GLP-1R結構和功能及小分子藥物篩選研究進展
2017-02-22 06:51:08胡中平程念楊帆蘇正定
生物技術通報 2017年2期
關鍵詞:信號
胡中平 程念 楊帆 蘇正定
(湖北工業大學生物醫藥研究院 工業發酵協同創新中心 教育部發酵工程重點實驗室,武漢 430068)
GLP-1R結構和功能及小分子藥物篩選研究進展
胡中平 程念 楊帆 蘇正定
(湖北工業大學生物醫藥研究院 工業發酵協同創新中心 教育部發酵工程重點實驗室,武漢 430068)
胰高血糖素樣肽-1受體(glucagon-like peptide-1 receptor,GLP-1R)作為2-型糖尿病(T2DM)藥物研發和治療的靶點有著十分重要的臨床意義。盡管通過結構生物學,蛋白質工程等方法和手段對于GLP-1R結構的研究有了較大突破。但是關于其全長結構解析,多肽結合受體的分子機理及受體激活的內在機制還不曾得到解決。近些年有關GLP-1R相關研究發展較快,簡述了該受體的結構與功能以及已有的小分子藥物先導化合物,并討論GLP-1受體分子結構作用機制的發展方向及應用前景,旨為進一步探尋2型糖尿病的治療方案提供有利的幫助。
GLP-1R;分子結構;小分子藥物
胰高血糖素樣肽-1受體(glucagon-like peptide-1 receptor,GLP-1R)是2-型糖尿病最為有效的治療靶點之一。經過多年基礎研究積累,轉化和臨床研究表明GLP-1與其受體相互作用能夠有效調控機體糖穩態和能量代謝[1]。GLP-1R屬于G蛋白偶聯受體(GPCR)B簇亞族(B1)的一員,它的典型特征是具有一個相對比較大的胞外域(ECD)和有α-螺旋束構成的7次跨膜核心域(TMD)。GLP-1R作為GLP-1/GLP-1R途徑下游信號的靶標,主要是通過“two-domain model”來激活受體。首先GLP-1 C端域(cGLP-1)同GLP-1R胞外域(ECD)形成的“affinity trap”結合,從而確保GLP-1 N端域(nGLP-1)與受體核心域(TMD)形成的“pocket”互交。這種相互作用能夠有效激活PKA、PI3K、MAPK等多種下游信號通路,參與諸如胰島素的釋放,β-細胞增生,胰高血糖素釋放減少,延遲胃排空[2],增強記憶等重要生理過程[3]。所以,GLP-1R作為GLP-1發揮效能的靶點對于人們研究多種疾病有著潛在的指導意義。本文就GLP-1R的結構和功能的研究進展進行綜述,并針對基于該受體的小分子先導化合物進行歸納,以便為進一步開發合適治療T2DM的口服藥物提供借鑒。
1 GLP-1R的結構特性概況
胰高血糖素樣肽(GLP-1)和葡萄糖依賴性促胰島素多肽(GIP)作為腸促胰素都能夠直接作用于胰島細胞加強餐后胰島素釋放[4,5],但是在治療2-型糖尿病(T2DM)方面首選GLP-1。主要有兩個原因:首先,GLP-1和GIP都會抑制胃的排空,延遲餐后血糖升高,但只有GLP-1會引起飽足感;其次,這兩種多肽對胰高血糖素的釋放具有相反的效用,GLP-1起抑制作用但GIP會促進胰高血糖素釋放[6]。GLP-1主要是由胰高血糖素原基因表達通過腸L細胞分泌的多肽類激素,其很容易被DPP-Ⅳ降解成無生理活性GLP-1(9-37),半衰期較短僅有1-2 min[6]。GLP-1發揮生理功能主要是通過結合并激活G蛋白偶聯受體B家族(分泌素家族)中的GLP-1R,誘發信號分子cAMP增加,使偶聯的G蛋白α亞基與β,γ亞基解離,并分別介導胞內不同信號通路來完成的[7]。
1.1 GLP-1R的表達及糖基化
人的GLP-1受體(hGLP-1R)基因位于人染色體6p21上,編碼463個氨基酸。GLP-1R表達于模式生物和人體中并呈現出高度保守性,它在機體中廣泛分布于被檢測的胰島、胃、小腸、心臟、腎臟、肺及大腦等組織中。GLP-1R在胰島β細胞中表達較多,在人胰島α和δ細胞中表達尚屬爭議[8]。人GLP-1受體(hGLP-1R)同小鼠GLP-1受體(rGLP-1R)序列具有高度相似性,同源性達到84%。該受體屬于G蛋白偶聯受體B簇中胰高血糖素受體亞家族,這類受體有3個顯著的特征:一個相對較長的大約100-150個氨基酸的胞外N端域(ECD),與之相連的7次跨膜結構域(7TMD)以及連接跨膜段的相對較短的胞內域C端域(ICD)[9]。其中胞外域(ECD)包括兩組通過loop環相連的反向β折疊,6個保守的半胱氨酸形成的3對二硫鍵以及一個柔性的α螺旋(圖1,參照PDB ID:2QKH[10])。