丹江口水庫枯水期浮游細菌群落組成及影響因素研究
2017-02-22 07:24:02陳兆進丁傳雨朱靜亞杜宗明李玉英
中國環境科學 2017年1期
陳兆進,丁傳雨,朱靜亞,李 冰,黃 進,杜宗明,王 亞,李玉英*
(1.南陽師范學院農業工程學院,河南省南水北調中線水源區生態安全重點實驗室,河南 南陽 473061;2.南水北調中線水源區水安全河南省協同創新中心,河南 南陽 473061;3.南水北調中線渠首環境監測應急中心,河南 淅川 474475)
丹江口水庫枯水期浮游細菌群落組成及影響因素研究
陳兆進1,2,丁傳雨1,2,朱靜亞1,2,李 冰2,3,黃 進2,3,杜宗明2,3,王 亞2,3,李玉英1,2*
(1.南陽師范學院農業工程學院,河南省南水北調中線水源區生態安全重點實驗室,河南 南陽 473061;2.南水北調中線水源區水安全河南省協同創新中心,河南 南陽 473061;3.南水北調中線渠首環境監測應急中心,河南 淅川 474475)
為了解丹江口庫區南水北調中線工程通水后浮游細菌群落組成及影響因素,于2016枯水期(1月)對丹江庫區內5個典型生態點位表層水樣進行采集.水質監測結果表明庫區水質總體較好,除總氮和高錳酸鹽指數外,其他各項指標符合I類水標準要求.總氮(TN)和高錳酸鹽指數(CODMn)為庫區的主要污染因子,水體整體處于中營養狀態.采用高通量測序技術對浮游細菌群落組成進行研究,結果表明主要由放線菌門(Actinobacteria)、變形菌門(Proteobacteria)、藍藻門(Cyanobacteria)、擬桿菌門(Bacteroidetes)、疣微菌門(Verrucomicrobia)、壁厚菌門(Firmicutes)、酸桿菌門(Acidobacteria)等22個門和323個屬組成.5位點樣品具有豐富的群落組成,浮游細菌群落多樣性排序為:黑雞嘴>臺子山>庫心>宋崗>渠首.對浮游細菌群落組成與環境因子的關系進行典范對應分析(CCA),結果表明: T、pH值、NH4+-N、SD顯著影響浮游細菌群落結構.
丹江口庫區;浮游細菌;群落結構;高通量測序;環境因子
湖泊微生物中,浮游細菌數量巨大,在有機物降解及營養物循環過程中具有重要作用,是水生態系統的重要組成部分.全面了解湖泊生態系統中浮游細菌群落多樣性、分布特征及其在生態系統中的功能和作用,對于管理和維護湖泊生態環境具有深遠的意義[1-2].
由于只有不到 1%的微生物可以純培養,限制了自然界中絕大部分微生物的認識.基于免培養的變性梯度凝膠電泳、分子克隆文庫等技術被應用到對海洋、湖泊水體浮游細菌的研究之中,增加了人們對這一微生物群體多樣性及動態規律的認識[3].近年來,高通量測序技術的出現,大大加速了宏基因組學的發展.該技術操作簡單、成本低廉,能快速高通量得到特定的DNA 片段,較之前的分子生物多樣性研究能更全面地展示生物群落結構,極大地推動了生物多樣性研究[4,5].采用高通量測序和變性梯度凝膠電泳相結合的方法研究廈門外海微型浮游生物群落,結果表明MiSeq測序能得到更豐富的浮游生物群落信息,表現出技術上的優勢[6].
丹江口水庫作為南水北調中線工程核心水源區,其水質狀況直接關系到受水區居民的飲水安全.根據流域內降水特征及水文節律,每年 12月至次年4月為枯水期,此階段徑流量較小,不利庫區水體污染物稀釋、擴散、降解,水生態系統承受著巨大的污染壓力[7].有關丹江口庫區浮游細菌群落研究開展較少,本研究選取通水后第 2年枯水期(2016年1月)丹江庫區5個典型站位,采用高通量測序技術對浮游細菌群落16S rDNA多樣性進行分析,研究其群落組成和分布特征.結合典范對應分析(CCA)進一步探討細菌豐度、群落結構與環境因子間的關系,以期為丹江口水庫水環境保護提供參考依據.
