過表達Nampt通過激活NF-κB誘導心肌細胞肥大
2017-02-08 05:31:44黃珺珺劉夏雯黃碧云
中國藥理學通報 2017年1期
關鍵詞:檢測
蔡 軼,黃珺珺,劉夏雯,黃碧云,朱 柳,吳 波
(廣州醫科大學藥學院藥物研發中心,廣東 廣州 511436)
◇論 著◇
過表達Nampt通過激活NF-κB誘導心肌細胞肥大
蔡 軼,黃珺珺,劉夏雯,黃碧云,朱 柳,吳 波
(廣州醫科大學藥學院藥物研發中心,廣東 廣州 511436)
目的 研究過表達尼克酰胺磷酸核糖轉移酶(nicotinamide phosphoribosyltransferase, Nampt)對心肌細胞肥大的影響,并初步探討NF-κB在其中的作用。方法 心肌細胞給予苯腎上腺素(PE)或過表達Nampt處理,采用Real-time PCR檢測Nampt、ANP和BNP的mRNA表達,利用Western blot檢測Nampt的蛋白水平,使用免疫熒光檢測心肌細胞表面積,采用雙熒光素酶報告基因的方法檢測NF-κB依賴的轉錄活性。結果 過表達Nampt能夠誘導ANP和BNP等肥大基因的激活,增加心肌細胞表面積;過表達Nampt能夠增加NF-κB依賴的轉錄活性,而沉默NF-κB的p65亞基能夠部分取消Nampt對心肌細胞肥大的調控作用。結論 過表達Nampt能夠誘導心肌細胞肥大,其機制可能與NF-κB的激活有關。
Nampt;心肌肥大;NF-κB;苯腎上腺素;ANP; BNP
心肌肥大是一個非常重要的病理進程,它是心臟在病理狀態下維持適當收縮功能的代償反應,主要表現為心肌細胞體積增大、重量增加、間質細胞增生以及心肌重構等。長期的心肌肥大可導致冠心病、心律失常、心力衰竭,是心功能惡化及心源性死亡的獨立危險因素,因此,研究心肌肥大的分子機制具有十分重要的意義[1-3]。
尼克酰胺磷酸核糖轉移酶(nicotinamide phosphoribosyltransferase, Nampt),又名內臟脂肪素visfatin,或前B細胞克隆增強因子PBEF,是哺乳動物生命活動中不可或缺的蛋白[4]。在哺乳動物細胞中,Nampt是煙酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide,NAD)生物合成過程中的限速酶,廣泛表達于心、腦、肺、骨髓、肝臟、肌肉等機體各組織和器官中,與糖尿病、肥胖、動脈粥樣硬化、血管內皮炎性損傷等密切相關[5-7],提示Nampt在心血管疾病的發生發展過程中發揮重要作用。然而,Nampt在心肌細胞肥大中的作用國內外少有報道,值得進一步研究。本實驗以新生大鼠心肌細胞作為主要研究對象,通過檢測過表達Nampt對心肌細胞肥大的影響,并初步探索NF-κB依賴的信號通路在其中的作用。
1 材料與方法
