大鱗副泥鰍生長(zhǎng)優(yōu)秀家系篩選以及基因型對(duì)生長(zhǎng)性能的影響
2017-02-16 01:56:52潘文賢李彩娟王國(guó)成葉竹青凌去非
水產(chǎn)養(yǎng)殖 2017年1期
關(guān)鍵詞:優(yōu)勢(shì)生長(zhǎng)
潘文賢,李彩娟,王國(guó)成,葉竹青,凌去非
(1.蘇州大學(xué)水產(chǎn)研究所,江蘇蘇州 215123;2..常熟市水產(chǎn)技術(shù)推廣站,江蘇常熟 215500)
大鱗副泥鰍生長(zhǎng)優(yōu)秀家系篩選以及基因型對(duì)生長(zhǎng)性能的影響
潘文賢1,2,李彩娟1,王國(guó)成1,葉竹青1,凌去非1
(1.蘇州大學(xué)水產(chǎn)研究所,江蘇蘇州 215123;2..常熟市水產(chǎn)技術(shù)推廣站,江蘇常熟 215500)

本試驗(yàn)構(gòu)建了11個(gè)大鱗副泥鰍家系,并進(jìn)行了生長(zhǎng)性能優(yōu)秀家系的篩選,利用生長(zhǎng)相關(guān)SSR標(biāo)記對(duì)各家系進(jìn)行生長(zhǎng)性狀與基因型相關(guān)分析,探究不同基因型對(duì)生長(zhǎng)性能的影響。結(jié)果顯示AB2013F-7、AB2013F-89、AB2013F-45等家系的體質(zhì)量、體長(zhǎng)和全長(zhǎng)均值顯著(P<0.05)大于其他家系,其中AB2013F-7家系各生長(zhǎng)指標(biāo)均值最大;以上三個(gè)家系117 d內(nèi)的絕對(duì)增重率也表現(xiàn)最好,分別為0.068、0.051和0.048 g/d。對(duì)各家系基因型進(jìn)行分析,顯示各家系所擁有優(yōu)勢(shì)基因型的數(shù)量與生長(zhǎng)性狀均值大小成正相關(guān),含生長(zhǎng)相關(guān)標(biāo)記優(yōu)勢(shì)基因型的數(shù)量越多,家系的生長(zhǎng)性能表現(xiàn)越優(yōu)秀。
大鱗副泥鰍;家系;生長(zhǎng);基因型;微衛(wèi)星
家系選擇(family selection)是一種選擇育種策略,即通過(guò)建立家系,對(duì)家系后代的經(jīng)濟(jì)性狀進(jìn)行有目的的選擇和比較分析,是水產(chǎn)動(dòng)物種質(zhì)改良的重要途徑之一[1]。然而傳統(tǒng)的家系選育育種周期長(zhǎng),耗時(shí)耗力,需要經(jīng)歷十多代的交配繁殖來(lái)獲得相關(guān)純系或純種[2-3],通過(guò)分子標(biāo)記輔助選擇則可縮短育種進(jìn)程。目前,國(guó)內(nèi)外學(xué)者對(duì)牙鲆[4]、鮑[5]、馬氏珠母貝[6]等進(jìn)行了分子標(biāo)記輔助育種相關(guān)研究,改善了相關(guān)水產(chǎn)動(dòng)物抗病性以及生長(zhǎng)等性狀。