1 材料與方法
1.1 樣品采集與水質理化指標測定
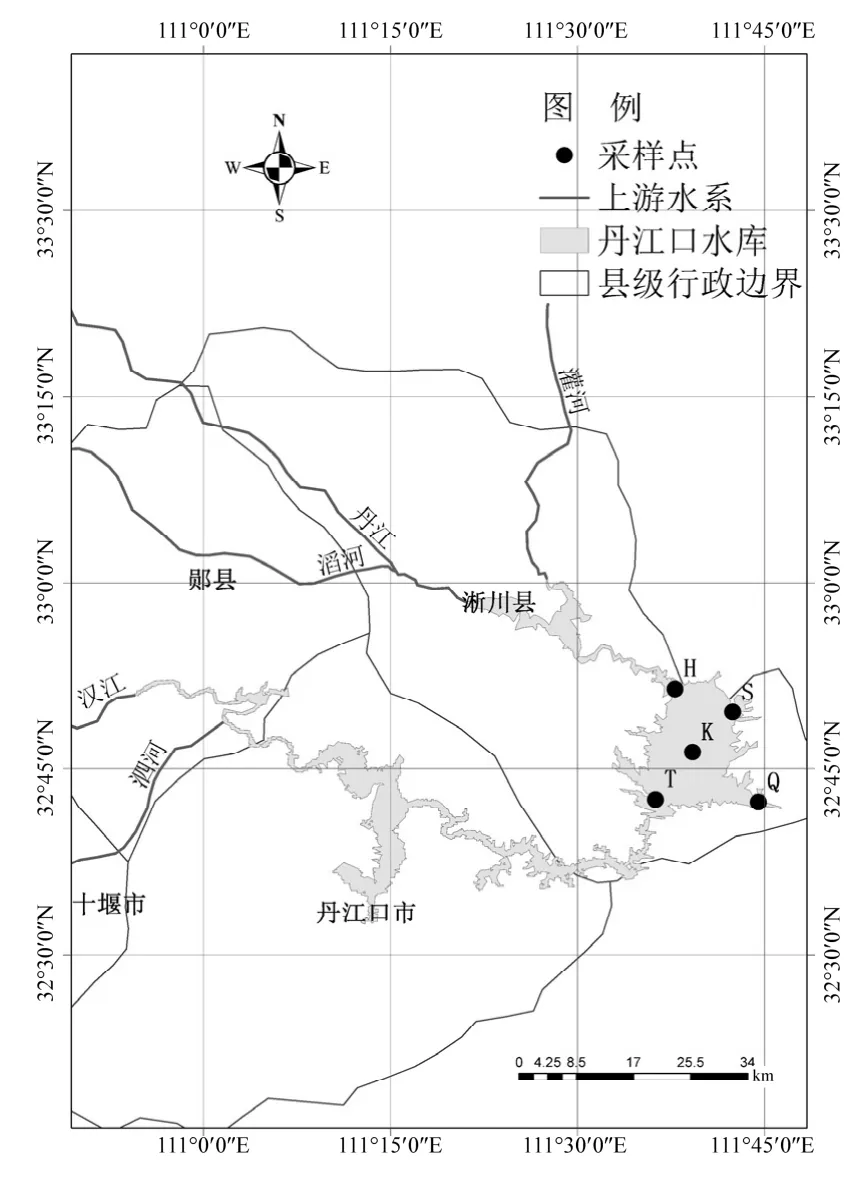
圖1 采樣點分布Fig.1 Location map of the sample sites of Danjiangkou reservoir
根據丹江口水庫的地理位置特征和人類活動影響程度,選取屬于河南省南陽市的丹江庫區庫心(樣品編號為K,無人為干擾)、渠首(樣品編號為 Q,重度人為干擾,陶岔渠首大壩取水口上游 100m)、黑雞嘴(樣品編號為 H,中度人為干擾,有居民點,上游支流匯入)、宋崗(樣品編號為S,重度人為干擾,有居民點、輪渡碼頭)和臺子山(樣品編號為 T,輕度人為干擾,為丹江庫區與漢江庫區分界線)5個位點為采樣點,具體位置如圖 1所示.根據庫區降水特征及水文節律,于2016年1月(枯水期)月初進行水樣的采集工作.參照《水和廢水監測分析方法》(第四版)[8]用柱狀采水器采集表層(深度 50cm)水樣 2L,所有點位均取平行樣3組. 500mL水樣用于提取細菌總 DNA,置于預先滅菌容器中存于冰盒,4h內運回實驗室進行檢測分析,剩余水樣用于水質指標測定.參照《水和廢水監測分析方法》(第四版)[8]測定水樣pH、水溫(T)、總氮(TN)、銨氮(NH4+-N)、總磷(TP)、透明度(SD)、電導率(Cond)、化學需氧量(COD)、生化需氧量(BOD)、溶解氧(DO)、高錳酸鹽指數(CODMn)以及葉綠素a含量(Chl a)理化指標.
1.2 樣品總 DNA提取
500mL表層新鮮水樣經0.22μm無菌微孔濾膜過濾,用以收集浮游細菌,將濾膜剪碎置于50mL無菌離心管中.按照 Omega Water DNA Kit (Omega, USA)的說明,提取水體中的DNA.將提取得到的細菌總DNA通過微量紫外分光光度計(NanoDrop?ND-1000, Wilmington, DE, USA)測定DNA濃度和純度.
1.3 高通量測序
采用通用引物(338F/806R)對浮游細菌 16S rRNA基因的V3~V4區擴增,修飾后的通用引物含有不同的 Tag標簽用以區分不同樣品[5].PCR擴增體系為20μL,其中含5×FastPfu Buffer 4μL、2.5mmol/L dNTPs 2μL、Forward Primer (5μmol/L 0.8μL、Reverse Primer(5μmol/L) 0.8μL、FastPfu Polymerase 0.4μL、DNA模板 10ng.補ddH2O至20μL.PCR擴增的反應條件為: 94℃,5min;30× (94℃,30s;54℃,30s;72℃.45s);72℃.10min.每個樣品3個重復,將不同樣品的PCR擴增產物均一化至10nmol/L后等體積混合,利用上海美吉生物醫藥科技有限公司的MiSeq PE300測序儀(Illumina Inc..San Diego.CA.USA)完成序列測定.
