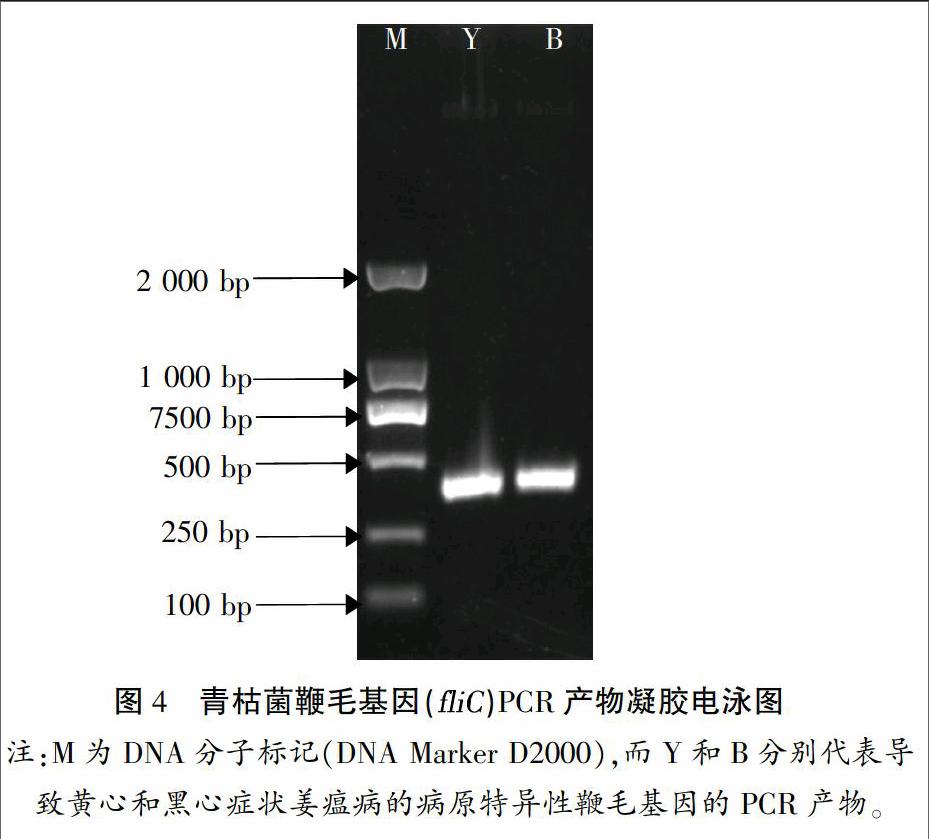
為進一步確定銅陵白姜姜瘟病病原,根據(jù)前人研究結果[8],針對青枯菌特異性鞭毛基因(fliC)在NCBI上設計了1對引物,用于青枯菌特檢測,結果如圖4所示。測序結果顯示,黃心和黑心癥狀的青枯菌fliC基因PCR產(chǎn)物長度分別為368、370 bp。由于5'端20~30 bp和3'端10 bp測序質量較低,因此,將兩端低質量的堿基去掉,所得序列與NCBI中所保存的青枯菌fliC基因序列進行比對。如圖5所示,導致白姜黃心和黑心癥狀的青枯菌fliC基因序列與NCBI保存的青枯菌完全一致。綜上所述,可以確定導致銅陵白姜黃心和黑心癥狀姜瘟病的病原均為青枯菌。
3 討論與結論
銅陵白姜為我國具有地方特色的名特優(yōu)農(nóng)產(chǎn)品,而近年來,日益嚴重的姜瘟病已經(jīng)給當?shù)匕捉a(chǎn)造成嚴重的經(jīng)濟損失,制約了白姜產(chǎn)業(yè)的健康、可持續(xù)發(fā)展。然而長期以來,缺乏對其病原菌的科學鑒定。本研究通過實地調研,發(fā)現(xiàn)姜瘟病病株表現(xiàn)為黃心和黑心2種不同癥狀,而通過TTC平板分離、形態(tài)學、致病性測定和分子生物學等手段,對銅陵白姜姜瘟病病株進行分離和鑒定,明確其病原均為青枯菌,但二者的致病性存在明顯差異。
16S rDNA序列分析是植物病原細菌鑒定和分類的重要手段和依據(jù) [9]。經(jīng)序列比較和系統(tǒng)進化樹分析證明銅陵白姜姜瘟病2種癥狀的病原與已知青枯菌在同一分支。然而,盡管2種癥狀病原遺傳距離相當近,但仍然存在微小差距。盡管青枯菌特異性鞭毛基因擴增結果表明,黃心和黑心癥狀病原的序列與已知青枯菌完全一致,但由于致病性和16S rDNA序列存在差異,提示導致黃心和黑心癥狀青枯菌可能為不同的株系或者分化型[8]。前人研究表明,不同地區(qū)或寄主的青枯菌表現(xiàn)出明顯生理分化或菌系多樣性[10],而在地理位置和寄主相同的情況下,發(fā)現(xiàn)毒力差異性的青枯菌的相關報道尚不多見。青枯菌的致病性受多種基因調控[11],如果能從基因組學角度,深入研究其在DNA和轉錄組水平方面的差異,將有助于揭示青枯菌致病性的調控機制[12]。
參考文獻
[1] Hayward A C. Biology and epidemiology of bacterial wilt caused by Pseudomonas solanacearum[J]. Annual Review of Phytopathology, 1991(29): 65-87.
[2] 劉銘,張敏,戢俊臣,等.中國姜瘟病的研究進展[J].中國農(nóng)學通報,2005(6):337-340.
[3] 趙友前,趙德明,汪濤,等.銅陵縣白姜姜瘟病的致病因素及防治技術[J].現(xiàn)代農(nóng)業(yè)科技,2012(7):177.
[4] 方中達.植病研究方法[M].北京:中國農(nóng)業(yè)出版社,1998:156-211.
[5] 車建美,劉波,張彥,等.青枯雷爾氏菌致病性生物測定方法的研究[J].福建農(nóng)業(yè)學報,2011,26(5):804-807.
[6] 陳莉,高智謀,楊自保,等.安徽省姜瘟病病原細菌鑒定及有效藥劑篩選[J].安徽農(nóng)業(yè)科學,2007,35(18):5 479,
5 516.
[7] 趙志祥,嚴婉榮,陳圓,等.海南生姜青枯病病原菌鑒定[J]. 基因組學與應用生物學,2015,34(4):763-768.
[8] Sch?觟nfeld J, Heuer H, Van Elsas J D, et al. Specific and sensitive detection of Ralstonia solanacearum in soil on the basis of PCR amplification of fliC fragments [J]. Applied Environmental Microbiology, 2003, 69(12): 7 248-7 256.
[9] Weisburg W G, Barns S M, Pelletier D A, et al. 16S ribosomal DNA amplification for phylogenetic study[J]. Journal of Bacteriology, 1991, 173(2): 697-703.
[10] 徐進,顧鋼,潘哲超,等.福建煙草青枯菌演化型及生化變種鑒定研究[J].中國煙草學報,2010,16(6):66-71.
[11] 蔡劉體,劉艷霞,石俊雄.青枯菌致病力的主要決定因子研究進展[J].貴州農(nóng)業(yè)科技,2015,43(8):109-113.
[12] Salanoubat M, Genin S, Artiguenave F, et al. Genome sequence of the plant pathogen Ralstonia solanacearum [J]. Nature, 2002, 415(6 871): 497-502.