1.1 主要材料 出生1~3 d的SD大鼠,♀♂不限,購自廣東省醫學實驗動物中心。DMEM高糖培養基、胰蛋白酶和胎牛血清均購自Gibco BRL公司;苯腎上腺素(PE)購自Sigma-Aldrich公司,用無菌超純水配成0.1 mol·L-1儲存液,并于-80℃保存;Nampt兔多抗購自Sigma-Aldrich公司;β-actin小鼠單抗、辣根過氧化物酶標記的羊抗兔二抗均購自廣州市潔利生物醫學有限公司;Opti-MEMI培養基和Lipofectamine 2000轉染試劑購自Invitrogen公司;含有NF-κB結合位點的PGL4.32-NF-κB-RE-luc2P和作為內參使用的PRL-TK質粒均購自Promega公司。
1.2 方法
1.2.1 心肌細胞培養與處理 取1~3 d新生SD大鼠,用75%酒精擦凈胸部及腹部皮膚后,開胸,取出心臟,放入預冷的PBS緩沖液中。剔除心臟上的結締組織,并剪成1 mm3的碎塊。連同PBS吸入離心管,瞬時離心后去上清。將心肌組織移入錐形瓶,加入15 mL 0.08%的胰蛋白酶于磁力攪拌器上37℃水浴消化。5 min后收集細胞上清至5 mL含10%胎牛血清的DMEM培養液中,1000 r·min-1離心5 min,棄上清,重懸細胞沉淀,剩余組織繼續消化,直到組織塊消化完全為止。收集各次消化細胞懸液,1000 r·min-1離心5 min,用DMEM培養基重懸細胞,接種于培養瓶中,置于37℃,5%CO2培養箱中孵育1 h后,收集上清,并加入0.1 mmol·L-1Brdu以抑制心肌成纖維細胞生長。培養24h之后,換新鮮含10%胎牛血清的DMEM高糖培養基繼續培養,此時部分細胞開始出現搏動。在處理細胞之前換含0.1%胎牛血清的DMEM培養基處理細胞24 h,使細胞同步化[8]。再給予PE(100 μmol·L-1)或轉染Nampt及空載體處理細胞24 h,并進行后續實驗。
1.2.2 Real-time PCR檢測mRNA表達 乳鼠心肌細胞給予不同濃度PE處理不同時間或給予過表達Nampt及其空載體處理之后,棄培養基,采用TRIzol一步法提取心肌細胞中的總RNA,RNA 定量后逆轉錄合成cDNA,用BIO-RAD熒光定量PCR系統進行實時熒光定量PCR反應,反應條件為:95℃ 10 s;95℃ 5 s,60℃ 30 s,循環50次;95℃ 15s,60℃ 1 h;65℃ 30 s,循環61次。每組樣品設3個復孔,以保證實驗數據的有效性。各目的基因及內參引物以Primer premier 5.0 軟件設計,由上海生工生物工程技術服務有限公司合成,引物序列見Tab 1。

Tab 1 Sequence of primers for quantitative real-time PCR
1.2.3 Western blot檢測蛋白表達 乳鼠心肌細胞分別過表達空載體質粒或EGFP-Nampt質粒24 h之后,棄培養基,加入三去污裂解液提取細胞總蛋白,依次進行 SDS-PAGE凝膠電泳、轉膜、封閉,并加入待測一抗Nampt(稀釋比: 1 ∶1 000)和內參抗體β-actin(稀釋比:1 ∶1 500),于4 ℃孵育過夜,次日加入相應的二抗,化學發光法檢測目的蛋白表達水平,采用Image J軟件對顯影的條帶進行灰度分析。