1.4 數據分析
1.4.1 高通量數據分析 高通量數據的生物信息學分析采用Qiime進行,根據序列的相似度,將序列歸為多個OTU,OTU產出后,統計各個樣品含有OTU情況及每個OTU中含有序列的數目,得到每個 OTU的分類學信息[9].選取相似度在97%條件下的OTU生成預期的稀釋曲線,并利用軟件mothur計算豐富度指數Chao1和ACE,覆蓋度指數(good's coverage)以及多樣性指數Simpson和Shannon指數進行Alpha 多樣性分析[10].利用 PCoA、Correlation聚類法分別進行數據處理、細菌群落分布、主成分分析和聚類分析.采用 Linear Discriminant Analysis(LDA) coupled with effect size measurements(LEfSe)在線工具尋找組與組之間有統計學差異的生物標志物[11].
1.4.2 典范對應分析 典范對應分析(CCA)能將可能相關的多個環境因子一起進行分析,可以很好地反映種群或群落與環境的關系.本文采用CANOCO軟件(版本 4.5) 進行浮游細菌和環境因子的CCA分析,排序結果用物種-環境因子關系的雙序圖表示[12].在主軸1和主軸2構成的平面中,箭頭表示環境因子,向量長短代表著相應環境因子在主軸中的作用,箭頭所處象限表示環境因子與排序軸間相關性的正負[13].
1.4.3 方差分析 浮游細菌群落多樣性與環境因子之間的相關性采用 SPSS19.0軟件進行分析.不同采樣點位間的環境因子比較采用單因素方差分析,顯著性水平設定為P<0.05,相關圖表制作在 Excel中完成.
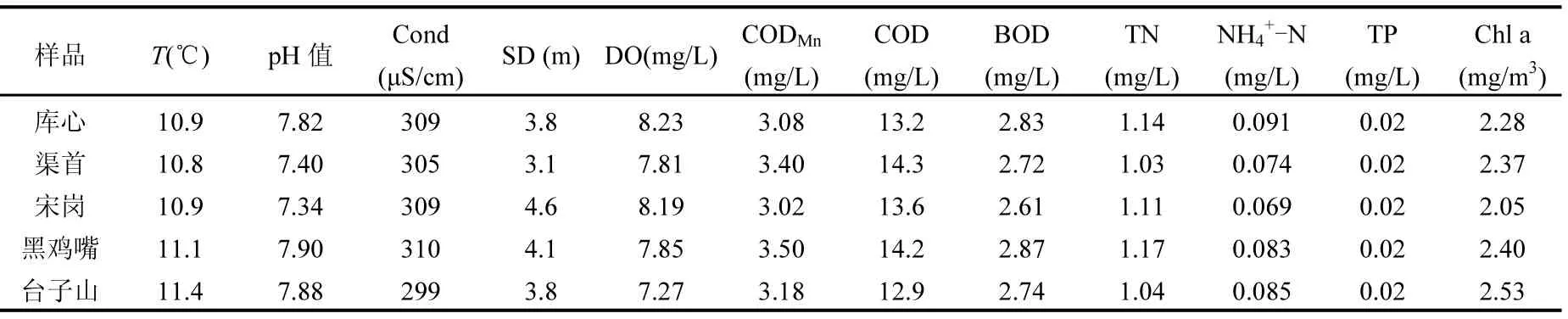
表1 各采樣點水體主要理化參數Table 1 The main physical and chemical properties of water samples
2 結果與分析
2.1 水質營養狀態評價
2016年1月對丹江口水庫庫心、渠首等5位點水質的理化監測結果表明各水體的水質總體較好,除總氮(均超過 1.00mg/L,為 IV類地表水標準)和高錳酸鹽指數外(均超過3.00mg/L,為II類地表水標準),其他各項指標符合《地表水環境質量標準》(GB38382-2002)I類水標準要求(表 1)[14].其中黑雞嘴總氮含量最高,其次為庫心和宋崗.從圖1可知黑雞嘴、庫心和宋崗集中分布于丹江庫區靠近丹江入水口,研究表明丹江流域存在農業面源污染,其中總氮明顯超標,是超標的原因[7,15-16].按照《地表水資源質量評價技術規程》(SL395-2007)[17]進行水庫營養狀態的評價,以總氮、總磷、高錳酸鹽指數、葉綠素a和透明度5個參數作為水質營養狀態參數,丹江庫區庫心、渠首等5位點總氮賦分值均為60,總磷、高錳酸鹽指數、葉綠素a和透明度的賦分值介于30~40之間.庫心、渠首等5位點營養狀態指數EI均為38.綜合評價,丹江庫區水體處于中營養狀態.
2.2 浮游細菌稀釋性曲線
丹江庫區浮游細菌稀釋性曲線隨測序條數的增加物種豐富度呈現前期增加后期趨向平緩(圖2).由圖2可知,庫心、渠首等5位點每個樣本的測序曲線在測序條數達到 20000條以上時物種數都基本趨向平穩,說明此時測序數據量合理,足夠代表物種的豐富度.