大鱗副泥鰍(Paramisgurnus dabryanus Sauvage)是鰍科小型重要經(jīng)濟(jì)魚(yú)類,也是近年來(lái)養(yǎng)殖規(guī)模增長(zhǎng)較大的魚(yú)類之一,其味道鮮美,營(yíng)養(yǎng)價(jià)值較高,在國(guó)內(nèi)外市場(chǎng)也廣受歡迎。目前分子標(biāo)記輔助家系選育工作在相關(guān)水產(chǎn)動(dòng)物中得到了一定的應(yīng)用[7-9],但國(guó)內(nèi)外均未見(jiàn)大鱗副泥鰍家系選育的相關(guān)報(bào)道。本研究通過(guò)建立大鱗副泥鰍家系,并對(duì)家系父母本基因型進(jìn)行分析,預(yù)測(cè)子代基因型情況,探討優(yōu)勢(shì)基因型與家系生長(zhǎng)性狀的關(guān)系,為大鱗副泥鰍分子標(biāo)記輔助選育奠定基礎(chǔ)。
1 材料與方法
1.1 家系構(gòu)建
試驗(yàn)用大鱗副泥鰍由蘇州奧博生物科技有限公司泥鰍養(yǎng)殖基地提供。2013年6月6日挑選來(lái)自洪澤湖的野生大鱗副泥鰍親本11組,按照雌雄配比為1∶1進(jìn)行人工催產(chǎn),各組親本分別放于獨(dú)立水族箱中產(chǎn)卵,產(chǎn)卵后,撈出各家系父母本,剪鰭條保存于95%的乙醇中備用。1 d后魚(yú)苗孵化出膜,2 d后平游,投喂開(kāi)口餌料。10 d后將11家系移至設(shè)置在同一池塘的11個(gè)網(wǎng)箱中進(jìn)行培育,網(wǎng)箱規(guī)格為2 m×1m×1 m,網(wǎng)箱網(wǎng)目為80目,每個(gè)家系密度控制在1 000尾,培育期間,保持各家系的生長(zhǎng)環(huán)境盡可能相同。于2013年7月21日再次對(duì)試驗(yàn)網(wǎng)箱中各家系密度進(jìn)行控制,每個(gè)家系密度為55尾,并測(cè)量各家系的平均初始體質(zhì)量。2013年11月15日,每個(gè)家系撈取30尾,進(jìn)行生長(zhǎng)指標(biāo)(體長(zhǎng)、體質(zhì)量和全長(zhǎng))測(cè)量。
1.2 父母本基因組DNA提取
60尾大鱗副泥鰍基因組DNA的提取參照標(biāo)準(zhǔn)酚-氯仿抽提程序進(jìn)行。紫外分光光度計(jì)測(cè)定DNA樣品的濃度和純度,瓊脂糖凝膠電泳測(cè)定DNA基因組的完整性,稀釋DNA至50~100 ng/μL,-20℃保存?zhèn)溆谩?/p>
1.3 引物合成和PCR反應(yīng)
采用的微衛(wèi)星標(biāo)記由本實(shí)驗(yàn)室第二代測(cè)序技術(shù)自行開(kāi)發(fā)獲得[10],由上海生工生物工程有限公司合成引物,11個(gè)標(biāo)記是(表1)本實(shí)驗(yàn)室前期已篩選的大鱗副泥鰍生長(zhǎng)相關(guān)微衛(wèi)星標(biāo)記。PCR擴(kuò)增采用20μL體系:2×Mix 10μL,H2O 7μL,DNA 1μL,上下游引物各1μL(10μM)。整個(gè)反應(yīng)28個(gè)循環(huán),94℃變性30 s,退火30 s,72℃延伸30 s,各對(duì)引物的特異退火溫度見(jiàn)表1。首次循環(huán)前先預(yù)變性3 min,最后一次循環(huán)結(jié)束后72℃延伸5 min。