1.2.4 免疫熒光檢測心肌細胞表面積 在細胞培養皿中放入玻片,將心肌細胞接種在玻片上,心肌細胞分別給予PE(100 μmol·L-1)處理或過表達Nampt 24 h之后,棄培養基,冷PBS緩沖液洗3次,并用4%多聚甲醛室溫固定15~30 min,在PBS緩沖液中加入0.1%羅丹明標記的鬼筆環肽(Rhodamine-phalloidin)和0.1% saponin,充分混勻后加入培養皿中,室溫靜置1 h(避光);用含有0.1% saponin的PBS緩沖液洗細胞3次,每次5 min,在載玻片上滴加一小滴含DAPI的封片劑,取出蓋玻片,并吸干水分使其倒扣在載玻片上,當封片劑分布均勻后,在蓋玻片四周涂上指甲油,4℃過夜,用激光共聚焦顯微鏡對其進行觀察,每次實驗過程中,各組隨機選取7個視野,每個視野包含5~10個心肌細胞,該實驗重復3次,測量35~70個細胞的表面積,利用Image J圖像分析軟件計算細胞表面積,并分析計算結果。1.2.5 雙熒光素酶報告基因檢測NF-κB依賴的轉錄活性 心肌細胞按5×104/cm2密度接種于96孔板中,使用Lipofectamine 2000轉染PGL4.32-NF-κB-RE-luc2P質粒和作為內參使用的PRL-TK載體,細胞于培養箱中繼續孵育6 h后換入新鮮的含10%胎牛血清的DMEM培養基培養12 h。按照Promega公司Dual-Luciferase Reporter Assay System說明書進行操作,加入 luciferase reagent充分混合均勻后,將待測液轉移入白色不透光的96孔板中,用Varioskan Flash型全波長掃描式多功能讀數儀(美國Thermo公司)測量可見光強度10 s(此時的熒光為PGL4.32-NF-κB-RE-luc2P載體轉錄翻譯產生的螢火蟲luciferase發出),在96孔板中加入Stop&Glo Reagent,混合均勻,用Varioskan Flash型全波長掃描式多功能讀數儀進一步測量可見光強度10 s(此時的熒光為內參載體PRL-TK轉錄翻譯產生的海腎luciferase發出),將所得的數值,以海腎luciferase為參照進行標準化,計算均值,統計分析。
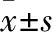
2 結果
2.1 過表達Nampt對Nampt mRNA和蛋白水平的影響 為了驗證過表達EGFP-Nampt是否成功,我們分別利用Real-time PCR和Western blot的方法檢測轉染EGFP-Nampt和空載體質粒之后Nampt的mRNA和蛋白表達變化。如Fig 1所示,與轉染空載體組(Vector)相比,EGFP-Nampt組Nampt的mRNA和蛋白水平明顯升高。
