棉花與模式植物DGAT2基因的鑒定與分析
2017-02-05 19:56:51張曉瓊喬琳胡利宗廖立冰王
江蘇農(nóng)業(yè)科學(xué) 2016年10期
張曉瓊++喬琳++胡利宗++廖立冰++王秋霞++解恒昌++周琳

摘要:二脂酰甘油酰基轉(zhuǎn)移酶Ⅱ(DGAT2)是催化三酰甘油生物合成的關(guān)鍵酶,利用生物信息學(xué)手段在二倍體棉花和6種模式植物中共鑒定出25個(gè)DGAT2基因,詳細(xì)剖析這些基因的結(jié)構(gòu)、序列和進(jìn)化特征。基因結(jié)構(gòu)分析表明,絕大多數(shù)植物包含8個(gè)或9個(gè)外顯子,且內(nèi)含子相位高度保守,說(shuō)明該基因結(jié)構(gòu)起源于陸生植物。蛋白序列分析顯示,核心保守基序1、2、3與功能結(jié)構(gòu)域相互重疊,說(shuō)明功能結(jié)構(gòu)域起源于藻類植物。進(jìn)化分析顯示:在棉花中,DGAT2基因發(fā)生特異性擴(kuò)增,其分子機(jī)制為串聯(lián)重復(fù);結(jié)合滑動(dòng)窗口方法和位點(diǎn)模型選擇壓力檢測(cè)結(jié)果,DGAT2蛋白均受制于負(fù)選擇作用。這些結(jié)果為棉花及模式植物DGAT2的功能研究提供幫助。
關(guān)鍵詞:二酰甘油酰基轉(zhuǎn)移酶;保守基序;棉花;模式植物;進(jìn)化
中圖分類號(hào): S562.03;Q78文獻(xiàn)標(biāo)志碼: A文章編號(hào):1002-1302(2016)10-0069-04
收稿日期:2015-08-28
基金項(xiàng)目:周口師范學(xué)院高層次人才科研啟動(dòng)經(jīng)費(fèi)(編號(hào):ZKNU2014109);周口師范學(xué)院大學(xué)生科研創(chuàng)新基金(編號(hào):ZKNUD15037)。
作者簡(jiǎn)介:張曉瓊(1991—),女,河南安陽(yáng)人,碩士研究生,主要從事植物遺傳學(xué)研究。E-mail:zxqxiaoqiong00@163.com。
通信作者:胡利宗,博士,副教授,主要從事植物遺傳學(xué)研究。E-mail:hulizong@126.com。二酰甘油酰基轉(zhuǎn)移酶(DGAT,EC 2.3.1.20)是催化三酰甘油(TAG)合成最后一步的關(guān)鍵酶[1]。目前,在植物中存在3種類型的DGAT,即DGAT1、DGAT2和DGAT3[2]。前人研究表明,DGAT1、DGAT2和DGAT3隸屬于不同基因家族,其蛋白序列相似性極低,但三者均具有二脂酰甘油酰基轉(zhuǎn)移酶活性,過(guò)表達(dá)這些基因均可不同程度提高植物種子含油量[3]。前人研究顯示,DGAT酶參與種子油脂合成、種子萌發(fā)與發(fā)育以及幼苗發(fā)育等眾多生物學(xué)過(guò)程,具有多樣化的功能[4]。