1.4 電泳及染色
PCR擴(kuò)增產(chǎn)物用6%非變性聚丙烯酰胺凝膠電泳分離,電泳緩沖液為0.5×TBE,上樣1μL,150 V電泳80 min。參照改良的銀染方法檢測(cè)微衛(wèi)星帶型[10],染色10min,水洗2次(1 min/次),顯色液預(yù)冷,顯帶后水洗;采用凝膠成像系統(tǒng)(BIO-RAD)拍照并分析帶型。
1.5 數(shù)據(jù)處理

表1 11對(duì)大鱗副泥鰍微衛(wèi)星引物信息
采用SPSS 20.0對(duì)11個(gè)大鱗副泥鰍家系體質(zhì)量、體長(zhǎng)、全長(zhǎng)均值進(jìn)行單因素方差分析(One way ANOVA),利用最小顯著極差法(least significant difference,LSD)對(duì)各個(gè)家系間的測(cè)量數(shù)據(jù)進(jìn)行多重比較;計(jì)算各家系絕對(duì)增重率。
絕對(duì)增重率AGRW(g/d)=(W2-W1)/(T2-T1)式中W1、W2分別為T(mén)1和T2時(shí)的體質(zhì)量。
根據(jù)各家系父母本基因型,結(jié)合試驗(yàn)1中的各生長(zhǎng)相關(guān)SSR標(biāo)記的優(yōu)勢(shì)基因型,分析優(yōu)勢(shì)基因型對(duì)家系生長(zhǎng)性能的影響。
2 結(jié)果
2.1 大鱗副泥鰍不同家系間生長(zhǎng)性能比較
11個(gè)家系全長(zhǎng)、體長(zhǎng)和體質(zhì)量的平均值、標(biāo)準(zhǔn)差見(jiàn)表2。各家系全長(zhǎng)大小關(guān)系為:AB2013F-7>AB2013F-89>AB2013F-45>AB2013F-76>AB2013F-74>AB2013F-3>AB2013F-27>AB2013F-70>AB2013F-77>AB2013F-36>AB2013F-81;體長(zhǎng)大小關(guān)系為:AB2013F-7>AB2013F-89>AB2013F-45>AB2013F-76>AB2013F-74>AB2013F-3>AB2013F-27>AB2013F-77>AB2013F-70>AB2013F-81>AB2013F-36;體質(zhì)量大小關(guān)系為:AB2013F-7>AB2013F-89>AB2013F-45>AB2013F-76>AB2013F-3>AB2013F-27>AB2013F-74>AB2013F-81>AB2013F-70>AB2013F-77> AB2013F-36。由此可見(jiàn),AB2013F-7、AB2013F-89和AB2013F-45家系的全長(zhǎng)、體長(zhǎng)以及體質(zhì)量均值為所有家系中最高的。通過(guò)LSD多重比較發(fā)現(xiàn)AB2013F-7號(hào)家系全長(zhǎng)、體長(zhǎng)顯著(P<0.05)大于除AB2013F-89號(hào)家系外的其他所有家系,而其體質(zhì)量顯著(P<0.05)大于所有家系;AB2013F-89和AB2013F-45的全長(zhǎng)、體長(zhǎng)以及體質(zhì)量也與其他家系具有統(tǒng)計(jì)學(xué)意義差異(P<0.05)(見(jiàn)表2),其他家系之間各指標(biāo)無(wú)統(tǒng)計(jì)學(xué)意義差異。