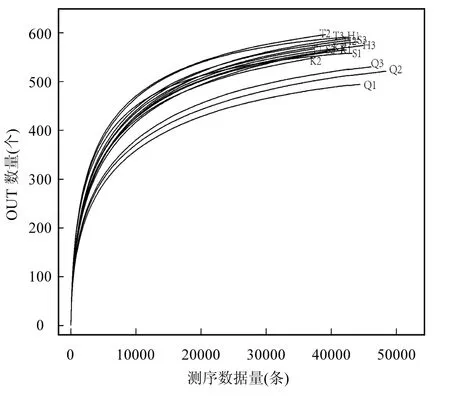
圖2 浮游細菌稀釋性曲線Fig.2 Rarefaction curve of bacterioplankton community
2.3 浮游細菌群落多樣性的評估
高通量測序表明丹江口水庫庫心、渠首等5位點樣品具有豐富的群落組成,其群落多樣性均較高(表2).5組樣品文庫覆蓋率均在99.80%以上,完全能夠反映該區域浮游細菌群落的種類和結構.渠首樣品OTU數、豐富度指數Chao1和ACE、Shannon指數低于其他組樣品,其中 OTU數、Chao1和ACE差異達到顯著水平(P<0.05),表明其浮游細菌群落多樣性低于其他樣品(表2).綜合OTU數、豐富度指數Chao1和ACE以及多樣性指數Simpson和Shannon指數,5組樣品浮游細菌群落多樣性排序為:黑雞嘴>臺子山>庫心>宋崗>渠首.

表2 浮游細菌群落多樣性評估表Table 2 Estimation of bacterioplankton community diversity
2.4 浮游生細菌多樣性的主成分分析
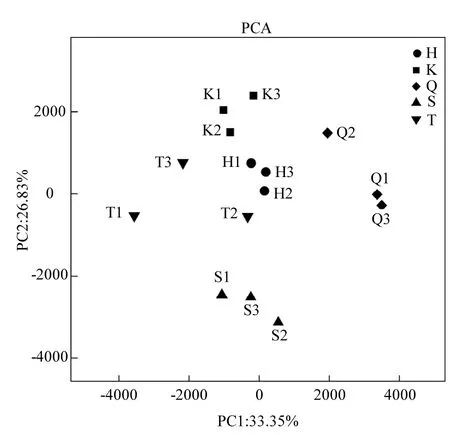
圖3 浮游細菌多樣性的主成分分析Fig.3 PCA analysis of bacterioplankton community diversity

圖4 不同樣品相似性聚類樹(A)和門水平上物種相對豐度分布(B)Fig.4 Similarity tree (A) and relative read abundance (B) of different bacterioplankton community structures at phylum levels in different treatments
利用Qiime 軟件對庫心、渠首等5位點樣品相關群落進行了主成分分析.如圖3所示,橫坐標 PC1貢獻度為33.35%,縱坐標 PC2貢獻度為26.83%,5位點中無人為干擾的庫心樣品聚集在PCA 分析圖上側.人為干擾較輕的臺子山樣品、黑雞嘴樣品與庫心樣品在PCA 分析圖中距離較近.人為干擾較重的宋崗樣品聚集于圖3下側,渠首樣品聚集于圖3右側,渠首和宋崗樣品與其他3組樣品浮游細菌群落差異較大.綜合以上分析表明不同樣品來源浮游細菌群落結構有所差異,其中人為干擾對浮游細菌群落影響較大.后續使用基于Unweighted UniFrac的UPGMA方法對庫心、渠首等5位點樣品浮游細菌群落構成的相似性進行聚類分析,通過樣品加權的聚類分析,同樣可以有效的將不同人為干擾程度樣品實現組間聚類(圖4A).
2.5 浮游細菌群落結構分析
高通量測序結果表明,15組實驗樣品平均序列條數為41916,OTUs平均數561(表2),經過分析主要為細菌的 22個門,包括放線菌門(Actinobacteria)、變形菌門 (Proteobacteria)、藍藻門 (Cyanobacteria)、擬桿菌門 (Bacteroidetes)、疣 微 菌 門 (Verrucomicrobia)、 壁 厚 菌 門(Firmicutes)、酸桿菌門 (Acidobacteria)等(圖4B).其中黑雞嘴、臺子山樣品包含19門,宋崗樣品包括18門,庫心樣品包括17門,渠首樣品包括16門.浮游細菌主要分布于323個屬,其中宋崗樣品包括245屬,黑雞嘴樣品包括239屬,臺子山樣品包含237屬,庫心樣品包括218屬,渠首樣品包括194屬.屬于放線菌門、變形菌門、藍藻門、擬桿菌門的序列總和占全部序列的 71.56%~ 83.53%,這些細菌為優勢種群.其中,放線菌門為庫心、黑雞嘴樣品最優勢種群和臺子山樣品第二優勢菌群,分別占總群落的27.86%、28.31%和23.68%;變形菌門為宋崗、臺子山樣品最優勢種群和庫心、黑雞嘴樣品第二優勢菌群,分別占總群落的29.72%、24.62%、20.49%和20.44%;藍藻門為渠首最優勢種群和宋崗第二優勢菌群,分別占總群落的 27.48%和 20.58%;擬桿菌門為渠首第二優勢菌群,占總群落的22.37%.