自Lardizabal等成功克隆MrDGAT2基因以來(lái),在人類、擬南芥和藻類等物種中均克隆出DGAT2基因[5-7]。DGAT2基因功能研究顯示,DGAT2基因具有底物特異性,優(yōu)先利用特殊脂肪酸進(jìn)行三酰甘油合成[8]。因此,DGAT2基因的相關(guān)研究越來(lái)越受到人們的重視。棉花不僅是一種重要的經(jīng)濟(jì)作物,還是天然纖維與油料作物。雖然在棉花纖維發(fā)育、纖維品質(zhì)以及抗逆性等方面開展了許多研究,但棉花DGAT2基因的研究仍未見(jiàn)報(bào)道。此外,二倍體雷蒙德氏棉[9]和亞洲棉[10]全基因組序列測(cè)定已經(jīng)被完成,這為在棉花中研究DGAT2基因提供了數(shù)據(jù)資源。利用生物信息學(xué)手段鑒定棉花DGAT2基因,并剖析這些基因的結(jié)構(gòu)、保守基序、功能結(jié)構(gòu)域以及系統(tǒng)進(jìn)化關(guān)系。這些結(jié)果能為闡明棉花油脂積累的分子機(jī)制奠定基礎(chǔ),為進(jìn)一步研究棉花DGAT2基因的功能提供線索。
1材料與方法
1.1物種抽樣與基因序列下載
本研究涉及的物種包括亞洲棉、雷蒙德氏棉和其他6種模式植物。序列下載步驟為:在NCBI數(shù)據(jù)庫(kù)直接檢索并下載已知的DGAT2基因序列,以這些序列為檢索序列,利用BLAST工具在默認(rèn)參數(shù)條件下分別搜索相關(guān)數(shù)據(jù)庫(kù)CottonGen(http://www.cottongen.org/)、Greenphyl V4(http://www.greenphyl.org/cgi-bin/index.cgi)和Phytozome V9.1(http://www.phytozome.net/),最終獲取棉花及其他模式植物的DGAT2基因。并利用Pfam工具[11]確定這些基因是否含有DAGAT(PF03982)。
1.2序列特征與蛋白特性分析
通過(guò)DNA和cDNA的比較,棉花與模式植物DGAT2基因外顯子和內(nèi)含子的組織結(jié)構(gòu)模式被鑒定,由GSDS 2.0工具繪制[12]。MEME服務(wù)器被用于剖析棉花與模式植物DGAT2蛋白的保守基序組成模式,參數(shù)設(shè)置如下:保守基序氨基酸長(zhǎng)度為6~180 bp;保守基序數(shù)目最大值為10;E閾值為 0.000 01,其他參數(shù)默認(rèn)[13]。此外,利用HMMTOP工具剖析DGAT2蛋白的跨膜結(jié)構(gòu)域的數(shù)目與位置[14]。
1.3系統(tǒng)進(jìn)化與選擇壓力檢測(cè)