研究發現在RINm5F細胞[11]中,糖基化肽酶F能夠使GLP-1受體分子量從63 kD減少到51 kD,表明GLP-1受體的N端連接有聚糖分子。衣霉素也會減少GLP-1受體的表達,但不會影響其配體親和力。在重組CHO細胞中,任何兩三個N端糖基化位點(Asn63、Asn82、Asn115)的突變都會阻止受體穿梭于質膜[12]。在HEK-293細胞中,GLP-1R胞外域(ECD)存在的信號肽(23個氨基酸)會在GLP-1受體正確加工和運輸過程中被切斷,以便在質膜形成成熟和完整的糖基化。此外,抑制 hGLP-1R糖基化會阻礙該受體細胞穿膜表達[13,14]。可見,糖基化和信號肽對GLP-1R的穿梭和加工十分重要。
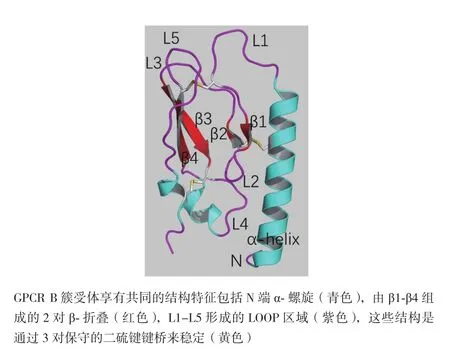
圖1 GPCR B簇受體胞外域(ECD)通用結構示意圖
1.2 GLP-1R胞外域(ECD)特征
鑒于人們研究GLP-1R全長的長期性和復雜性,單獨分離出GLP-1R胞外域(ECD)已經成為了探究GLP-1受體結構和功能的重要手段。1996年,Wilmen等[15]運用大腸桿菌表達ECD(殘基20-144)并通過六聚組氨酸標簽純化到可溶性片段。盡管純化的具有活性功能蛋白也能像GLP-1R全長受體一樣同125I-GLP-1(用碘125標記的GLP-1)發生互交作用,但是這些純化的蛋白處于較低的表達水平,之后用β-巰基乙醇處理發現125I-GLP-1結合受體的能力喪失。2002年,Bazarsuren等[16]也通過大腸桿菌表達出GLP-1R胞外域。與前者不同之處在于表達的蛋白是以包涵體形式存在,后來經過變性、重折疊和復性成功的做出了具有生物活性的GLP-1R胞外域(ECD),實現了蛋白可逆性復性。之后用胃蛋白酶處理發現存在有二硫鍵,進一步分析證實二硫鍵的位置分別是Cys46和Cys71、Cys62和Cys104以及Cys85和Cys126。這些具有保守性二硫鍵對蛋白結構的穩定具有至關重要的意義。2008年,Runge等[17]解析出GLP-1R胞外域同Exendin-4(9-39)形成的復合體,其分辨率為2.2 ?晶體結構。該結構通過N端的α螺旋和兩個反向β延伸形成的loop結構闡釋了其疏水結合位點并發現了Asp67、Trp72、Pro86、Arg102、Gly108和Trp110六個保守的氨基酸殘基。其中Asp67形成分子內相互作用,通過水分子與Arg102相互作用并直接作用于Trp72和Arg121(圖2,參照PDB ID:3C5T數據[17])。另外,Asp67還會與Tyr69和Ala70作用來穩定β1和β2鏈的反轉。可見,這些殘基對胞外域折疊和受體結構穩定必不可少。2009年,Chritina等[18]解析出GLP-1胞外域同GLP-1形成的復合體,其分辨率為2.2 ?晶體結構。該結構表明激動劑和拮抗劑與受體結合是通過疏水性相互作用來完成的并且這種作用呈現保守性,但是配體結合位點的某些殘基賦予了GLP-1的特殊構象(圖3-A,3-B,參照PDB ID:3IOL數據[18])。當GLP-1結合受體胞外域時,會呈現出扭曲構象,但從Thr13到Val33展現出連續的α-螺旋結構,這與之前解析的Exendin-4(9-39)與GLP-1R-ECD形成的晶體復合體結構有所不同(圖3-C)。2011年,Day等[19]發現GLP-1受體N端域Glu68殘基的改變會使GLP-1的親和力減少8倍,加強GLP-1多肽的C端的正電荷有利于多肽傾向于受體68位氨基酸殘基處靠攏,說明Glu68氨基酸對于配體的行為十分重要。最近研究表明,GLP-1受體N端域較大的疏水性作用能夠驅使多肽的結合并定位于配體N端激活受體[20]。高蔚豐等[21]通過S52R突變會引起GLP-1受體N端片段活性失活,但缺失前面20個氨基酸以及后面10個氨基酸都不會影響N端域的生物活性,表明Ser52對于維持配體活性至關重要。盡管在探究胞外域結構方面戰果累累,然而與所預期的GLP-1R胞外域結構相比,ECD上的很多殘基未曾出現過共價修飾。
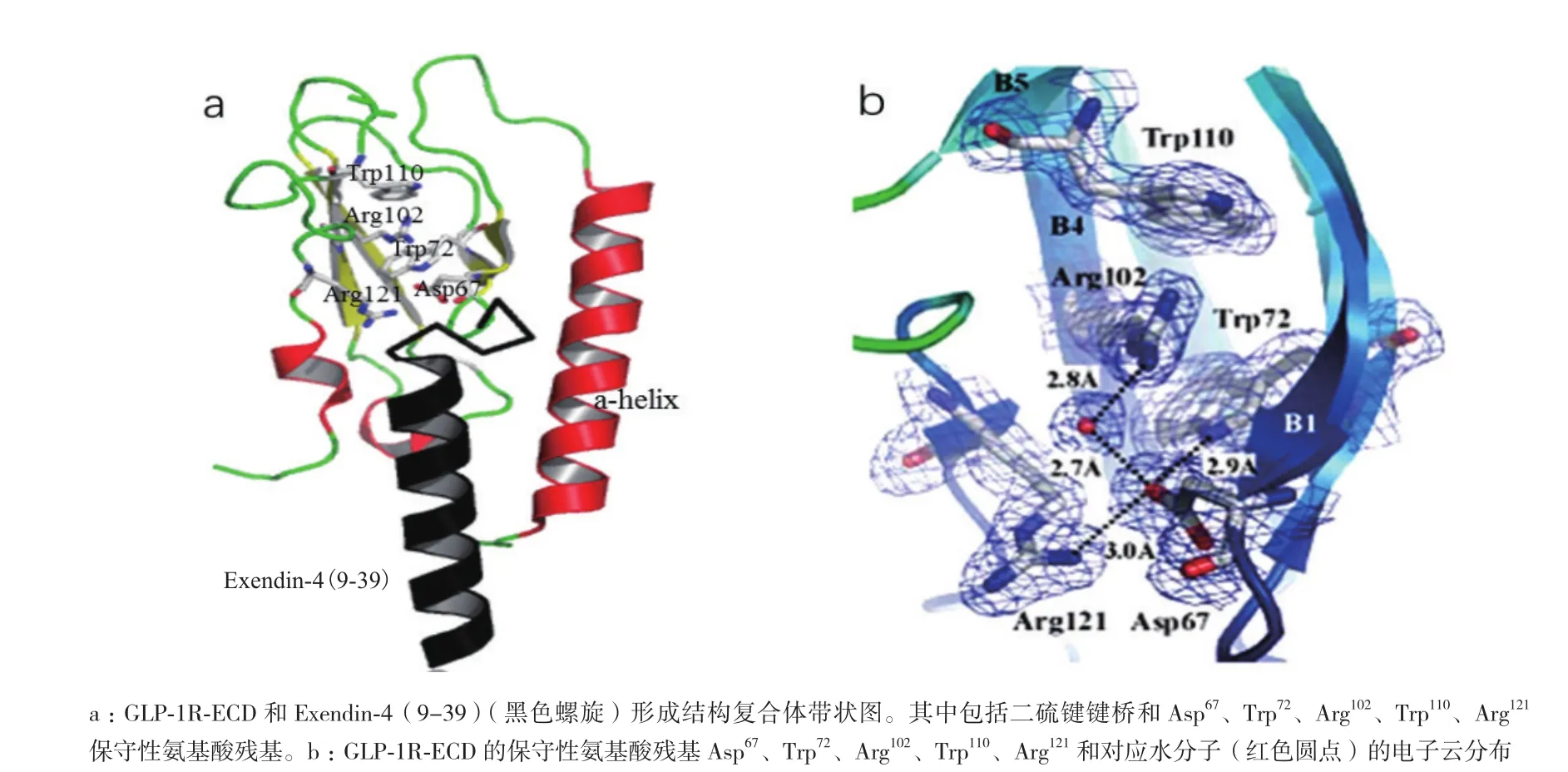
圖2 GLP-1R胞外域(ECD)與Exendin-4(9-39)形成復合物結構[17]
1.3 GLP-1R核心域特征