2.6 不同樣品差異細菌分析
為了進一步確定各組細菌種屬的豐富度差異,我們采用在線統計工具LEfSe來尋找宏基因組生物標志物.該方法是建立在統計學差異、生物學一致性和相同檢驗效能估計基礎之上的,通過分析高通量測序數據在每個樣品中的分類信息,在生物學上提供不同級別的可能的生物標志物[11].LEfSe法計算出不同樣品中的差異菌屬,所有菌在門、綱、目、科、屬水平的差異信息用餅形圖表示,餅形圖紅色、綠色、藍色、紫紅色、青色分別代表在黑雞嘴、庫心、渠首、宋崗和臺子山組中達到 LEfSe程序生物標志物篩選標準(圖5).
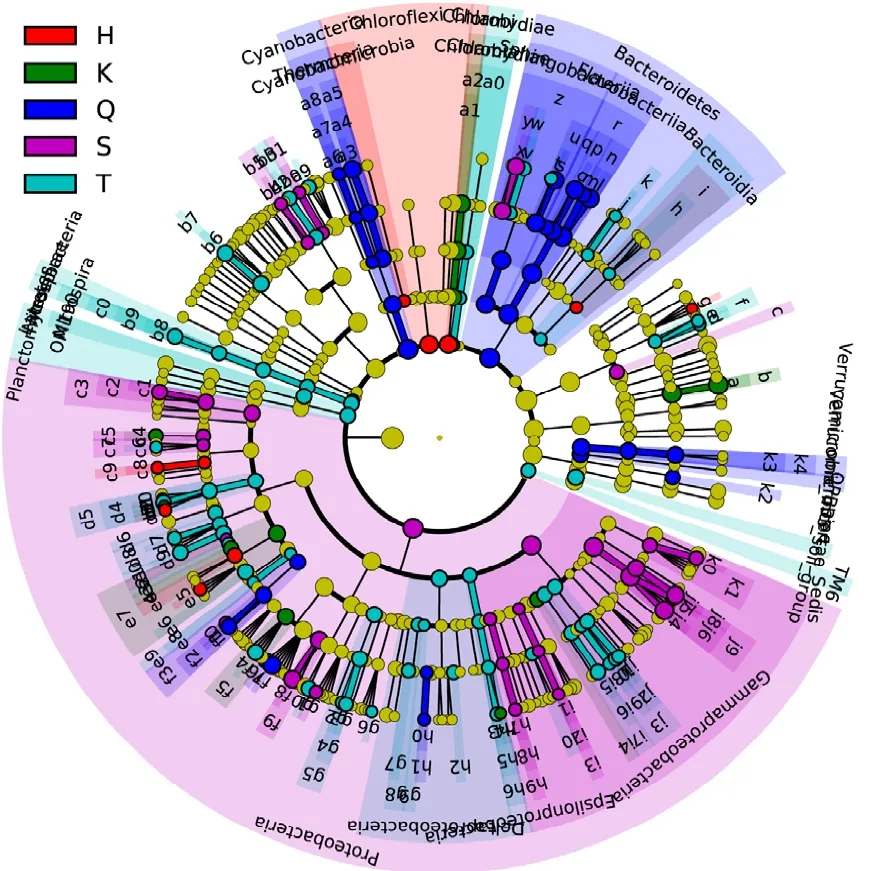
圖5 不同樣品差異細菌分布Fig.5 Taxon with statistical differences between different bacterioplankton communities
最內圈表示門水平的差異,依次往外的圈表示綱、目、科、屬,不同顏色扇形面積表示在此范圍內的菌均有差異(圖 5).在門的水平上,宋崗樣品中1個主要門(變形菌門)的細菌存在顯著差異,渠首樣品中2個門(藍藻門、擬桿菌門)的細菌存在顯著差異,臺子山和黑雞嘴樣品中各有2個門的細菌存在顯著差異,庫心樣品在門水平上無顯著差異.在屬的水平上,宋崗樣品中變形菌門的Brevundimonas、Sphingomonas、 Massilia、Deefgea、 Pseudomonas,壁 厚 菌 門 的Exiguobacterium、Planococcus、擬桿菌門的Pedobacter等共13屬的細菌存在顯著差異,渠首樣品中擬桿菌門的 Fluviicola、Owenweeksia、Flavobacterium,變形菌門的 Sphingopyxis、Albidiferax、Limnohabitans、Anaeromyxobacter、 Anaeromyxobacter,藍藻門的Synechococcus等共10個屬的細菌存在顯著差異,臺子山樣品中放線菌門的 Candidatus Aquiluna,擬桿菌門的Pseudarcicella、 Terrimonas,壁 厚 菌 門 的Oceanobacillus、Paenibacillus,變形菌門的Hyphomicrobium、Paracocccus、Defluviicoccus Reyranella、Limnobacter、Methylotenera等共20個屬細菌存在顯著差異,庫心樣品中放線菌門的CL500-29marine group 和 變 形 菌 門 的Bradyrhizobium、Roseovarius共3個屬細菌存在顯著差異,黑雞嘴樣品中放線菌門的Arthrobacter和變形菌門的 Methylobacterium、Gemmobacter、Candidatus Defluviella共4個屬細菌存在顯著差異.