為闡明DGAT2蛋白的系統(tǒng)進(jìn)化關(guān)系,利用Clustal X軟件[15]在默認(rèn)條件下對(duì)DGAT2蛋白進(jìn)行序列比對(duì),其比對(duì)結(jié)果直接導(dǎo)入MEGA軟件[16]中,輸出進(jìn)化樹并進(jìn)行細(xì)微調(diào)整。在進(jìn)化樹的末端,可鑒定出同源基因?qū)Γ瑫r(shí)基于滑動(dòng)窗口方法剖析同源基因?qū)Φ倪x擇壓力,其計(jì)算由DnaSP 5.10完成,參數(shù)設(shè)置為:滑動(dòng)窗口100 bp和步長(zhǎng)10 bp[17]。此外,利用Pal2nal工具準(zhǔn)備所有DGAT2基因的密碼子比對(duì)序列[18],利用位點(diǎn)特異模型對(duì)DGAT2基因進(jìn)行選擇壓力分析,其計(jì)算過(guò)程由PAML 3.15軟件包中的CODEML程序完成[19]。
2結(jié)果與分析
2.1棉花與模式植物DGAT2鑒定與蛋白序列特征
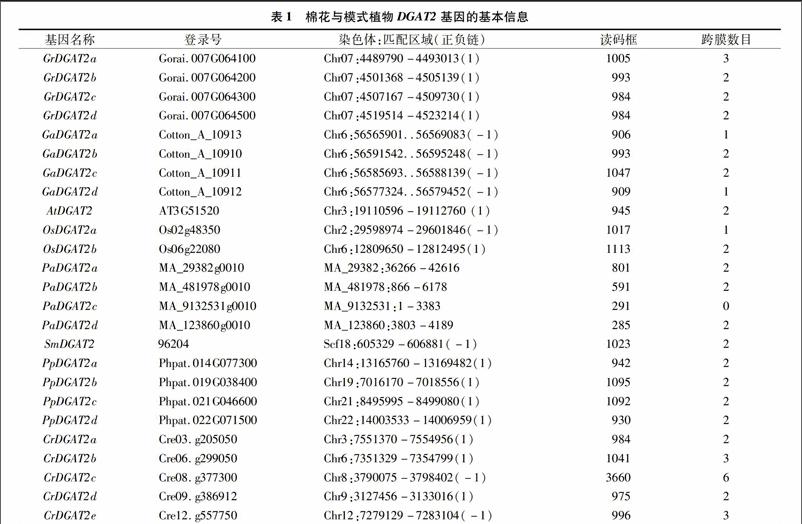
利用Blast工具在2種棉花與6種模式植物中共鑒定出25個(gè)DGAT2基因,這些基因的位點(diǎn)標(biāo)識(shí)符、染色體、正負(fù)鏈位置以及基因組匹配區(qū)段均被列出(表1)。功能結(jié)構(gòu)域分析表明,DGAT2基因編碼的蛋白均具有典型的功能結(jié)構(gòu)域DAGAT(PF03982)。聚類分析顯示,2種棉花DGAT2與擬南芥和水稻DGAT2聚在一起形成了一個(gè)雙子葉植物類群;低等植物DGAT2傾向于聚為一類,但有些低等植物DGAT2蛋白單獨(dú)形成一個(gè)分支(圖1-A)。此外,利用HMMTOP工具分析DGAT2蛋白的跨膜結(jié)構(gòu),結(jié)果顯示,除PaDGAT2c外,棉花與模式植物DGAT2蛋白均具有至少1個(gè)跨膜結(jié)構(gòu)域(表1)。
利用MEME軟件對(duì)DGAT2蛋白的保守基序進(jìn)行分析,結(jié)果表明,在25個(gè)棉花和模式植物DGAT2蛋白中存在10個(gè)保守基序(Motif1~10),共形成13種保守基序組織模式,其中6個(gè)棉花DGAT2蛋白共享的保守基序組成模式為 Motif4-6-2-1-3,而AtDGAT2、OsDGAT2a/b和PpDGAT2d等共享的保守基序模式為Motif9-6-2-1-3(圖1-B)。就單個(gè)保守基序而言,所有DGAT2蛋白保守基序被分為兩大類型,即種屬普遍類型和種屬特異類型。保守基序1、2、3、6、9幾乎存在于所有DGAT2蛋白中,屬于種屬普遍類型;但保守基序4、5、7、8、10表現(xiàn)出不同程度的種屬特異性,屬于種屬特異類型。此外,DGAT2蛋白的DAGAT(PF03982) 功能結(jié)構(gòu)域與保守基序1、2、3、6、9相互重疊,這說(shuō)明植物的5個(gè)保守基序均可能與DGAT2活性高度相關(guān)。
2.2棉花與模式植物DGAT2基因的外顯子與內(nèi)含子組成模式