由于受體跨膜域結構的特殊性,人們主要是通過定點誘變以及蛋白互交實驗等相關手段來探究其結構。Xiao等[22]對跨膜域 TM2-TM3的5個殘基(K197A、D198A、K202A、D215A、R227A) 進 行定點誘變,發現配體結合力存在明顯的下降(IC50值相對于WT高出4-20倍)。研究發現,GLP-1受體第二個跨膜螺旋(TMH2)的膜外端的一個殘基Asp198[23]和第4個跨膜螺旋(TMH4)的殘基Lys288[24]特異性的參與了GLP-1R N端域的結合。將M204A和Y205A進行雙突變[25]發現該突變受體同GLP-1的親和力相對于野生型(WT)受體明顯減少了30倍,而Exendin-4只減少了3倍并且對拮抗劑卻沒有影響,原因可能是這兩個位點側鏈的疏水性發生了改變。研究調查表明,Ⅱ型糖尿病患者胰島β細胞中GLP-1受體出現 T149M天然突變會引起GLP-1 結合力下降并且能夠降低多肽誘使cAMP調控信號的效能,但cAMP的含量不變。說明這種受體的多態性會影響其結合激動劑的效能[26]。Mann等[27]發現Cys226(TM3)和Cys296(ECL2)之間會形成二硫鍵,并靠近受體激活“pocket”。進一步通過ECL2上殘基定點誘變得出該loop對于激動劑激活受體十分重要。Coopman等[28]發現跨膜域的一些殘基(K197A、W284A、R310A等)的突變更傾向與受體效能的降低。人們用點突變和嵌合GLP1R/ GIPR同嵌合GLP-1/GIP 相互作用的分子模擬[29]表明GLP-1的His1和Thr7會同GLP-1受體中的殘基Asn302(ECL2)、Ile196(TMH2)和Leu232/Met233(ECL1)相互作用,這為配體與受體結合以及受體激活提供了很重要的線索。Cassandra等[30]通過將跨膜段第2個loop域的相應殘基(C296A、D297A、R299A、N300A、N302A、N304A、Y305A和L307A等)進行突變都會影響到GLP-1的結合力和耦合效率及CAMP形成,鈣離子內流和胞外信號調控激酶1和2的磷酸化(PERK1/2)的激活,說明第2個loop域的結構對配體結合十分重要,尤其是Trp306Ala會使受體的生物活性喪失。Jin等[31]通過GLP-1R突變發現ECL3上保守殘基Arg380與疏水性殘基Leu379和Phe381相連可能會同GLP-1的Asp9和Gly4相互作用。后來又用VPAC1R的ECL3去取代GLP-1R的ECL3區域形成GLP-1Rs嵌合受體,結果表明此嵌合受體同GLP-1的結合比較弱,說明GLP-1R受體的ECL3對于受體激活顯得十分重要。進一步進行氨基酸點突變表明,GLP-1R受體保守殘基Arg380以及疏水性殘基Leu379和Phe381可以調控GLP-1的Gly4和Asp9的相互作用。從而提出了配體結合“pocket”主要由GLP-1受體TMH2,ECL1,ECL2和 ECL3的保守性殘基構成。近些年,發現了C端對GLP-1R跨膜域表達和內源化產生很大影響,研究表明氨基酸殘基411-418對于跨膜域定位于質膜很關鍵,氨基酸殘基419-430可能對于受體耦合Gαs,cAMP誘使產生受體活性十分重要,氨基酸殘基431-450對于激動劑誘導hGLP-1R內源化是必不可少的[32]。通過分子模擬發現,受體胞外域靠近跨膜核心區的ECL3并且GLP-1R同配體結合也存在著“closed state”和“open state”兩種形式[33],這與之前的胰高血糖素受體(GCGR)與配體結合狀況相似[34]。
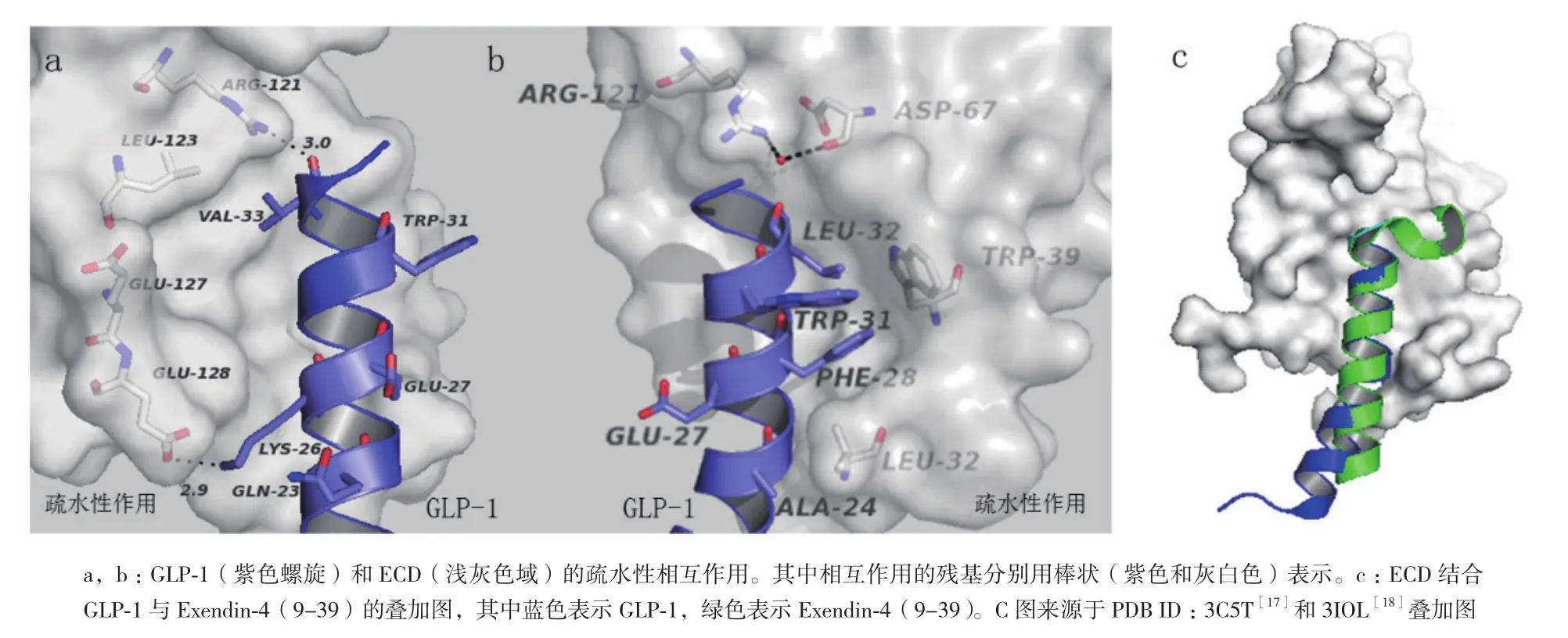
圖3 GLP-1R胞外域(ECD)同GLP-1疏水性相互作用
2 GLP-1R的功能相關性研究
2.1 受體激活
GLP-1R屬于G蛋白偶聯受體B簇亞類。GPCR B簇受體與配體作用機制主要是被大家公認的“twodomain model”[35](圖4)。配體的C端螺旋首先同GLP-1R胞外域(ECD)結合,從而確保配體的N端同GLP-1受體的核心區域(TMD)進行二次交合,后者相互作用對于激動劑激活受體至關重要[35]。因此提出了解釋GPCR B簇受體激活模型,即配體結合誘使內源性激動劑構象改變,然后同受體核心域相互作用產生活性[36]。已經報道了配體的α螺旋二級結構在二域模型中調控受體胞外域ECD的起始作用十分重要,并提出了螺旋包被模型來解釋GPCR B簇不同配體是怎樣激活受體[9]。
GLP-1和Exendin-4都是α-螺旋的多肽,能與GLP-1R胞外域多個接觸位點相互作用誘導受體信號。它們的兩性特征決定了它們同受體ECD相互作用呈現出保守性,其中配體的疏水面是相互作用的關鍵并且受體激動劑多肽疏水區形成的微小結合能對結合最為關鍵。受體激活模型的第二階段表明,ECD對接多肽直接促進配體N端同受體核心“pocket”相互作用引起跨膜α-螺旋構象重排,誘使膜內Loop環刺激胞內信號傳導。