2.7 浮游細菌群落組成與環境因子的相關分析
采用CCA分析水體環境因子與LEfSe分析篩選出的主要細菌之間的關系.由圖6可知,所分析的環境因子中T、TN、SD、Cond與第一排序軸 (AX1)正相關,其中SD相關系數最大,為0.881,呈極顯著正相關(P<0.01). pH、CODMn、BOD等與AX1負相關,均未達顯著水平. T、pH、NH4+-N與第二排序軸(AX2)正相關,相關系數都較大,分別為0.780、0.945、0.850,呈顯著正相關(P<0.05), Cond、DO、COD和CODMn與第二排序軸 (AX2)負相關.從第一、第二排序軸的相關性分析可以看出, T、pH、NH4+-N、SD對浮游細菌群落有較大影響, TN、DO、Cond等對其也有一定影響.相關性分析結果表明 Candidatus Aquiluna、Crenothrix、Methylobacter、Sulfuricurvum等 8個屬的細菌與 T呈顯著正相關, Anaeromyxobacter、Fluviicola與T呈顯著負相關. Defluviicoccus、Legionella、Nitrosomonas等 8個屬的細菌與 COD 呈顯著負相關, Synechococcus與 COD 呈 顯 著 正 相 關. Methyloversatilis 與 pH 呈 顯 著 正 相 關 , Methylomonas、Sulfuricurvum、Paracocccus與Cond呈顯著負相關, Sphingomonas與SD呈顯著正相關, Methylotenera、Pseudarcicella與DO呈顯著負相關, Defluviicoccus與CODMn呈顯著負相關, Aeromonas、Rheinheimera與NH4+-N呈顯著負相關,CL500-29marine group與NH4+-N呈顯著正相關, Candidatus Defluviella與TN呈顯著正相關. NH4+-N、TN、T、pH與浮游細菌多樣性指數中OTU數、豐富度指數Ace、Chao1、Shannon指數均呈正相關.

圖6 浮游細菌物種與環境因子的CCA排序Fig.6 CCA ordination biplot between bacterioplankton species and environmental factors
3 討論
3.1 南水北調通水后丹江口庫區枯水期水質測定
為了解蓄水后丹江口庫區水質狀況及影響庫區水質的主要污染因子,本研究測定了通水后第二年枯水期(2016年1月)丹江庫區5個典型站位水質,結果表明總氮和高錳酸鹽指數為庫區主要污染因子,對水庫營養狀態進行評價,EI指數均為 38,處于中營養狀態,與朱媛媛等
[7]結果基本一致.5位點中總氮含量較高的黑雞嘴、庫心和宋崗集中分布于丹江庫區靠近丹江入水口,徐國策等[15]研究表明丹江流域存在農業面源污染,其中總氮明顯超標,其可能是水體中總氮超標的主要原因.蓄水新增淹沒的農田面積近 26萬畝,研究表明農業面源污染已經成為丹江口水庫的主要污染源,水庫新增淹沒區農田養分物質釋放的潛在風險不容忽視[18].蓄水同時導致庫容大幅增加,流速降低,各項水文要素均發生了很大變化,而這些水文要素的變化相應地會帶來水環境要素的改變,從而影響庫區水質[19].
3.2 高通量測序揭示丹江口庫區浮游細菌群落組成
丹江口水庫作為南水北調中線工程的水源區,關于浮游細菌群落組成及其影響因素研究開展很少,相關工作亟待開展.本課題組曾于 2005年對水源區浮游微生物群落進行測定,選用選擇性培養基對細菌、真菌、放線菌、霉菌、異養菌、大腸桿菌、糞大腸桿菌的豐度和分布進行了調查研究,并分析了理化性質與水域微生物豐度和水溫、Chla、COD、BOD、DO等諸多生態因子的關系[20].在此之后,關于丹江口庫區浮游細菌群落研究鮮見報道.為了解蓄水后丹江口庫區浮游細菌群落組成,本實驗采用Illumina公司MiSeq測序平臺對丹江庫區庫心、渠首等5位點浮游細菌群落組成進行了研究,發現其主要由放線菌門、變形菌門、藍藻門、擬桿菌門、疣微菌門、壁厚菌門、酸桿菌門等22個門, CL500-29marine group、Limnohabitan、Bacillus、Synechococcus等323個屬組成,表現出群落組成的豐富性.丹江口庫區浮游細菌群落組成與之前研究的鄱陽湖、太湖、滇池等淡水湖泊[2,21-24]以及長江、南明河等河流[25-26]及其河口[3]等水體典型的細菌組成類群相似,這些典型的淡水細菌大都隸屬于變形菌門、藍細菌門、擬桿菌門、放線菌門和疣微菌門等[2,26].本次實驗結果與 2005年研究[20]相比,更全面、更準確了解丹江口庫區浮游細菌群落組成,揭示了丹江口庫區的細菌種群類別,為丹江口庫區水質保護提供重要的依據.