通過(guò)比較DNA與CDS序列,棉花與模式植物DGAT2基因的結(jié)構(gòu)特點(diǎn)均被闡明,并鑒定其內(nèi)含子相位。根據(jù)外顯子數(shù)目,這些DGAT2基因結(jié)構(gòu)被分為三大類型:(1)8個(gè)外顯子類型,這種類型的基因包括GrDGAT2a、GrDGAT2d和AtDGAT2等5個(gè)基因;(2)9個(gè)外顯子類型,這種類型的基因包括GrDGAT2b、GaDGAT2a和OsDGAT2a等11個(gè)基因;(3)2、3、5、6、7、13個(gè)或14個(gè)外顯子類型,這類基因包括CrDGAT2a、PpDGAT2b和PaDGAT2c等9個(gè)基因(圖1-C)。結(jié)合DGAT2蛋白聚類結(jié)果可知,位于同一進(jìn)化分支的基因往往具有相同或相似的基因結(jié)構(gòu),甚至外顯子和內(nèi)含子長(zhǎng)度、內(nèi)含子相位具有高度保守性;棉花DGAT2蛋白聚為一類,具有高度相似的基因結(jié)構(gòu),并且這些基因的內(nèi)含子相位高度保守;此外,有些物種DGAT2蛋白單獨(dú)形成一個(gè)分支,具有獨(dú)特的基因結(jié)構(gòu)如PaDGAT2d、CrDGAT2a和PaDGAT2b,它們分別擁有2、6、5個(gè)外顯子,并且其內(nèi)含子相位變異較大。總之,棉花與模式植物DGAT2基因的結(jié)構(gòu)多數(shù)以8個(gè)或9個(gè)外顯子形式存在,尤其是在高等植物中該基因的結(jié)構(gòu)具有較高的保守性。
2.3棉花與模式植物DGAT2基因的系統(tǒng)進(jìn)化關(guān)系
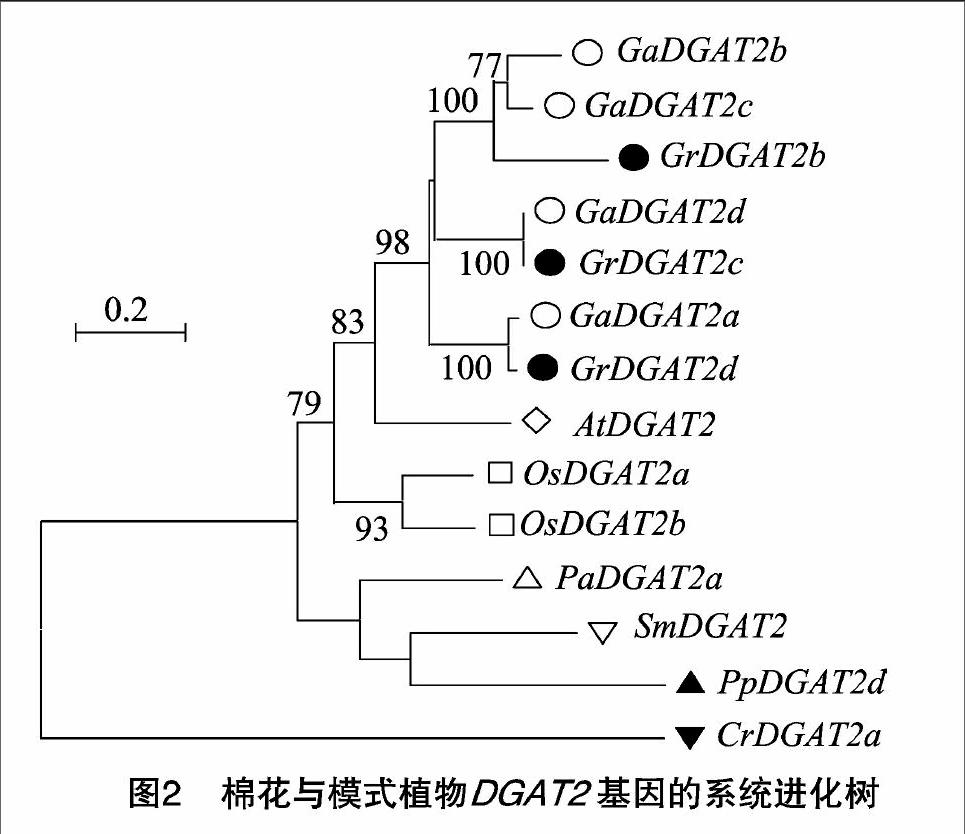
在構(gòu)建棉花與模式植物DGAT2基因的進(jìn)化樹前,移走覆蓋度低于80%、相似度低于30%的序列,進(jìn)而構(gòu)建由亞洲棉、雷蒙德氏棉和水稻等8種植物的14個(gè)DGAT2蛋白形成的進(jìn)化樹。系統(tǒng)進(jìn)化分析表明,藻類植物CrDGAT2a蛋白單獨(dú)形成1個(gè)進(jìn)化分支,與其他植物具有較遠(yuǎn)的親緣關(guān)系;其他苔蘚類、蕨類、裸子與被子植物DGAT2蛋白明顯被分為2個(gè)進(jìn)化分支,其中被子植物DGAT2蛋白形成1個(gè)進(jìn)化分支,蘚類、蕨類和裸子植物DGAT2形成另一個(gè)進(jìn)化分支(圖2)。此外,在進(jìn)化樹末端分支上,存在4對(duì)同源基因,其中2對(duì)屬于直系同源基因,2對(duì)屬于旁系同源基因;旁系同源基因包括GaDGAT2b/GaDGAT2c和OsDGAT2a/OsDGAT2b,它們起源于物種特異的基因擴(kuò)增;直系同源基因包括GrDGAT2c/GaDGAT2d和GaDGAT2a/GrDGAT2d,它們起源于物種分化事件(圖2)。
2.4基于同源對(duì)水平的棉花與模式植物DGAT2基因的選擇壓力