2.2 過表達Nampt對肥大基因表達的影響 為了探討過表達Nampt對心肌細胞肥大的影響,我們在原代培養的新生大鼠心肌細胞中分別轉染了EGFP-Nampt和空載體質粒或給予PE(100 μmol·L-1)處理,24 h之后分別檢測了ANP和BNP的表達情況。如Fig 2所示,我們發現PE處理心肌細胞24 h可明顯增加ANP和BNP的表達,而過表達EGFP-Nampt能夠模擬PE的作用誘導心肌細胞肥大,過表達空載體質粒則無此效應。
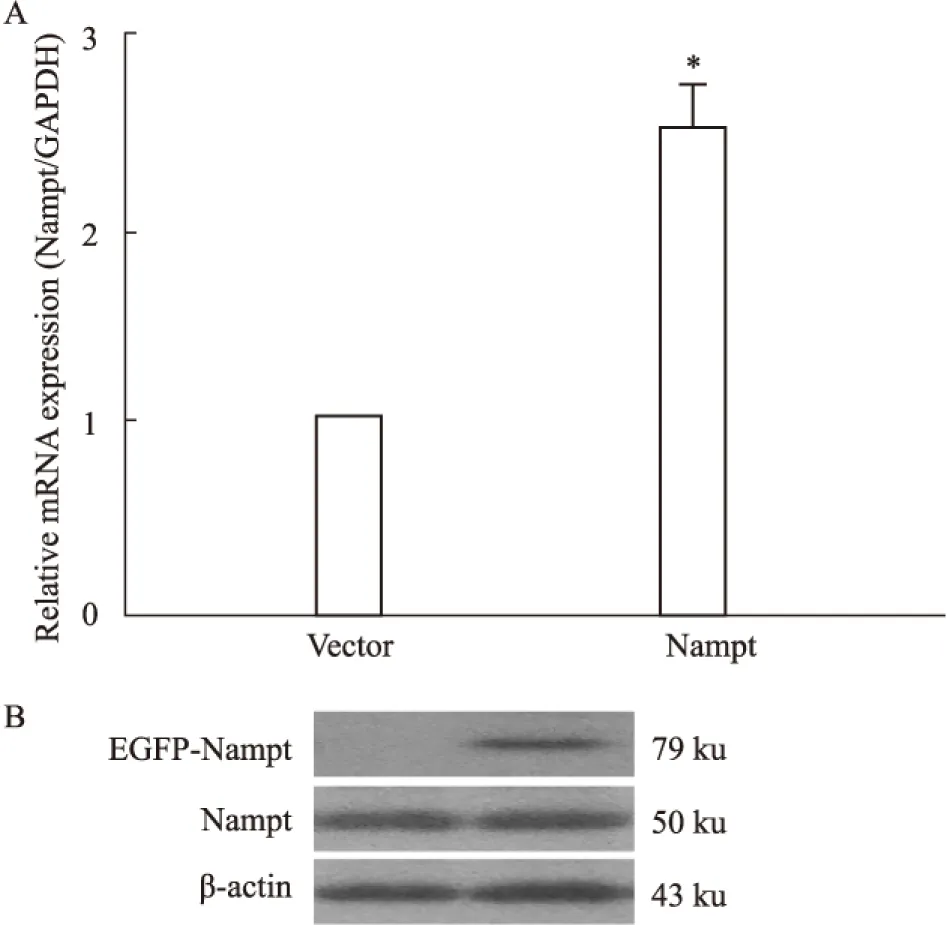
Fig 1 EGFP-Nampt transiently
Cardiomyocytes were transfected with Nampt. A: the mRNA expression of Nampt was detected by Real-time PCR; B: the protein expression of Nampt was measured by Western blot.*P<0.05vsvector group
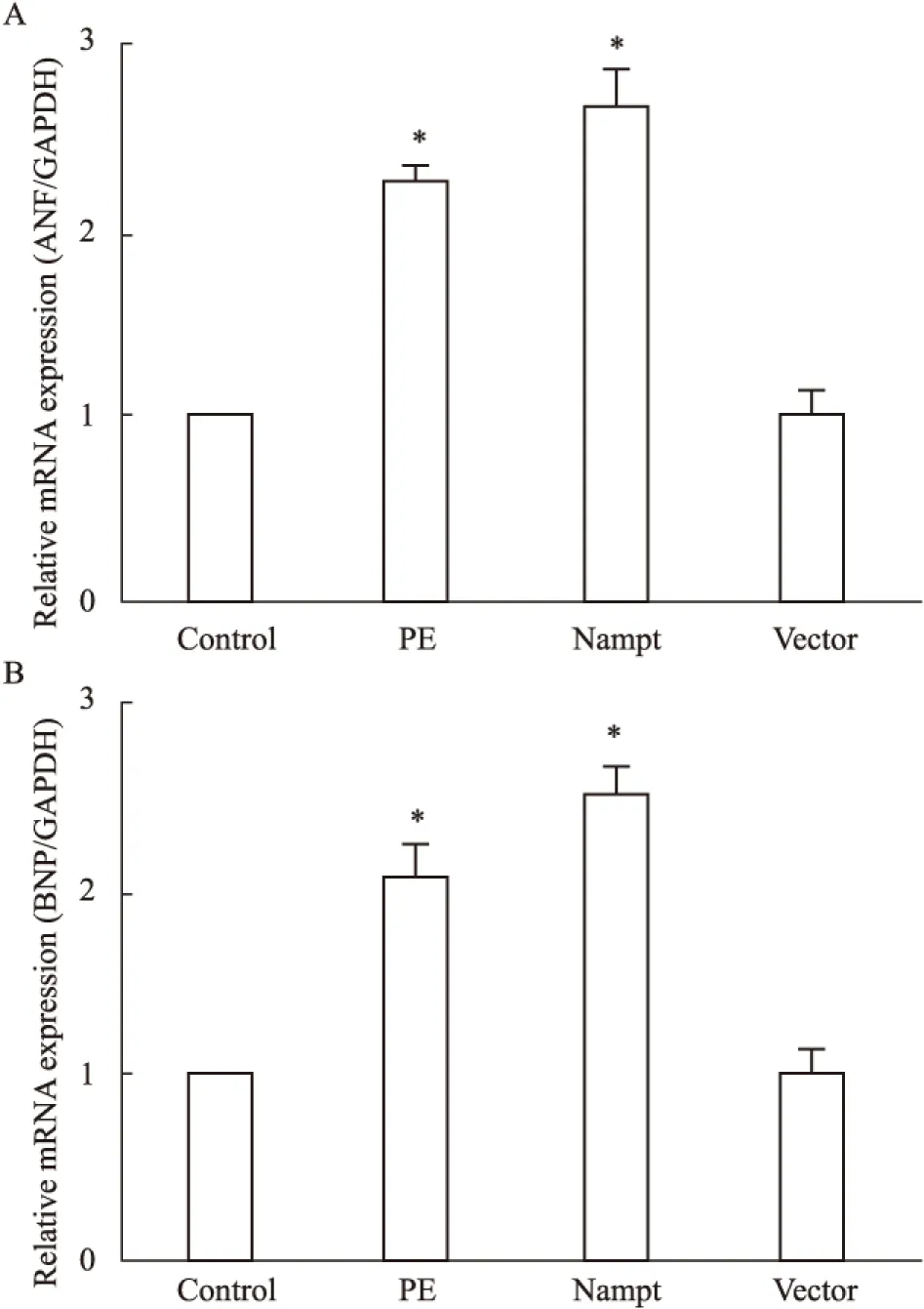
Fig 2 The mRNA expression of ANP and BNP in
The cardiomyocytes were transfected with EGFP-Nampt or treated with 100 μmol·L-1PE for 24 h. A,B: the mRNA expression of ANP and BNP were detected by Real-time PCR.*P<0.05vscontrol group
2.3 過表達Nampt對心肌細胞表面積的影響 為了進一步明確Nampt在心肌細胞肥大中的功能,我們在轉染了EGFP-Nampt和空載體質粒或給予PE處理24 h之后進一步檢測了心肌細胞表面積。如Fig 3所示,在心肌細胞中PE可明顯增大心肌細胞表面積,而過表達EGFP-Nampt能夠模擬PE的作用增加心肌細胞表面積,過表達空載體質粒則無此效應。
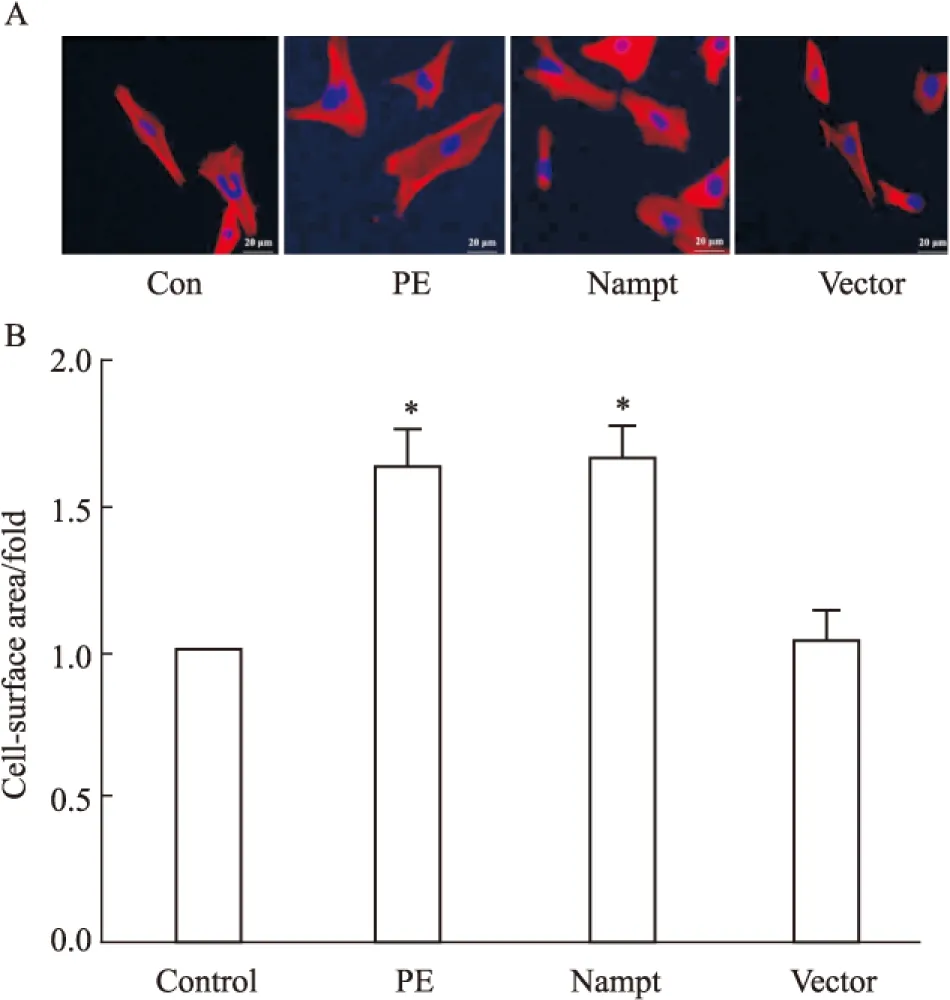
Fig 3 The cell-surface area in cardiomyocytes transfected with EGFP-Nampt
The cardiomyocytes were transfected with EGFP-Nampt or treated with 100 μmol·L-1PE for 24 h. A-B: the cell-surface area was measured.*P<0.05vscontrol group
2.4 過表達Nampt對NF-κB依賴的轉錄活性的影響 NF-κB作為一個重要的轉錄因子,激活后能夠調控下游與心肌肥大相關靶基因的轉錄[9]。為了研究過表達Nampt誘導心肌細胞肥大的分子機制,我們將含有NF-κB結合位點的PGL4.32-NF-κB-RE-luc2P質粒和內參質粒PRL-TK共轉染入心肌細胞,如Fig 4所示,給予心肌細胞PE處理24 h后NF-κB依賴性轉錄活性明顯增加,過表達EGFP-Nampt質粒也能夠增加NF-κB依賴性轉錄活性,而過表達空載體質粒則無此效應。