3.3 浮游細菌群落與環境因子的關系
研究表明,水體中的環境因子,包括湖泊的形態特征(例如:湖泊的大小和深度)、物理化學特征(例如:溫度、pH、鹽度、無機營養鹽等)、有機質的濃度和類型以及食物網的組成和物種間的相互作用等均能影響浮游細菌的群落組成
[2].本實驗采用Illumina公司MiSeq測序平臺對丹江庫區庫心、渠首等5位點浮游細菌群落組成進行了研究,采用在線統計工具LEfSe分析各組細菌種屬的豐富度差異,同時對差異浮游細菌群落與理化指標進行CCA分析.結果表明, T、pH、NH4+-N、SD對浮游細菌群落有較大影響, Candidatus Aquiluna、Crenothrix等8個屬的細菌與T呈顯著正相關, Anaeromyxobacter、Fluviicola與 T呈顯著負相關.擬桿菌門的CL500-29marine group屬是浮游細菌最為優勢種群,占 5組樣品浮游細菌的 10.18%~16.74%,相關性分析表明與 NH4+-N呈顯著正相關.另外,Aeromonas、Rheinheimera與NH4+-N呈顯著負相關, Candidatus Defluviella與TN呈顯著正相關.在各種水生態環境中,溶解無機氮是異養細菌重要的 N源,同時過量的氮可能導致水體富營養化,造成水質惡化.Haukka等[27]報道在淡水生態系統中添加總氮和磷酸鹽后,浮游細菌的群落結構發生變化,這種變化可能與生態系統中添加營養物質后引起的水體富營養化有關.丹江流域存在農業面源污染,其中總氮明顯超標[15,18],可能會影響丹江口水庫浮游細菌群落組成.
4 結論
4.1 2016年1月丹江口水庫庫心、渠首等5位點監測結果表明水體的水質總體較好,除總氮和高錳酸鹽指數外,其他各項指標符合 I類水標準要求.以總氮、總磷、高錳酸鹽指數、葉綠素 a和透明度5個參數作為水質營養狀態參數,丹江口水庫水體處于中營養狀態.
4.2 高通量測序表明丹江口水庫具有豐富的浮游細菌群落組成,主要由放線菌門、變形菌門、藍藻門、擬桿菌門、疣微菌門、壁厚菌門、酸桿菌門等22個門和323個屬的細菌組成.浮游細菌群落多樣性排序為:黑雞嘴>臺子山>庫心>宋崗>渠首.
4.3 浮游細菌與環境因子 CCA分析表明,T、pH、NH4+-N、SD等對丹江口浮游細菌群落有較大影響,不同環境因子對于不同菌群具有不同影響.
[1] Brendan Logue J, Lindstr?m ES. Biogeography of Bacterioplankton in Inland Waters [J]. Freshwater Reviews, 2008,1(1):99-114.
[2] 任麗娟,何 聃,邢 鵬,等.湖泊水體細菌多樣性及其生態功能研究進展 [J]. 生物多樣性, 2013,21(4):421-432.
[3] 闞金軍,孫 軍.河口細菌群落多樣性及其控制因素:以切薩皮克灣為例 [J]. 生物多樣性, 2011,19(6):770-778.
[4] van Dijk E L, Auger H, Jaszczyszyn Y, et al. Ten years of next-generation sequencing technology [J]. Trends in Genetics, 2014,30(9):418-426.
[5] 張軍毅,朱冰川,超 徐,等.基于分子標記的宏基因組16S rRNA基因高變區選擇策略 [J]. 應用生態學報, 2015,26(11):3545-3553.
[6] Yu L Y, Zhang W J, Liu L M, et al. Determining microeukaryotic plankton community around xiamen island, southeast china, using Illumina MiSeq and PCR-DGGE techniques [J]. PLoS ONE, 2015,10(5):e0127721.
[7] 朱媛媛,田進軍,李紅亮,等.丹江口水庫水質評價及水污染特征[J]. 農業環境科學學報, 2016,35(1):139-147.
[8] 國家環境保護總局.水和廢水監測分析方法(第四版) [M]. 北京:中國環境科學出版社, 2002:239-284.
[9] Caporaso J G, Kuczynski J, Stombaugh J, et al. QIIME allows analysis of high-throughput community sequencing data [J]. Nature Methods, 2010,7(5):335-336.
[10] Schloss P D, Westcott S L, Ryabin T, et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported software for Describing and comparing microbial communities [J]. Applied and Environmental Microbiology, 2009,75(23):7537-7541.
[11] Segata N, Izard J, Waldron L, et al. Metagenomic biomarker discovery and explanation [J]. Genome Biology, 2011,12(6):1-18.
[12] Ter Braak CJ, Smilauer P. CANOCO reference manual and CanoDraw for Windows user's guide: software for canonical community ordination (version 4.5) [M]. Microcomputer Power, Ithaca, NY, 2002.
[13] 任學敏,楊改河,朱 雅,等.環境因子對太白山高山植被物種組成和豐富度的影響 [J]. 生態學報, 2014,34(23):6993-7003.
[14] GB/3838-2002 地表水環境質量標準 [S].
[15] 徐國策,李占斌,李 鵬,等.丹江中游典型小流域土壤總氮的空間分布 [J]. 地理學報, 2012,67(11):1547-1555.