為考察進(jìn)化樹末端的4對(duì)同源基因之間是否經(jīng)歷適應(yīng)性進(jìn)化,本研究利用DnaSP 5.10軟件對(duì)4對(duì)DGAT2同源基因進(jìn)行滑動(dòng)窗口分析,參數(shù)設(shè)置:窗口尺寸為100 bp,步移長(zhǎng)度為10 bp。結(jié)果顯示,4對(duì)同源基因的ω平均值從大到小為044(GrDGAT2d/GaDGAT2a)>0.31(GaDGAT2b/GaDGAT2c)>016(OsDGAT2b/OsDGAT2a)>0.15(GaDGAT2d/GrDGAT2c)。就旁系同源基因而言,亞洲棉旁系基因?qū)aDGAT2b/GaDGAT2c和水稻旁系同源基因?qū)sDGAT2b/OsDGAT2a的每個(gè)步移窗口對(duì)應(yīng)的進(jìn)化速率具有異質(zhì)性,但所有的步移窗口的ω值均小于1,這充分說(shuō)明這2對(duì)旁系同源基因均受控于負(fù)選擇作用,也揭示了基因重復(fù)之后并未發(fā)生適應(yīng)性進(jìn)化(圖3-A)。同理,在2種棉花物種中,直系同源基因?qū)rDGAT2d/GaDGAT2a 和GaDGAT2d/GrDGAT2c的ω均不大于1,這說(shuō)明物種分化產(chǎn)生重復(fù)基因受到負(fù)選擇作用,這可能與功能限制密切相關(guān)(圖3-B)。
2.5基于類群水平的棉花與模式植物DGAT2基因的選擇壓力
以亞洲棉、雷蒙德氏棉和水稻等8種植物的14個(gè)DGAT2基因?yàn)檠芯繉?duì)象,利用位點(diǎn)模型對(duì)DGAT2基因進(jìn)行選擇壓力檢測(cè),鑒定出正選擇位點(diǎn),并利用貝葉斯后驗(yàn)概率推斷每個(gè)正選擇位點(diǎn)概率值。似然比檢驗(yàn)表明,2ΔlnL=271.62>13277(M0/M3),這說(shuō)明M3極顯著優(yōu)于M0(P<0.01),因此,這些DGAT2蛋白位點(diǎn)間的進(jìn)化速率明顯具有異質(zhì)性;就M2a和M1a比較而言,似然比檢驗(yàn)值2ΔlnL=0<5.991,說(shuō)明M2a并不優(yōu)于M1a模型,DGAT2基因可能未經(jīng)歷適應(yīng)性進(jìn)化;同理,2ΔlnL=0<5.991(M7/M8),說(shuō)明M8模型并不優(yōu)于M7模型,進(jìn)而可推知棉花與模式植物DGAT2基因中并沒(méi)有鑒定到正選擇氨基酸位點(diǎn)(表2)。結(jié)合似然比測(cè)驗(yàn)和貝葉斯推斷分析可知,棉花與模式植物DGAT2基因均受制于負(fù)選擇作用,這意味著DGAT2具有高度保守性,這可能與嚴(yán)格的功能限制相關(guān)。
3結(jié)論與討論