2.5 沉默NF-κB對過表達Nampt所致心肌細胞肥大的影響 為了進一步探討NF-κB在Nampt調控心肌肥大中的作用,我們在心肌細胞中,檢測特異性沉默NF-κB的p65亞基之后,過表達Nampt對肥大基因表達和心肌細胞表面積的影響。結果如Fig 5所示,在心肌細胞中沉默NF-κB的p65亞基能夠取消Nampt對肥大基因和心肌細胞表面積的影響。
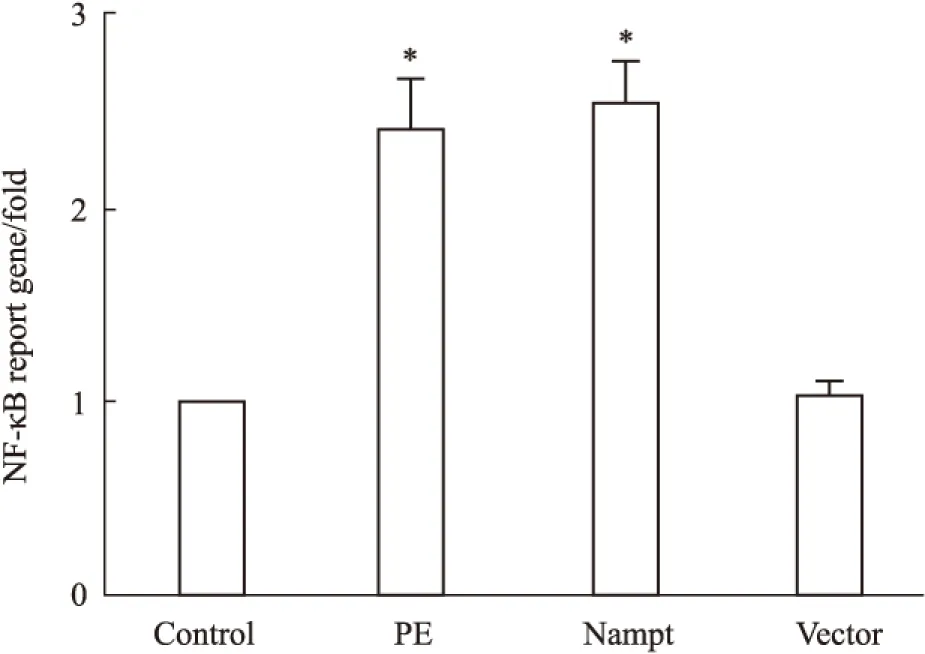
Fig 4 The NF-κB-dependent transcription activity in
The cardiomyocytes were transfected with EGFP-Nampt or treated with 100 μmol·L-1PE for 24 h. The NF-κB-dependent transcription activity was measured by luciferase reporter gene assays.*P<0.05 vs control group
3 討論
Nampt是NAD生物合成的限速酶,研究表明Nampt具有促血管新生、抗細胞凋亡、參與機體炎癥應答、促進多種細胞增殖分化成熟等生理功能, 對許多疾病如糖尿病、癌癥、自身免疫性疾病等有潛在的作用[10-11]。近年來,大量研究證明Nampt與心血管疾病的發生發展密切相關。在冠狀動脈不穩定粥樣硬化患者的頸動脈斑塊和急性心肌梗死病人破裂斑塊中Nampt表達明顯增加[11-12]。研究發現Nampt能夠增強MMP-2和MMP-9的酶活性,而MMP可以降解細胞外基質,使纖維帽變薄,導致斑塊破裂,提示Nampt在動脈粥樣硬化斑塊不穩定中起重要作用[13]。此外,Nampt抑制劑APO866治療心肌梗死可以減少梗死面積、中性粒細胞浸潤和在心肌缺血再灌注損傷中小鼠模型體內活性氧的產生[14]。因此,Nampt 藥理抑制劑作為一種有效的治療藥物,可以減少心肌梗死氧化介導的組織損傷。柴雅琴等[15]的研究還發現Nampt在妊娠高血壓綜合征產婦胎盤組織中高表達,且二者的表達水平呈明顯正相關,提示其與妊娠期高血壓密切相關。Schutte 等[16]認為Nampt可能通過炎癥反應、血管收縮等作用于心血管系統疾病。以上結果提示Nampt對動脈粥樣硬化、心肌梗死、妊娠高血壓等心血管疾病具有重要的調節作用,但Nampt能否影響心肌細胞肥大,目前國內外少有報道。在本課題中,我們發現過表達Nampt能夠模擬PE的作用誘導心肌細胞肥大,提示Nampt可能是一個潛在的防治心肌肥大的藥物作用靶點。