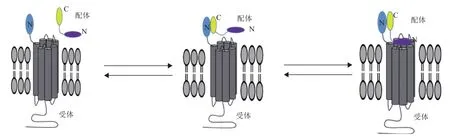
圖4 G蛋白偶聯受體與其配體結合的“two-domain model”
2.2 GLP-1R信號傳導調控機制
一般來說,GLP-1受體N端主要是識別特異性的配體,但是受體的核心區對于信號特異性傳導發揮重要作用。GLP-1R屬于G蛋白偶聯受體中Gs亞類。它是一種多效性偶聯受體,主要通過與多種G蛋白(Gαs、Gαi、Gαo和Gαq/11)偶聯來調控細胞通路。當與GLP-1結合后,G蛋白α亞基與β、γ亞基解離并對不同信號通路進行介導[7]。在β細胞中偶聯Gαs蛋白,激活腺苷環化酶,促使cAMP在細胞內含量升高并增加蛋白激酶A(PKA)和cAMP-2激活交換蛋白(Epac2)的含量,引起離子通道活性改變,鉀離子通道關閉,電壓依賴性鈣離子通道打開(VDCCs),鈣離子內流,胰島素原基因轉錄增加,胰島素分泌小泡釋放。GLP-1可以通過cAMPPKA途徑提高葡萄糖感受性,刺激血糖依賴性胰島素的分泌[37]。除此之外,GLP-1R受體還可以通過G蛋白β、γ亞基來激活磷脂酰基醇-3-激酶(PI3K)和絲裂原活化蛋白激酶(MAPK)的信號通絡誘導β細胞的增值和分化;另外,cAMP還可以以不依賴PKA的方式,通過與β細胞中cAMP調節的鳥苷酸交換因子(cAMP-regulated guaninenucleotide exchange factors,cAMP-GEFs)相互作用,激活Ras/MAPK[38](mitogen-activated protein kinase)信號通路,促進β細胞的生長和分化[38]。不僅如此,GLP-1R還能夠通過調控cAMP反應元件結合蛋白(CREB)和蛋白復活因子Bcl-2、Bcl-XL來抑制細胞凋亡(圖5)。近些年也發現β-arrestin的募集也參與了GLP-1R功能[39]。如敲出β細胞中β-arrestin1會誘使cAMP的降低和胰島素釋放減少80%[40],敲出β-arrestin2的野生小鼠會產生餐后高血糖,糖耐性降低并會引起胰島素抵抗等癥狀[41]。研究表明[42,43],β-arrestins主要是通過形成β-arrestins依賴性信號復合物和信號分子(ERK 1/2,JNK3)作為信號載體傳遞信號于MAPK,從而調控β-arrestins依賴性胰島素信號通路。
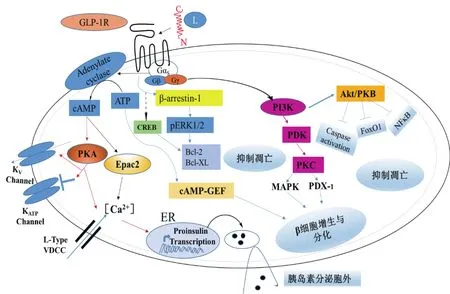
圖5 胰島β細胞中GLP-1R調控的信號通路
2.3 GLP-1R生理功能
GLP-1R作為糖尿病治療的重要靶點。在機體內廣泛表達由于分布于胃、小腸、心臟、腎臟、肺及大腦等組織。在胰島細胞中,GLP-1R主要是促進胰島素的釋放,增加胰島β細胞的再生,抑制β細胞的凋亡,降低胰高血糖素的釋放。在胃腸道等組織中[44],GLP-1R可以通過與其激動劑結合抑制胃腸道的蠕動和胃液分泌,延遲胃的排空,增加飽食感。在神經組織中,小分子GLP-1R激動劑能夠穿透于大腦激活GLP-1R表達的神經元子集,保護神經細胞的凋亡和加強學習記憶能力[45]。不僅如此,GLP-1R也能夠控制食物攝取來減輕體重[46]。在心血管方面,通過對2個月的鼠進行GLP-1R基因的敲出發現其靜息心率降低,左心室舒張壓升高[47];除此之外,GLP-1R還能調控降低氧化應激壓和抑制心肌細胞的凋亡[48]。
3 小分子藥物的篩選
GLP-1R非肽激動劑因具有潛在口服活性能避免T2DM患者長期自我注射,因而引起了人們的廣泛關注。目前,已報道的有關基于GLP-1受體治療糖尿病藥物而開發的小分子先導化合物較多(表1)。
3.1 小分子拮抗劑
事實上,第一個GLP-1R非肽類配體T-0632實際上是一種拮抗劑[49]。相比較125I-Exendin-4(9-39)而言,它同hGLP-1R的IC50為1.2 μmol/L并且在COS-7細胞中能夠抗拒GLP-1誘導產生cAMP。W33S突變會使得這種拮抗劑同受體結合力降低100倍,表明它同ECD的結合位點為Trp33,該氨基酸位于GLP-1受體胞外域α-螺旋中但不參與多肽的結合。盡管這種化合物能夠作為很好的分子工具被用來研究機體中的生理化學特征,但是結合力相對較弱以致不足以用來研究GLP-1受體。Catein是一種天然的多酚類物質[50],能夠作為GLP-1R選擇性的負調控變構調節劑。在鈣離子流動實驗中,Catein不會對激動劑調控信號產生影響。但是在cAMP積累實驗中,Catein卻發揮著GLP-1(7-36)NH2和GLP-1(1-36)NH2的負調控調節劑的作用。例如,Catein會降低由cAMP引起的GLP-1信號傳導效能,但對由GLP-1受體多肽(Exendin-4)引起的非cAMP信號通路不起調控作用[50]。