同時(shí),本研究也采用絕對(duì)增重率來(lái)評(píng)價(jià)家系的生長(zhǎng)性能(見(jiàn)表2),AB2013F-7、AB2013F-89和AB2013F-45三個(gè)家系117 d內(nèi)的絕對(duì)增重率分別為0.068 g/d、0.051 g/d、0.048 g/d,為11個(gè)家系中最高的三個(gè),其中AB2013F-7號(hào)家系絕對(duì)增重率是AB2013F-36家系的2.8倍。綜合多重比較結(jié)果與絕對(duì)增重率計(jì)算結(jié)果發(fā)現(xiàn),家系A(chǔ)B2013F-7、AB2013F-89和AB2013F-45生長(zhǎng)速度顯著快于其他家系,可確定為生長(zhǎng)快速家系。
2.2 11個(gè)大鱗副泥鰍家系父母本基因分型
采用11個(gè)SSR引物對(duì)11個(gè)家系父母本進(jìn)行基因分型,并推測(cè)出了子代基因型。本研究中的優(yōu)勢(shì)基因型為前期已篩選到的優(yōu)勢(shì)基因型,基因型統(tǒng)計(jì)發(fā)現(xiàn)11個(gè)家系表現(xiàn)優(yōu)勢(shì)基因型的SSR標(biāo)記個(gè)數(shù)為1-6個(gè)不等(表3),其中AB2013F-3、AB2013F-7家系有6個(gè)標(biāo)記在子代中具備表現(xiàn)優(yōu)勢(shì)基因型的機(jī)會(huì),為子代優(yōu)勢(shì)基因型含量最多的兩個(gè)家系,其中AB2013F-7家系的表型值也是所有家系中最高的;表現(xiàn)優(yōu)勢(shì)基因型最少的家系為AB2013F-27家系,僅有1個(gè)標(biāo)記在子代中表現(xiàn)優(yōu)勢(shì)基因型,另外家系A(chǔ)B2013F-36、AB2013F-70、AB2013F-74產(chǎn)生優(yōu)勢(shì)基因型的標(biāo)記在3個(gè)以內(nèi),AB2013F-27、36、70和74這四個(gè)家系其產(chǎn)生優(yōu)勢(shì)基因型數(shù)量比3個(gè)優(yōu)秀家系的優(yōu)勢(shì)基因型數(shù)量低,且這4個(gè)家系各生長(zhǎng)指標(biāo)均低于3個(gè)優(yōu)秀家系。但是結(jié)果中標(biāo)記數(shù)量多少與生長(zhǎng)性能的好壞并不是一一對(duì)應(yīng)的,可能是主效基因型與微效基因型對(duì)生長(zhǎng)影響不同造成的,也可能是父母本基因型在子代中因分離組合導(dǎo)致產(chǎn)生特定基因型個(gè)體的概率不同而產(chǎn)生的結(jié)果。