Nampt調控心肌細胞肥大的分子機制尚不清楚,值得進一步研究。臨床研究顯示Nampt的濃度在慢性炎癥性疾病中明顯增加[17],提示Nampt與炎癥反應密切相關。NF-κB是細胞內非常重要的核轉錄因子,不僅直接調控參與免疫炎癥相關基因的表達,而且也調節與促炎性細胞因子、趨化因子的產生、細胞增殖和細胞凋亡相關基因的轉錄[18]。近年來的研究提示,心肌細胞肥大的過程中需要NF-κB的激活,抑制心肌細胞肥大性反應與降低NF-κB活性呈正相關[19-20]。在心肌組織中,NF-κB可被多種引起心肌肥厚的刺激因子活化引起心肌細胞肥大,它的過表達還可啟動 ANP、BNP的表達[21]。此外,NF-κB還可以與MAPK等其他導致肥厚的信號轉導通路之間交互作用,共同促進心肌肥厚的發生發展[22]。以上結果提示NF-κB在心肌肥大的發生發展過程中具有重要作用。那么NF-κB是否參與了Nampt對心肌肥大的調控作用呢?為了解決這個問題,我們檢測了過表達Nampt對NF-κB依賴性轉錄活性的影響,發現過表達Nampt能夠增加NF-κB依賴性轉錄活性,進一步的研究表明沉默NF-κB的p65亞基能夠明顯逆轉過表達Nampt誘導心肌肥大的效應,提示NF-κB信號通路參與了Nampt對心肌細胞肥大的調控作用。然而,在本文中我們也發現沉默NF-κB并不能完全阻斷Nampt的作用,Nampt還可能通過其它途徑發揮調控心肌細胞肥大的作用。研究表明作為NAD合成的關鍵酶,Nampt與酶活性依賴于NAD的沉默信息調節因子2(silent information regulator 2, Sir2)家族成員關系密切,Nampt可通過調控NAD的合成影響SIRT1的活性進而調節SIRT1下游通路[23]。此外,Nampt還被報道參與調控AKT及MAPK等信號通路[24],這些通路在Nampt調控心肌肥大中的作用尚需我們進一步探討。
綜上所述,在本課題中我們發現過表達Nampt能夠模擬PE的作用誘導心肌細胞肥大,且其機制與NF-κB依賴性轉錄激活有關。今后的工作中,我們將通過腺病毒心室壁注射的方式在整體動物水平干預Nampt,進一步在體內明確Nampt對心肌肥大的調控作用,并深入探索Nampt調控NF-κB的具體分子機制以及Nampt調控心肌肥大的其它信號通路,為確定心肌肥大治療的新靶點提供初步的實驗數據。
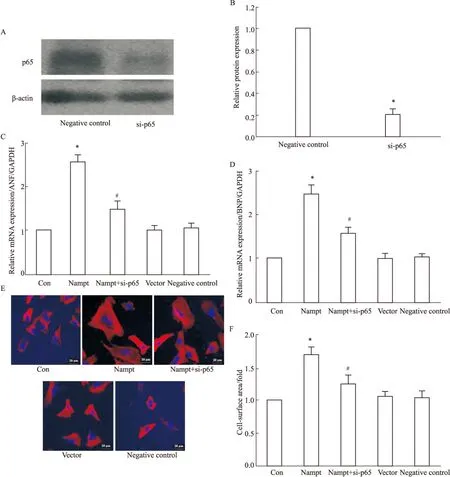
Cardiomyocytes overexpressed with Nampt after transfection with si-p65 were treated with PE for 24 h. A-B: the protein expression of p65 was measured by Western blot; C-D: the mRNA expression of ANP and BNP were detected by Real-time PCR; E-F: the cell-surface area was measured.*P<0.05 vs control group,#P<0.05vsNampt group.