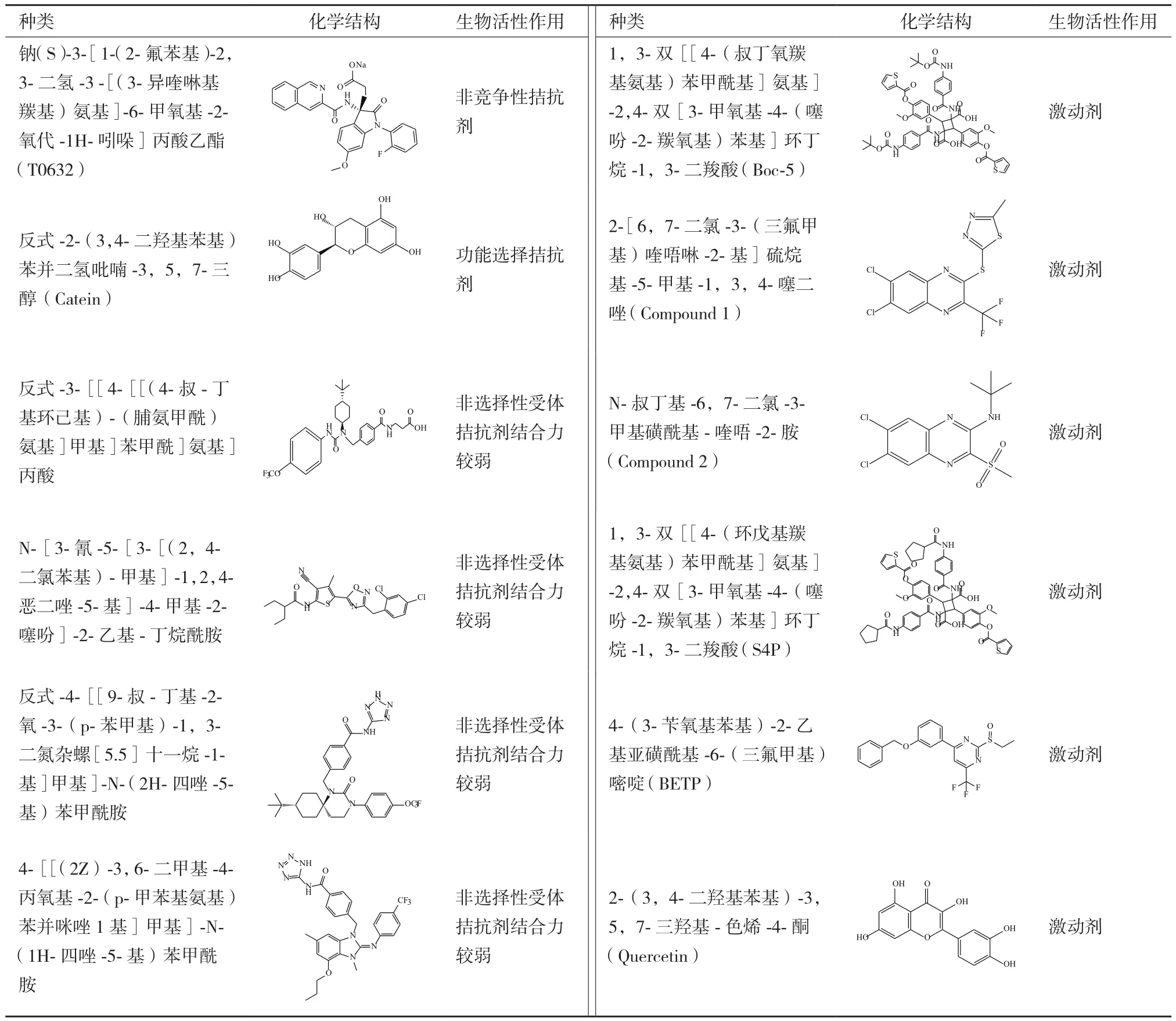
表1 基于GLP-1受體的先導化合物類別[58]
3.2 小分子激動劑
2007年,人們通過高通量篩選方法從多達48 000個的合成和天然化合物中篩選出兩個GLP-1R非肽激動劑,Boc5[51]和它的類似物S4P。它們會在DMSO長期儲存液中自發形成二聚化合物。基于CRE-熒光素酶分析表明,Boc5在HEK-293細胞表達rGLP-1R中扮演著完全激動劑的角色并具有1.08 μmol/L的效能,但是與其結構相似的小分子S4P卻是一種部分激動劑。然而,在cAMP積累實驗中卻發現Boc5和S4P都是部分激動劑。更加重要的是,用Boc5和S4P代替125I-GLP-1進行受體結合實驗也會引起Exendin-4(9-39)功能性拮抗作用。進一步實驗表明,Boc5能夠促進分離的鼠胰島糖依賴性胰島素的分泌,抑制鼠食物攝取并能夠減少db/db小鼠HbA1c含量到正常水平。由于它們缺乏類藥分子所具有的結構特征(背離了Lipinsky和Veber規律),Boc5只是作為一種有用的概念性分子,并且有關該分子結合受體的機理可能同GLP-1類似[52]。2007年發現另外一種非肽類激動劑,compound2(Cmp2)及其類似結構物[53]。這些化合物較Boc5分子量小并且具有不同的藥理學特性。它們是GLP-1R的一種變構調節劑,因為它們不僅能夠獨立發揮調節作用而且也能夠作為變構調節劑加強天然激動劑的活性。然而,這種變構調節劑只是局限于增加GLP-1親和力(26倍)而對其效能卻無任何作用。但是,在hGLP-1R表達的FIpIn-CHO細胞中有助于提高GLP-1的效能[54]。Cmp2還能夠刺激正常小鼠胰島糖依賴性胰島素的釋放但是對于GLP-1R敲出的小鼠卻效用較小。Cmp2和GLP-1對GLP-1R表達的HEK-293細胞產生影響表明這兩種激動劑都是通過Gαs發揮調控作用[55]。進一步研究表明EX4(9-39)會抑制Cmp2調控受體內源化。最新研究表明Cmp2結合GLP-1R會引起受體偶聯Gαs構象改變,但不會引起鈣離子聚集,ERK磷酸化以及受體內源化[56]。黃酮類化合物是另外一種GLP-1R的激動劑。盡管它不會直接激活GLP-1R受體,但可以通過GLP-1和Exendin-4加強鈣離子調控反應。黃銅類結構表明3-OH基團是發揮活性的關鍵基序[57]。
4 展望
已知有大約80%多肽類激素都是依靠激活GPCR信號通路來調控生理機能。因此,GPCR紊亂會引起人類的一些疾病。目前,有大約50%的藥物靶點都屬于GPCR[59]。GPCR B簇成員GLP-1R作為治療2-型糖尿病的重要靶點,對于該受體與配體的相互作用的研究顯得尤為重要。但是,有關配體結合受體核心區的位點目前還是停留在推測階段,受體激活的分子機制尚不明朗。但是有關GPCR B簇受體GCGR,CRF1R[60,61]相繼被解析,這為GLP-1R分子結構作用機理研究提供了較好的借鑒。盡管有關基于GLP-1受體藥物發展很多,但是還不曾有能夠有效口服GLP-1R小分子受體激活劑被用于治療。讓人更加欣慰的是,目前有關小分子GLP-1受體配基的鑒定步伐正在加速。盡管多肽結合機理和GLP-1R受體激活很復雜以致于模擬小分子化合物十分困難,但是通過變構模型促使小分子發揮效用的實例[52]表明針對靶點的藥物化學策略是可行的。因此,運用先進的結構生物方法學和更復雜分析系統及測試方案包括理解受體配體偏向性信號傳導將有可能推進類藥分子的發展。這也有助于為下一代糖尿病藥物研發利用提供參考,具有十分重要的臨床應用價值。
[1] Blad CC, Tang C, Offermanns S. G protein-coupled receptors for energy metabolites as new therapeutic targets[J]. Nature Reviews Drug Discovery, 2012, 11:603-619.