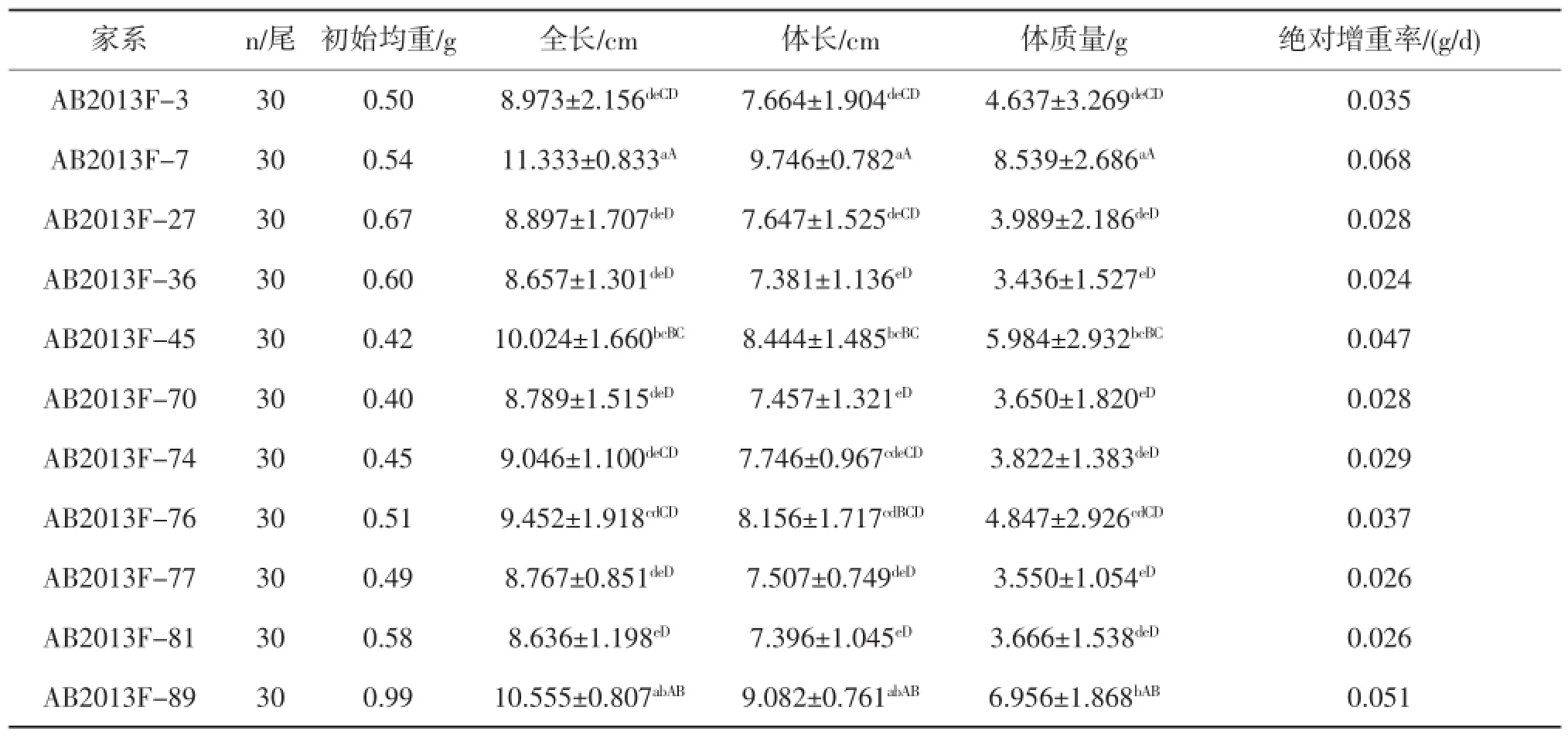
表2 11個(gè)大鱗副泥鰍家系絕對(duì)增重率以及生長(zhǎng)性能比較
3 討論
3.1 優(yōu)秀家系選擇
相關(guān)學(xué)者認(rèn)為魚(yú)類生活史的早期階段生長(zhǎng)速度最快,在此階段,魚(yú)類生長(zhǎng)易受到環(huán)境以及自身遺傳基因的影響,產(chǎn)生生長(zhǎng)上的差異[11]。部分學(xué)者對(duì)大鱗副泥鰍同科的泥鰍進(jìn)行了研究,認(rèn)為泥鰍在生長(zhǎng)初期遺傳差異造成表型差異表現(xiàn)較為顯著[12-13],故本研究采用了6月齡大鱗副泥鰍進(jìn)行表型值測(cè)定。本試驗(yàn)中各家系生存環(huán)境和飼喂條件均一致,但11個(gè)家系生長(zhǎng)差異較大,說(shuō)明各家系遺傳基礎(chǔ)的不同導(dǎo)致了生長(zhǎng)性能的差異。AB2013F-7、AB2013F-89和AB2013F-45三個(gè)家系生長(zhǎng)速度較快,可以作為進(jìn)一步選育的基礎(chǔ)群體。
3.2 各家系子代基因型效應(yīng)對(duì)生長(zhǎng)影響分析
分子標(biāo)記聚合育種是將幾個(gè)主效基因的優(yōu)勢(shì)基因型聚合到一個(gè)個(gè)體上的技術(shù)[14],優(yōu)勢(shì)基因型的聚合對(duì)于動(dòng)物品種性狀的改良與提升具有很大的促進(jìn)作用。相關(guān)學(xué)者在豬[15]產(chǎn)仔數(shù)、中國(guó)荷斯坦牛[16]產(chǎn)奶性狀的優(yōu)勢(shì)基因型聚合上進(jìn)行了研究,研究表明基因聚合能夠有效改良目標(biāo)性狀。徐磊[17]等采用8個(gè)生長(zhǎng)相關(guān)分子標(biāo)記進(jìn)行了生長(zhǎng)優(yōu)勢(shì)基因型的數(shù)量與大口黑鱸生長(zhǎng)速度相關(guān)性研究,發(fā)現(xiàn)生長(zhǎng)優(yōu)勢(shì)基因型的數(shù)量與生長(zhǎng)速度呈正相關(guān)。本研究發(fā)現(xiàn)各家系能表現(xiàn)優(yōu)勢(shì)基因型數(shù)量與生長(zhǎng)性狀均值大小有一定的關(guān)系,例如AB2013F-7家系生長(zhǎng)速度最快,其優(yōu)勢(shì)基因型數(shù)量也最多。但也存在例外情況,如AB2013F-3家系有6個(gè)優(yōu)勢(shì)基因型,比AB2013F-89要多,但生長(zhǎng)指標(biāo)卻低于后者,推測(cè)原因之一可能是相關(guān)標(biāo)記優(yōu)勢(shì)基因型在子代的表現(xiàn)概率大小不同,如在普通家系A(chǔ)B2013F-3家系中,Pda48的優(yōu)勢(shì)基因型BF表現(xiàn)概率為50%,而在優(yōu)秀家系A(chǔ)B2013F-89中為100%;還有可能是主效基因與微效基因相互作用的結(jié)果,主效基因與微效基因?qū)π誀畹呢暙I(xiàn)大小