(致謝:本實驗在廣州醫科大學藥物研發中心和醫學實驗中心完成,非常感謝藥物研發中心和醫學實驗中心的各位老師在實驗技術上的幫助。)
[1] Cai Y, Yu S S, Chen S R, et al. Nmnat2 protects cardiomyocytes from hypertrophy via activation of SIRT6[J].FEBSLett, 2012, 586(6): 866-74.
[2] Tsuruda T, Sekita-Hatakeyama Y, Hao Y, et al. Angiotensin Ⅱ Stimulation of Cardiac Hypertrophy and Functional Decompensation in Osteoprotegerin-Deficient Mice[J].Hypertension, 2016,67(5): 848-56.
[3] 黃 波,江 芬,薛 萊,等. PPARβ在虎杖苷抗高糖高胰島素誘導心肌肥大中的作用[J].中國藥理學通報,2015, 31(9):1264-9.
[3] Huang B, Jiang F, Xue L, et al. PPARβ-related signal pathway involves in the effect of polydatin on high glucose-and insulin-induced cardiomyocyte hypertrophy[J].ChinPharmacolBull, 2015, 31(9): 1264-9.
[4] Chen H, Wang S, Zhang H, et al. Nicotinamide phosphoribosyltransferase (Nampt) in carcinogenesis: new clinical opportunities[J].ExpertRevAnticancerTher, 2016,16(8): 827-8.
[5] Wang P, Li W L, Liu J M, et al. NAMPT and NAMPT-controlled NAD Metabolism in Vascular Repair[J].JCardiovascPharmacol, 2016,67(6): 474-81.
[6] Li S, Wang C, Li K, et al. NAMPT knockdown attenuates atherosclerosis and promotes reverse cholesterol transport in ApoE KO mice with high-fat-induced insulin resistance[J].SciRep, 2016, 6: 26746.
[7] Luizon M R, Palei A C, Belo V A, et al. Gene-gene interactions in the NAMPT pathway, plasma visfatin/NAMPT levels, and antihypertensive therapy responsiveness in hypertensive disorders of pregnancy[J].PharmacogenomicsJ, 2016.
[8] Geng B, Cai Y, Gao S, et al. PARP-2 knockdown protects cardiomyocytes from hypertrophy via activation of SIRT1[J].BiochemBiophysResCommun, 2013, 430(3): 944-50.
[9] Yuan Z, Matias F B, Wu J, et al. Koumine attenuates lipopolysaccaride-stimulated inflammation in RAW264.7 macrophages, coincidentally associated with inhibition of NF-kappaB, ERK and p38 pathways[J].IntJMolSci, 2016, 17(3): 430.
[10] Ju H Q, Zhuang Z N, Li H, et al. Regulation of the Nampt-mediated NAD salvage pathway and its therapeutic implications in pancreatic cancer[J].CancerLett, 2016, 379(1): 1-11.
[11] Kadoglou N P, Sailer N, Moumtzouoglou A, et al. Visfatin (nampt) and ghrelin as novel markers of carotid atherosclerosis in patients with type 2 diabetes[J].ExpClinEndocrinolDiabetes, 2010, 118(2): 75-80.
[12] Kadoglou N P, Sailer N, Kapelouzou A, et al. Effects of atorvastatin on apelin, visfatin (nampt), ghrelin and early carotid atherosclerosis in patients with type 2 diabetes[J].ActaDiabetol, 2012, 49(4): 269-76.
[13] 黃芬,熊曉昉,游莎,等. 內脂素經核因子κB途徑誘導人單核細胞基質金屬蛋白酶-2和9表達的實驗研究[J]. 中華心血管病雜志,2010, 38(5): 455-9.
[13] Huang F, Xiong X F, You S, et al. Visfatin upregulates MMP-2 and MMP-9 expresstom in human monocytes through activating NF-κB[J].ChinJCardiol, 2010, 38(5): 455-9.
[14] Montecucco F, Bauer I, Braunersreuther V, et al. Inhibition of nicotinamide phosphoribosyltransferase reduces neutrophil-mediated injury in myocardial infarction[J].AntioxidRedoxSignal, 2013,18(6): 630-41.