[2] 田洪斌. 注射用艾塞那肽凍干粉針劑的研究[D]. 長春:吉林大學, 2008.
[3] Koole C, Savage EE, Christopoulos A, et al. Minireview:Signal bias, allosterism, and polymorphic variation at the GLP-1R:implications for drug discovery[J]. Molecular Endocrinology, 2013, 27:1234-1244.
[4] Lauritsen KB, Moody AJ, Christensen KC, et al. Gastric inhibitory polypeptide(GIP)and insulin release after small-bowel resection in man[J]. Scandinavian Journal of Gastroenterology, 1980, 15:833-840.
[5] Thorens B, Widmann C. Structure and function of the glucagon-like peptide-1 receptor[M]. Springer Berlin Heidelberg, 1996:255-273.
[6] Brubaker PL, Drucker DJ. Structure-function of the glucagon receptor family of G protein-coupled receptors:the glucagon, GIP, GLP-1, and GLP-2 receptors[J]. Receptors & Channels, 2011, 8:179-188.
[7] Sloop FSW, Kyle W. Physiology and emerging biochemistry of the glucagon-like peptide-1 receptor[J]. Experimental Diabetes Research, 2012, 2012:344-350.
[8] Pabreja K, Mohd MA, Koole C, et al. Molecular mechanisms underlying physiological and receptor pleiotropic effects mediated by GLP-1R activation[J]. British Journal of Pharmacology, 2014, 171:1114-1128.
[9] Parthier C, Reedtz-Runge S, Rudolph R, et al. Passing the baton in class B GPCRs:peptide hormone activation via helix induction?[J]. Trends in Biochemical Sciences, 2009, 34:303-310.
[10] Parthier C, Kleinschmidt M, Neumann P, et al. Crystal structure of the incretin-bound extracellular domain of a G protein-coupled receptor[J]. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104:13942-13947.
[11] G?ke R, Just R, Lankat-Buttgereit B, et al. Glycosylation of the GLP-1 receptor is a prerequisite for regular receptor function[J]. Peptides, 1994, 15:675-681.
[12] Chen Q, Miller LJ, Dong M. Role of N-linked glycosylation in biosynthesis, trafficking, and function of the human glucagon-like peptide 1 receptor[J]. Ajp Endocrinology & Metabolism, 2010, 299:E62-68.