不同,導(dǎo)致表型值的大小與優(yōu)勢(shì)基因型數(shù)量不呈直線正相關(guān),例如優(yōu)秀家系A(chǔ)B2013F-89生長(zhǎng)相關(guān)標(biāo)記的數(shù)量?jī)H為4個(gè),但Pda48的BF基因型在此家系中表現(xiàn)概率為100%,為所有家系中最大概率,推測(cè)BF基因型對(duì)生長(zhǎng)指標(biāo)貢獻(xiàn)值較大的緣故。AB2013F-7家系優(yōu)勢(shì)基因型數(shù)量與AB2013F-3家系相同,但是標(biāo)記的種類和各標(biāo)記的表現(xiàn)概率不同,這可能是導(dǎo)致兩家系生長(zhǎng)狀況差異的原因。Pda261的優(yōu)勢(shì)基因型在AB2013F-7家系中表現(xiàn)概率為100%,在AB2013F-3家系中為50%,而Pda316的優(yōu)勢(shì)基因型在AB2013F-3家系中表現(xiàn)概率為100%,在AB2013F-7家系中為50%,但是AB2013F-7家系表型值顯著大于AB2013F-3家系,推測(cè)可能Pda261位點(diǎn)連鎖基因?qū)ιL(zhǎng)的貢獻(xiàn)值大于Pda316。
Pda247、Pda315在11個(gè)家系父母本中沒(méi)有相關(guān)優(yōu)勢(shì)等位基因,但可以通過(guò)在另外的群體(混合選育群體)中選擇具有優(yōu)勢(shì)等位基因的個(gè)體與家系個(gè)體交配,從而引入優(yōu)勢(shì)基因型。有的家系雖未表現(xiàn)優(yōu)勢(shì)基因型,但是卻含有組成優(yōu)秀基因型的等位基因,也具備育種潛力,如Pda371在普通家系A(chǔ)B2013F-3子代中為AB基因型,如果家系內(nèi)個(gè)體交配,則可產(chǎn)生含優(yōu)勢(shì)基因型AA個(gè)體。在非優(yōu)勢(shì)家系中,一些標(biāo)記也能產(chǎn)生優(yōu)勢(shì)基因型,優(yōu)勢(shì)家系中也不是全部標(biāo)記都可以產(chǎn)生優(yōu)勢(shì)基因型,主要因?yàn)樯L(zhǎng)相關(guān)標(biāo)記屬于數(shù)量性狀,其最終表現(xiàn)需要多基因的協(xié)同作用,主效基因與微效基因?qū)ιL(zhǎng)速度的影響效果差別較大,僅從有無(wú)優(yōu)勢(shì)等位基因及優(yōu)勢(shì)基因型的數(shù)量多少來(lái)判斷家系的好壞有一定的局限性,因此,后期研究應(yīng)重點(diǎn)加強(qiáng)主效基因標(biāo)記優(yōu)勢(shì)基因型的甄別和富集以進(jìn)一步加快良種選育進(jìn)程。
[1]吳仲慶.水產(chǎn)生物育種學(xué)[M].廈門(mén):廈門(mén)大學(xué)出版社. 2000.
[2]秦欽,邊文冀,蔡永祥,等.斑點(diǎn)叉尾鮰家系育種核心群生長(zhǎng)性能研究及優(yōu)良親本選擇[J].上海海洋大學(xué)學(xué)報(bào),2011(01):63-70.