DGAT2基因廣泛存在于真菌、藻類、植物和動(dòng)物基因組中,尤其是在藻類植物中通常以多拷貝形式存在[1,3,20-21],這說(shuō)明DGAT2基因起源較早,是一個(gè)古老的基因家族。與藻類植物相比,被子植物中DGAT2基因數(shù)目較少,絕大多數(shù)物種僅包括1個(gè)或2個(gè)成員。但令人感興趣的是,在2種二倍體棉花中均發(fā)現(xiàn)了4個(gè)DGAT2基因,并且在2種二倍體棉花中DGAT2基因均以串聯(lián)重復(fù)方式排列在同一個(gè)染色體區(qū)段內(nèi),這說(shuō)明棉花DGAT2基因發(fā)生了物種特異性串聯(lián)重復(fù)。基因結(jié)構(gòu)和蛋白保守基序分析顯示,所有物種DGAT2序列特征具有較高的多樣性,但在被子植物中DGAT2基因的結(jié)構(gòu)與蛋白基序均表現(xiàn)出高度保守性,這說(shuō)明被子植物DGAT2蛋白受到嚴(yán)格的功能限制,具有高度相似的生物學(xué)功能,這與前人的研究結(jié)果[21-22]基本一致。此外,水稻和棉花的DGAT2同源基因?qū)x擇壓力檢測(cè)顯示,同源基因?qū)Ψ只蟛](méi)有發(fā)生適應(yīng)性進(jìn)化。同時(shí),基于位點(diǎn)模型對(duì)DGAT2蛋白進(jìn)行選擇壓力分析,結(jié)果表明植物的14個(gè)DGAT2蛋白均受控于負(fù)選擇作用,受到嚴(yán)格的功能限制作用[21]。總之,二倍體棉花基因組中存在4個(gè)DGAT2基因,這些基因呈現(xiàn)串聯(lián)重復(fù)特征,并且棉花DGAT2基因受純凈選擇作用,這與其功能重要性密切相關(guān)。這些結(jié)果揭示了DGAT2基因的序列特征與進(jìn)化關(guān)系,為其功能研究提供了理論基礎(chǔ)。
參考文獻(xiàn):
[1]Weselake R J,Taylor D C,Rahman M H,et al. Increasing the flow of carbon into seed oil[J]. Biotechnol Advances,2009,27(6):866-878.
[2]唐桂英,柳展基,單雷. 二酰基甘油酰基轉(zhuǎn)移酶 (DGAT) 研究進(jìn)展[J]. 中國(guó)油料作物學(xué)報(bào),2010,32(2):320-328.
[3]Turchetto-Zolet A C,Maraschin F S,de Morais G L,et al. Evolutionary view of acyl-CoA diacylglycerol acyltransferase (DGAT),a key enzyme in neutral lipid biosynthesis[J]. BMC Evolutionary Biology,2011,11(1):263.
[4]Chai Y,Hao X,Yang X,et al. Validation of DGAT1-2 polymorphisms associated with oil content and development of functional markers for molecular breeding of high-oil maize[J]. Molecular Breeding,2012,29(4):939-949.