[13] Aiysha T, Venkateswarlu K. The regions within the N-terminus critical for human glucagon like peptide-1 receptor(hGLP-1R)cell surface expression[J]. Scientific Reports, 2014, 4:7410.
[14] Huang Y, Wilkinson GF, Willars GB. Role of the signal peptide in the synthesis and processing of the glucagon-like peptide-1 receptor[J]. Experimental Biology & Medicine, 2007, 67:141-145.
[15] Wilmen A, Goke B, G?ke R. The isolated N-terminal extracellular domain of the glucagon-like peptide-1(GLP)-1 receptor has intrinsic binding activity[J]. Febs Letters, 1996, 398(45):43-47.
[16] Bazarsuren AGU, Wozny M, Reusch D, et al. In vitro folding, functional characterization, and disulfide pattern of the extracellular domain of human GLP-1 receptor[J]. Biophysical Chemistry, 2002, 96:305-318.
[17] Steffen R, Henning TG, Kjeld M, et al. Crystal structure of the ligand-bound glucagon-like peptide-1 receptor extracellular domain[J]. Journal of Biological Chemistry, 2008, 283:11340-11347.
[18] Christina Rye U, Patrick G, Lotte Bjerre K, et al. Crystal structure of glucagon-like peptide-1 in complex with the extracellular domain of the glucagon-like peptide-1 receptor[J]. Journal of Biological Chemistry, 2010, 285:723-730.
[19] Day JW, Li P, Patterson JT, et al. Charge inversion at position 68 of the glucagon and glucagon-like peptide-1 receptors supports selectivity in hormone action[J]. Journal of Peptide Science, 2011, 17:218-225.
[20] Patterson JT, Li P, Day JW, et al. A hydrophobic site on the GLP-1 receptor extracellular domain orients the peptide ligand for signal transduction[J]. Molecular Metabolism, 2013, 2:86-91.
[21] 高蔚豐, 王娟. 保守的第52位色氨酸突變引起的胰高血糖素樣肽1受體N端片段活性喪失[J]. 生物工程學報, 2013, 29:87-94.
[22] Xiao Q, Jeng W, Wheeler MB. Characterization of glucagonlike peptide-1 receptor-binding determinants[J]. Journal of Molecular Endocrinology, 2000, 25:321-335.
[23] Maturana RLD, Dan D. The glucagon-like peptide-1 receptor binding site for the N-terminus of GLP-1 requires polarity at Asp198 rather than negative charge[J]. Febs Letters, 2002, 530:244-248.
[24] Al-Sabah S, Donnelly D. The positive charge at Lys-288 of the glucagon-like peptide-1(GLP-1)receptor is important for binding the N-terminus of peptide agonists-FEBS Letters[J]. Febs Letters, 2003, 553:342-346.
[25] Rakel LDM, Janet TB, Fatima A, et al. Met-204 and Tyr-205 are together important for binding GLP-1 receptor agonists but not their N-terminally truncated analogues[J]. Protein & Peptide Letters, 2004, 11:15-22.
[26] Beinborn M, Worrall CI, Mcbride EW, et al. A human glucagonlike peptide-1 receptor polymorphism results in reduced agonist responsiveness[J]. Regulatory Peptides, 2005, 130:1-6.
[27] Mann RJ, Suleiman AS, Rakel López DM, et al. Functional coupling of Cys-226 and Cys-296 in the glucagon-like peptide-1(GLP-1)receptor indicates a disulfide bond that is close to the activation pocket[J]. Peptides, 2010, 31:2289-2293.
[28] Coopman K, Wallis R, Robb G, et al. Residues within the transmembrane domain of the glucagon-like peptide-1 receptor involved in ligand binding and receptor activation:modelling the ligand-bound receptor[J]. Molecular Endocrinology, 2011, 25:1804-1818.
[29] Jin MM, Hee Young K, Sumi P, et al. Evolutionarily conserved residues at glucagon-like peptide-1(GLP-1)receptor core confer ligand-induced receptor activation[J]. Journal of Biological Chemistry, 2011, 287:3873-3884.
[30] Cassandra K, Denise W, John S, et al. Second extracellular loop of human glucagon-like peptide-1 receptor(GLP-1R)has a critical role in GLP-1 peptide binding and receptor activation[J]. Journal of Biological Chemistry, 2012, 287:3642-3658.
[31] Jin MM, Yoo-Na L, Sumi P, et al. Ligand binding pocket formed by evolutionarily conserved residues in the glucagon-like peptide-1(GLP-1)receptor core domain[J]. Journal of Biological Chemistry, 2015, 290:5696-5706.
[32] Thompson A, Kanamarlapudi V. Distinct regions in the C-Terminus required for GLP-1R cell surface expression, activity and internalisation[J]. Molecular & Cellular Endocrinology, 2015, 413:66-77.
[33] Zhang J, Gu S, Sun X, et al. Computational insight into conformational states of glucagon-like peptide-1 receptor(GLP-1R)and its binding mode with GLP-1[J]. Rsc Advances, 2016, 6:13490-13497.
[34] Yang L, Yang D, De GC, et al. Conformational states of the fulllength glucagon receptor[J]. Nature Communications, 2015, 6:2708-2713.
[35] Hoare SRJ. Mechanisms of peptide and nonpeptide ligand binding to Class B G-protein-coupled receptors[J]. Drug Discovery Today, 2005, 10:417-427.
[36] Dong M, Gao F, Pinon DI, et al. Insights into the structural basis of endogenous agonist activation of family B G protein-coupled receptors[J]. Molecular Endocrinology, 2008, 22:1489-1499.
[37] Holz GG, Leech CA, Heller RS, et al. cAMP-dependent mobilization of intracellular Ca2+stores by activation of ryanodine receptors in pancreatic beta-cells. A Ca2+signaling system stimulated by the insulinotropic hormone glucagon-like peptide-1-(7-37)[J]. Journal of Biological Chemistry, 1999, 274:14147-14156.
[38] Leech CA, Holz GG, Chepurny O, et al. Expression of cAMP-regulated guanine nucleotide exchange factors in pancreatic β-cells[J]. Biochemical & Biophysical Research Communications, 2000, 278:44-47.
[39] Noriyuki S, Takeshi I, Takeshi Y, et al. Beta-Arrestin-1 mediates glucagon-like peptide-1 signaling to insulin secretion in cultured pancreatic beta cells[J]. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105:6614-6619.