[3]孫效文,魯翠云,賈智英,等.水產(chǎn)動(dòng)物分子育種研究進(jìn)展[J].中國(guó)水產(chǎn)科學(xué),2009(06):981-990.
[4]Fuji K,Hasegava O,Honda K,etal.Marker-assisted breeding of a lymphocystis disease-resistant Japanese flounder(Paralichthysolivaceus)[J].Aquaculture,2007(272):291-295.
[5]Hayes B,Baranski M,Goddard M E,et al.Optimisation of marker assisted selection for abalone breeding programs[J]. Aquaculture,2007(265):61-69.
[6]Qiu Y,Lu H,Zhu JT,et al.Characterization of novel ESTSSR markers and their correlations with growth and nacreous secretion traits in the pearl oysterPinctadamartensii(Dunker)[J].Aquaculture,2014(420-421):92-97.
[7]Grimholt U,Larsen S,Nordmo R,et al.MHC polymorphism and disease resistance in Atlantic salmon(Salmo salar);facing pathogenswith single expressed major histocompatibility class I and class II loci[J].Immunogenetics,2003,55(4):210-219.
[8]魯翠云,曹頂臣,孫效文,等.微衛(wèi)星分子標(biāo)記輔助鏡鯉家系構(gòu)建[J].中國(guó)水產(chǎn)科學(xué),2008(06):893-901.
[9]劉磊,李健,劉萍,等.三疣梭子蟹微衛(wèi)星標(biāo)記與生長(zhǎng)相關(guān)性狀的相關(guān)性分析[J].水產(chǎn)學(xué)報(bào),2012(07):1034-1041.
[10]李彩娟.基于第二代測(cè)序的大鱗副泥鰍微衛(wèi)星分子標(biāo)記的開(kāi)發(fā)與應(yīng)用[D].蘇州:蘇州大學(xué),2014.
[11]Ware D M.Power and evolutionary fitness of teleosts[J]. Canadian Journal of Fisheries and Aquatic Sciences,1982,39(1):3-13.
[12]王有基.泥鰍早期攝食節(jié)律及不同餌料和延遲投餌對(duì)其生長(zhǎng)和成活的影響[D].武漢:華中農(nóng)業(yè)大學(xué),2008.
[13]于連洋,凌去非,劉洋,等.48個(gè)泥鰍家系魚(yú)種階段的生長(zhǎng)比較[J].水生態(tài)學(xué)雜志,2010,3(5):62-68.
[14]孫效文.魚(yú)類分子育種學(xué)[M].北京:科學(xué)出版社,2010.
[15]陳克飛,李寧.豬FSHβ及ESR合并基因型對(duì)產(chǎn)仔數(shù)性狀的影響[J].科學(xué)通報(bào),2000,45(18):1963-1966.
[16]賈祥捷,王長(zhǎng)法,楊桂文,等.中國(guó)荷斯坦牛POU1F1基因與PRL基因的多態(tài)性及其聚合效應(yīng)對(duì)產(chǎn)奶性狀的影響[J].遺傳,2011,33(12):1359-1365.
[17]徐磊,白俊杰,李勝杰.大口黑鱸生長(zhǎng)性狀相關(guān)標(biāo)記的聚合效果分析[J].中國(guó)水產(chǎn)科學(xué),2014,21(1):53-58.