[16] 肖春艷,俐 武,趙同謙,等.南水北調中線源頭區蓄水前土壤氮磷分布特征 [J]. 中國環境科學, 2013,33(10):1814-1820.
[17] SL/395-2007 地表水資源質量評價技術規程 [S].
[18] 王 劍,尹 煒,趙曉琳,等.丹江口水庫新增淹沒區農田土壤潛在風險評估 [J]. 中國環境科學, 2015,35(1):157-164.
[19] 馮 靜,何太蓉,韋 杰.三峽工程蓄水前后庫區水質變化及對策分析 [J]. 重慶師范大學學報(自然科學版), 2011,28(2):23-27.
[20] 李玉英,張乃群,胡蘭群,等.南水北調中線工程水源區微生物菌群研究 [J]. 水利漁業, 2006,26(4):57-59.
[21] Niu Y, Shen H, Chen J, et al. Phytoplankton community succession shaping bacterioplankton community composition in Lake Taihu, China [J]. Water Research, 2011,45(14):4169-4182.
[22] 呂明姬,汪 杰,范 錚,等.滇池浮游細菌群落組成的空間分布特征及其與環境因子的關系 [J]. 環境科學學報, 2011,31(2): 299-306.
[23] Dai Y, Yang Y, Wu Z, et al. Spatiotemporal variation of planktonic and sediment bacterial assemblages in two plateau freshwater lakes at different trophic status [J]. Applied Microbiology and Biotechnology, 2016,100(9):4161-4175.
[24] 寇文伯,黃正云,張 杰,等.鄱陽湖湖泊細菌群落組成及結構-以松門山為例 [J]. 生態學報, 2015,35(23):7608-7614.
[25] 毛 莉,張 明,白 芳,等.三峽庫區支流藍藻水華對浮游細菌群落組成的影響 [J]. 水生生物學報, 2016,40(3):609-614.
[26] 唐 婧,徐小蓉,商傳禹,等.南明河城區河段細菌多樣性與環境因子的關系 [J]. 微生物學報, 2015,55(8):1050-1059.
[27] Haukka K, Kolmonen E, Hyder R, et al. Effect of nutrient loading on bacterioplankton community composition in lake Mesocosms [J]. Microbial Ecology, 2006,51(2):137-146.
Community structure and influencing factors of bacterioplankton during low water periods in Danjiangkou Reservoir.
CHEN Zhao-jin1,2, DING Chuan-yu1,2, ZHU Jing-ya1,2, LI Bing2,3, HUANG Jin2,3, DU Zong-ming2,3, WANG Ya2,3, LI Yu-ying1,2*
(1.Key Laboratory of Ecological Security for Water Source Region of Mid-line Project of South-to-North Diversion Project of Henan Province, College of Agricultural Engineering, Nanyang Normal University, Nanyang 473061, China;2.Henan Collaborative Innovation Center of Water Security for Water Source Region of Mid-line Project of South to North Diversion Project, Nanyang 473061, China;3.Emergency Centre for Environmental Monitoring of Mid- line Project of South to North Water Division, Xichuan 474475, China). China Environmental Science, 2017,37(1):336~344
The Danjiangkou reservoir is the main source of water for the “middle-route” of the South to North Water Diversion Project in China. Its water quality directly affects the safety of drinking water for residents along this route. In this study, 15surface-water samples were collected from the reservoir at low-water period (January) in 2016, and 12 essential environmental factors were investigated. The results showed that the water quality in the reservoir was strongly influenced by total nitrogen (TN) and the permanganate index, with the water quality being ranked at IV or II for all locations. The water in this reservoir was in the mesotrophic state. The high-throughput sequencing approach was adopted to analyze the distribution characteristics of the community structure and diversity of bacterioplankton and their relationship to environmental factors. Phylogenetic analysis based on 16S rDNA sequences showed that bacterioplankton could be divided into 22 major phylogenetic groups. The dominant phylogenetic groups included Actinobacteria, Proteobacteria, Cyanobacteria, Bacteroidetes, Verrucomicrobia, Firmicutes, and Acidobacteria. At the genus level, the dominant phylogenetic groups included CL500-29 marine group, hgcI clade, Fluviicola, Sediminibacterium, Flavobacterium, LD28 freshwater group, Limnohabitans, Synechococcus, Bacillus, and Cyanobacteria norank. Diversity of the bacterioplankton community in the Danjiangkou reservoir can be ranked in the following order: Heijizui >Taizishan>Kuxin>Songgang>Qushou. The results of canonical correspondence analysis (CCA) for the relationship between the22 phyla of bacterioplankton and 12environmental factors showed that the T, pH, NH4+-N, and SD were the main environmental factors affecting the distribution characteristics of the bacterioplankton community.
Danjiangkou Reservoir;bacterioplankton;community structure;high-throughput sequencing;environmental factor
X524
A
1000-6923(2017)01-0336-09
陳兆進(1985-),男,安徽合肥人,講師,博士,主要從事土水污染控制、修復與資源化利用研究.發表論文10余篇.
2016-03-22
國家自然科學基金項目(41601332);河南省教育廳高等學校重點科研項目(16A210012,14A180015);南陽師范學院高層次人才科研啟動費資助項目(ZX2014069)
* 責任作者, 教授, lyying200508@163.com