[40] Shukla AK, Xiao K, Lefkowitz RJ. Emerging paradigms of β-arrestin-dependent seven transmembrane receptor signaling[J]. Trends in Biochemical Sciences, 2011, 36:457-469.
[41] Luan B, Zhao J, Wu H, et al. Deficiency of a β-arrestin2 signal complex contributes to insulin resistance[J]. Nature, 2009, 457:1146-1149.
[42] Dalle S, Ravier MA, Bertrand G. Emerging roles for β-arrestin-1 in the control of the pancreatic β-cell function and mass:New therapeutic strategies and consequences for drug screening[J]. Cellular Signalling, 2011, 23:522-528.
[43] Etienne K, Ljiljana N, May S, et al. Differential regulation of endosomal GPCR/β-arrestin complexes and trafficking by MAPK[J]. Journal of Biological Chemistry, 2014, 289:23302-23317.
[44] Koliaki C, Doupis J. Incretin-based therapy:a powerful and promising weapon in the treatment of type 2 diabetes mellitus[J]. Diabetes Therapy, 2011, 2:101-121.
[45] During MJ, Cao L, Zuzga DS, et al. Glucagon-like peptide-1 receptor is involved in learning and neuroprotection[J]. Nature Medicine, 2003, 9:1173-1179.
[46] Baggio LL, Drucker DJ. Glucagon-like peptide-1 receptors in the brain:controlling food intake and body weight[J]. Journal of Clinical Investigation, 2014, 124:4223-4226.
[47] Robert G, Xiaomang Y, Baggio LL, et al. Cardiac function in mice lacking the glucagon-like peptide-1 receptor[J]. Endocrinology, 2003, 144:2242-2252.
[48] Laviola L, Leonardini A, Melchiorre M, et al. Glucagon-like peptide-1 counteracts oxidative stress-dependent apoptosis of human cardiac progenitor cells by inhibiting the activation of the c-Jun N-terminal protein kinase signaling pathway[J]. Endocrinology, 2012, 153:5770-5781.
[49] Tibaduiza EC, Chen C, Beinborn M. A small molecule ligand of the glucagon-like peptide 1 receptor targets its amino-terminal hormone binding domain[J]. Journal of Biological Chemistry, 2001, 276:37787-37793.
[50] Denise W, John S, Cassandra K, et al. Modulation of the glucagonlike peptide-1 receptor signaling by naturally occurring and synthetic flavonoids[J]. Journal of Pharmacology & Experimental Therapeutics, 2011, 336:540-550.
[51] Chen D, Liao J, Li N, et al. A nonpeptidic agonist of glucagon-like peptide 1 receptors with efficacy in diabetic db/db mice[J]. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104:943-948.
[52] Lin F, Wang R. Molecular modeling of the three-dimensional structure of GLP-1R and its interactions with several agonists[J]. Journal of Molecular Modeling, 2008, 15:53-65.
[53] Knudsen LB, Kiel D, Teng M, et al. Small-molecule agonists for the glucagon-like peptide 1 receptor[J]. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104:937-942.
[54] Koole C, Wootten D, Simms J, et al. Allosteric ligands of the glucagon-like peptide 1 receptor(GLP-1R)differentially modulate endogenous and exogenous peptide responses in a pathway-selective manner:implications for drug screening[J]. Molecular Pharmacology, 2010, 78:456-465.
[55] Coopman K, Huang Y, Johnston N, et al. Comparative effects of the endogenous agonist Glucagon-Like Peptide-1(GLP-1)-(7-36)amide and the small-molecule ago-allosteric agent "compound 2" at the GLP-1 receptor[J]. Journal of Pharmacology & Experimental Therapeutics, 2010, 334:795-808.
[56] Thompson A, Stephens JW, Bain SC, et al. Molecular characterisation of small molecule agonists effect on the human glucagon like peptide-1 receptor internalisation[J]. PLoS One, 2016, 11:e0154229.
[57] Wootten D, Simms J, Koole C, et al. Modulation of the glucagon-like peptide-1 receptor signaling by naturally occurring and synthetic flavonoids[J]. Journal of Pharmacology & Experimental Therapeutics, 2011, 336:540-550.
[58] Willard FS, Bueno AB, Sloop KW. Small molecule drug discovery at the glucagon-like peptide-1 receptor[J]. Journal of Diabetes Research, 2012, 2012:344-350.
[59] Millar RP, Newton CL. The year in G protein-coupled receptor research[J]. Molecular Endocrinology, 2009, 24:261-274.
[60] Siu FY, He M, de Graaf C, et al. Structure of the human glucagon class B G-protein-coupled receptor[J]. Nature, 2013, 499:444-449.
[61] Hollenstein K, Kean J, Bortolato A, et al. Structure of class B GPCR corticotropin-releasing factor receptor 1[J]. Nature, 2013, 499:438-443.
(責任編輯 馬鑫)
Research Progress on Structure and Function of GLP-1R and Screening for Small Molecule Drugs
HU Zhong-ping CHENG Nian YANG Fan SU Zheng-ding
(Institute of Biomedical and Pharmaceutical Sciences,Hubei Collaborative Innovation Center for Industrial Fermentation,the Key Laboratory of Industrial Fermentation of Ministry of Education,Hubei University of Technology,Wuhan 430068)
Glucagon-like peptide-1 receptor(GLP-1R)as an important target for type 2 Diabetes mellitus(T2DM)therapy,presents clinic significance. The breakthroughs on GLP-1R structures and functions have been made via structural biology and protein engineering. However,it is still unknown on the analysis of its full length structure,the molecular mechanism of polypeptide binding receptors,and the intrinsic mechanism of receptor activation. Owing to the rapid research progresses relating to GLP-1R,this article briefly describes the structure and function of the GLP-1 receptor and the leading compound of existing small molecule drugs,also discusses the developing direction and application prospects of action mechanism of the GLP-1 receptor molecule structure,aiming to provide structure base for the treatment of T2DM.
GLP-1R;molecular structure;small molecule drugs
10.13560/j.cnki.biotech.bull.1985.2017.02.005
2016-05-23
武漢市自然科學基金重點項目(2015060101010033)
胡中平,男,碩士研究生,研究方向:蛋白質結構;E-mail:15623601030@163.com
蘇正定,男,教授,研究方向:蛋白質工程與生物醫藥;E-mail:zhengdingsu@mail.hbut.edu.cn
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