湖北荊門(mén)發(fā)布通知2017年年底前全部拆除全市養(yǎng)殖網(wǎng)箱
記者從2016年12月擬訂的《關(guān)于取締湖庫(kù)圍欄圍網(wǎng)網(wǎng)箱養(yǎng)殖的實(shí)施方案》上了解到,關(guān)于湖庫(kù)圍欄圍網(wǎng)網(wǎng)箱養(yǎng)殖的目標(biāo)為:通過(guò)規(guī)范整治,在荊門(mén)市境內(nèi)實(shí)現(xiàn)水域無(wú)珍珠養(yǎng)殖、湖庫(kù)無(wú)圍欄圍網(wǎng)(網(wǎng)箱)、水庫(kù)無(wú)投肥投餌養(yǎng)殖的“三無(wú)”目標(biāo)。到2016年12月底前全市拆圍達(dá)到30%以上,2017年12月底以前全面拆除完成;全市湖庫(kù)水質(zhì)明顯改善,飲用水源區(qū)湖庫(kù)水質(zhì)達(dá)到Ⅲ類標(biāo)準(zhǔn)以上。到2020年,所有湖庫(kù)水質(zhì)全部達(dá)標(biāo)。
根據(jù)方案,從現(xiàn)在起至2017年6月,全面啟動(dòng)拆圍工作。從2017年6月至8月開(kāi)展聯(lián)合整治。嚴(yán)格控制在湖庫(kù)集水區(qū)范圍內(nèi)新增污染物排放設(shè)施。對(duì)公共水域水面漁業(yè)養(yǎng)殖權(quán)和捕撈權(quán)對(duì)外承包,在2017年8月底前全部收回,并注銷《水域?yàn)┩筐B(yǎng)殖證》。對(duì)不按照規(guī)范時(shí)間拆除的,由公安、環(huán)保、水務(wù)、水產(chǎn)等具有執(zhí)法權(quán)的單位聯(lián)合予以強(qiáng)制拆除。從2017年9月至11月,對(duì)湖庫(kù)進(jìn)行全面巡查檢查,防止反彈,對(duì)已經(jīng)治理仍然進(jìn)行珍珠養(yǎng)殖、圍欄圍網(wǎng)(網(wǎng)箱)養(yǎng)殖的依法予以處罰,發(fā)現(xiàn)一起查處一起。
(www.bbwfish.com)
Screening fast-grow ing fam ilies and analysis of the effect of genotypes on grow th of Paramisgurnus dabryanus
Pan Wenxian1,LiCaijuan2,W ang Guocheng2,Ye Zhuqing2,Ling Qufei2
(1.Fisheries Institute of Soochow University,Suzhou 215123,China 2.Fisheries Technology Extension Station of Changshu city,Changshu 215500,China)
Eleven families of large-scale loach were established and fast-growing families were screened by comparing growth traits among the families,and growth-associated SSRswere used to analyze the effect of genotype on growth by genotyping.The results bymultiple comparisons showed that:themean value of body length,total length and body height of family AB2013F-7,AB2013F-89 and AB2013F-45 were significantly greater than other families(P<0.05);with family AB2013F-7 growing fastest in all three growth traits.During the 117 days of cultivation,the three families with the highest daily weight gain were AB2013F-7,AB2013F-89 and AB2013F-45(0.068 g/d,0.051 g/d and 0.048 g/d respectively).The results of genotyping of 11 pairs of parents showed that the growth rate of each family were in accord with the number of growth associated dominant genotypes.The number of dominant genotypes of each families and the mean of growth trait was positive correlation. Themore dominantgenotype the family had,the faster the growth rate of the family.
Paramisgurnus dabryanus;family;growth;genotype;SSR
Q173
:A
:1004-2091(2017)01-0001-06
10.3969/j.issn.1004-2091.2017.01.001
2016-04-29)
江蘇省科技廳農(nóng)業(yè)重大科技支撐項(xiàng)目(BE2012354);江蘇省水產(chǎn)三項(xiàng)工程項(xiàng)目(PJ2011-62);江蘇省農(nóng)業(yè)自主創(chuàng)新項(xiàng)目[CX(13)2042]
潘文賢(1958-),男,工程師,研究方向水產(chǎn)養(yǎng)殖.E-mail:PWX8210@163.com
凌去非,教授.E-mail:lingqf@suda.edu.cn
猜你喜歡
我愛(ài)學(xué)·笑話與口才(2025年3期)2025-02-24 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
趣味(語(yǔ)文)(2020年3期)2020-07-27 01:42:46
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
中國(guó)衛(wèi)生(2015年2期)2015-11-12 13:13:54