[15] 柴雅琴,楊艷花,黃桂香. MCP-1和Visfantin/PBEF在妊高癥患者胎盤中的表達及意義[J]. 醫學信息,2014, 27(4):126-7.
[15] Chai Y Q,Yang Y H,Huang G X. Expression and significance of MCP-1and Visfantin/PBEF in the placenta of pregnancy-induce hypertension syndreome patients[J].MedInform,2014, 27(4):126-7.
[16] Schutte A E, Huisman H W, Schutte R, et al. Adipokines and cardiometabolic function: How are they interlinked?[J].RegulPept, 2010, 164(2-3): 133-8.
[17] Yarar-Fisher C, Bickel C S, Kelly N A, et al. Heightened TWEAK-NF-kappaB signaling and inflammation-associated fibrosis in paralyzed muscles of men with chronic spinal cord injury[J].AmJPhysiolEndocrinolMetab, 2016,310(9): E754-61.
[18] Jones W K, Brown M, Wilhide M, et al. NF-kappaB in cardiovascular disease: diverse and specific effects of a “general” transcription factor?[J].CardiovascToxicol, 2005,5(2): 183-202.
[19] Zelarayan L, Renger A, Noack C, et al. NF-kappaB activation is required for adaptive cardiac hypertrophy[J].CardiovascRes, 2009,84(3): 416-24.
[20] Gupta S, Young D, Maitra R K, et al. Prevention of cardiac hypertrophy and heart failure by silencing of NF-kappaB[J].JMolBiol, 2008, 375(3): 637-49.
[21] Gaspar-Pereira S, Fullard N, Townsend PA, et al. The NF-kappaB subunit c-Rel stimulates cardiac hypertrophy and fibrosis[J].AmJPathol, 2012,180(3): 929-39.
[22] Chen G, Pan SQ, Shen C, et al. Puerarin inhibits angiotensin Ⅱ-induced cardiac hypertrophy via the redox-sensitive ERK1/2, p38 and NF-kappaB pathways[J].ActaPharmacolSin, 2014, 35(4): 463-75.
[23] Ming G F, Wu K, Hu K, et al. NAMPT regulates senescence, proliferation, and migration of endothelial progenitor cells through the SIRT1 AS lncRNA/miR-22/SIRT1 pathway[J].BiochemBiophysResCommun, 2016,478(3): 1382-8.
[24] Yu X Y, Qiao S B, Guan H S, et al. Effects of visfatin on proliferation and collagen synthesis in rat cardiac fibroblasts[J].HormMetabRes, 2010,42(7): 507-13.
Overexpression of Nampt induced cardiac hypertrophy by activating NF-κB
CAI Yi, HUANG Jun-jun, LIU Xia-wen, HUANG Bi-yun, ZHU Liu ,WU Bo
(CenterofPharmaceuticalResearchandDevelopment,GuangzhouMedicalUniversity,Guangzhou511436,China)
Aim To examine the effect of Nampt overexpression on cardiac hypertrophy, and elucidate the role of NF-κB. Methods The cultured neonatal cardiomyocytes were pretreated with 100 μmol·L-1PE or transfected with Nampt. The mRNA and protein expression of Nampt were determined by Real-time PCR and Western blot respectively. The cardiomyocyte hypertrophy was monitored by measuring cell-surface area and the mRNA levels of ANP and BNP, which were biomarkers of hypertrophic response. Moreover, we te- sted the effects of Nampt on NF-κB-dependent transcription activity through luciferase reporter gene assays. Results Nampt overexpression significantly increased cardiomyocyte surface area and the mRNA expression of ANP and BNP. In addition, Nampt overexpression could markedly increase NF-κB-dependent transcription activity. Moreover, when p65 was knocked down, cardiomyocytes with Nampt overexpression could not induce cardiac hypertrophy. Conclusion Overexpression of Nampt induces cardiac hypertrophy by increasing NF-κB-dependent transcription activity.
Nampt; cardiac hypertrophy; NF-κB; PE; ANP; BNP
時間:2016-12-27 16:13
http://www.cnki.net/kcms/detail/34.1086.R.20161227.1613.014.html
2016-09-05,
2016-11-15
國家自然科學基金資助項目(No 81300085);廣州市科技計劃-科學研究專項(No 201510010179);廣東省科技計劃項目-廣東省高科技發展專項項目(No 2013B010403024);廣州市屬高校科研計劃項目(No 1201430926,1201410511)
蔡軼(1984-),男,博士,助理研究員,研究方向:心血管藥理學,Tel:020-37104150,E-mail:yicaisysu@163.com; 吳 波(1971-),男,博士,研究員,研究方向:天然藥物化學,通訊作者,Tel:020-37103258,E-mail:gykyc@163.com
10.3969/j.issn.1001-1978.2017.01.007
A
1001-1978(2017)01-0033-07
R-332;R322.11;R345.54;R542.2;R977.3;R977.6
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48