[5]Lardizabal K D,Mai J T,Wagner N W,et al. DGAT2 is a new diacylglycerol acyltransferase gene family purification,cloning,and expression in insect cells of two polypeptides from Mortierella ramanniana with diacylglycerol acyltransferase activity[J]. Journal of Biological Chemistry,2001,276(42):38862-38869.
[6]房逢立,吳洪,周志剛. 缺刻緣綠藻二酰甘油酰基轉(zhuǎn)移酶2(DGAT2)的基因特性與功能鑒定[J]. 水產(chǎn)學(xué)報(bào),2013,37(8):1162-1172.
[7]袁崢嶸,柳小春,馬海明,等. 二脂酰甘油酰基轉(zhuǎn)移酶2(DGAT2)基因研究進(jìn)展[J]. 遺傳,2008,30(3):289-294.
[8]Li R,Yu K,Hildebrand D F. DGAT1,DGAT2 and PDAT expression in seeds and other tissues of epoxy and hydroxy fatty acid accumulating plants[J]. Lipids,2010,45(2):145-157.
[9]Paterson A H,Wendel J F,Gundlach H,et al. Repeated polyploidization of Gossypium genomes and the evolution of spinnable cotton fibres[J]. Nature,2012,492(7429):423-427.
[10]Li F,F(xiàn)an G,Wang K,et al. Genome sequence of the cultivated cotton Gossypium arboreum[J]. Nature Genetics,2014,46(6):567-572.
[11]Bateman A,Birney E,Cerruti L,et al. The Pfam protein families database[J]. Nucleic Acids Research,2002,30(1):276-280.
[12]郭安源,朱其慧,陳新,等. GSDS:基因結(jié)構(gòu)顯示系統(tǒng)[J]. 遺傳,2007,29(8):1023-1026.
[13]Bailey T L,Williams N,Misleh C,et al. MEME:discovering and analyzing DNA and protein sequence motifs [J]. Nucleic Acids Research,2006,34:369-373.
[14]Tusnady G E,Simon I. The HMMTOP transmembrane topology prediction server[J]. Bioinformatics,2001,17(9):849-850.
[15]Thompson J D,Gibson T J,Plewniak F. The CLUSTAL_X windows interface:flexible strategies for multiple sequence alignment aided by quality analysis tools[J]. Nucleic Acids Research,1997,25(24):4876-4882.
[16]Tamura K,Dudley J,Nei M,et al. MEGA4:molecular evolutionary genetics analysis (MEGA) software version 4.0[J]. Molecular Biology and Evolution,2007,24(8):1596-1599.
[17]Librado P,Rozas J. DnaSP v5:a software for comprehensive analysis of DNA polymorphism data[J]. Bioinformatics,2009,25(11):1451-1452.
[18]Suyama M,Torrents D,Bork P. PAL2NAL:robust conversion of protein sequence alignments into the corresponding codon alignments[J]. Nucleic Acids Research,2006,34:609-612.
[19]Yang Z. PAML4:Phylogenetic analysis by maximum likelihood[J]. Molecular Biology and Evolution,2007,24(8):1586-1591.
[20]朱紅霞,胡利宗,鄧小莉,等. 三種豆科植物 DGAT1 基因家族的分子特征與進(jìn)化分析[J]. 生物技術(shù)通報(bào),2011,3(10):163-166.
[21]Cao H. Structure-function analysis of diacylglycerol acyltransferase sequences from 70 organisms[J]. BMC Research Notes,2011,4(1):249.
[22]陳振璽,王鵬,蓋江濤,等. 在基因家族背景下對(duì)四種植物中DGAT2的鑒定和序列分析[J]. 植物科學(xué)學(xué)報(bào),2015,33(2):186-194.陶虎,索效軍,李曉峰,等. 山羊miR-27a靶基因預(yù)測(cè)及生物信息學(xué)分析[J]. 江蘇農(nóng)業(yè)科學(xué),2016,44(10):73-75.
