LINGO-1在脊髓神經(jīng)干細胞向少突膠質(zhì)細胞分化中的作用研究
2017-03-13 01:01:48趙晨光許剛琚芬孫瑋袁華牟翔
中國康復 2017年3期
趙晨光,許剛,琚芬,孫瑋,袁華,牟翔
LRR結(jié)構(gòu)和免疫球蛋白結(jié)構(gòu)域Nogo受體作用蛋白(LRR and Ig Domain Containing,Nogo Receptor Interacting Protein,LINGO-1)是髓鞘源性抑制因子(Myelin-Associated Inhibitory Factors,MAIFs)復合受體中最為重要的組成部分之一[1]。當脊髓損傷后,Nogo-A、髓磷脂相關(guān)糖蛋白(Myelin Associated Glycoprotein,MAG)等髓鞘源性抑制因子大量釋放,并通過與NgR/P75/LINGO-1或NgR/P75/LINGO-1復合受體結(jié)合來介導多種生物效應,在脊髓損傷后的神經(jīng)修復中扮演了極其重要的角色[2]。LINGO-1是由614個氨基酸構(gòu)成的跨膜生物蛋白,由15號染色體編碼而成,定位于15q24.3,其可以通過調(diào)控RhoA途徑,終使生長錐塌陷而使軸突再生受阻[3];也可以參與并下調(diào)表皮生長因子受體(Epidermal Growth Factor Receptor,EGFR),從而影響Akt通路而調(diào)控神經(jīng)元存活[4]。瑞典Frisen小組研究證實在小鼠室管膜區(qū)存在著“靜止”狀態(tài)的神經(jīng)干細胞(Neural Stem Cells, NSCs),在脊髓損傷等特定因素下被激活從而出現(xiàn)增殖進而分化的現(xiàn)象[5],本研究旨在觀察LINGO-1調(diào)控脊髓神經(jīng)干細胞(Spinal Cord Derived Neural Stem Cells,SpNSCs)向少突膠質(zhì)細胞分化及成熟的生物學作用,為進一步調(diào)控其促進脊髓損傷后神經(jīng)修復帶來新思路。
1 材料與方法
1.1 材料 試驗用Fisher344(10周齡)大鼠(160~180g)由第四軍醫(yī)大學實驗動物中心提供,共3只,進行3次生物學試驗重復。細胞培養(yǎng)用DMEM/F12、B27添加劑、N2添加劑、GlutaMAX添加劑及胎牛血清等均為Gibco公司產(chǎn)品;bFGF和EGF購于Peprotech公司,Accutase、Poly-L-Lysine及Laminin購于Sigma公司;LINGO-1、A2B5、O4、髓鞘堿性蛋白(Anti-Myelin Basic Protein,MBP),熒光二抗FITC-conjugated Goat anti-mouse,F(xiàn)ITC-conjugated Goat anti-rabbit及Texas Red-conjugated Goat anti-mouse IgG購自STEMCELL公司。
1.2 方法 ①成年大鼠SpNSCs的分離培養(yǎng):生長10周Fischer344大鼠處死后,椎板減壓暴露脊髓,取C3至T12節(jié)段脊髓置于預冷的Hanks液中,小心用眼科剪剝離硬脊膜,將脊髓剪碎后加入Accutase中于37°C中消化30 min,加入NSCs培養(yǎng)基終止消化,離心后用NSCs培養(yǎng)基重懸,用火焰拋光的巴斯德吸管反復吹打成單細胞懸液并用40μm濾網(wǎng)過濾后,胎盤藍計數(shù),按105/ml密度植于NSCs培養(yǎng)基(DMEM/F12+2%B27+1%N2+20ng/ml EGF+20ng/mlbFGF)中,每3天半量換液一次。生長至第7天時機械吹打法傳代處理。每7天傳代,大于3代后進行實驗。② LINGO-1 shRNA慢病毒表達載體構(gòu)建及包裝由上海吉凱生物基因公司提供。體外培養(yǎng)的大鼠脊髓來源神經(jīng)干細胞分為對照組及干擾組,干擾組通過LINGO-1 shRNA慢病毒感染SpNSCs下調(diào)LINGO-1表達,對照組表達Scramble-shRNA的慢病毒,shRNA插入序列為:Sense5`-TGCTGTAGTCTAGCAGGA-TGACGATCGTTTTGGCCACTGACTGACGATC-GTCACTGCTAGACTA-3 `, Antisense 5 `-CCTG-TAGTCTAGCAGTGACGATCGTCAGTCAGTGG-CCAAAACGATCGTCATCCTGCTAGACTAC-3`。③免疫熒光染色:吸棄培養(yǎng)基用PBS清洗1次以徹底去除殘留的培養(yǎng)基,4%多聚甲醛固定30min后用PBS清洗3次,每次5min;加入0.3%Triton X-100通透5min,PBS清洗2次。加入稀釋好的一抗4℃孵育過夜,次日吸棄一抗,PBS洗3次,每次5min。加入稀釋好的熒光二抗37℃避光孵育2h,PBS洗3次,每次5min,之后滴加DAPI封片劑,室溫孵育30min。鏡下觀察熒光染色的少突膠質(zhì)細胞形態(tài)及數(shù)量,并使用Image-Pro Plus 5.0軟件進行測量分析A2B5、O4陽性細胞和LINGO-1雙染陽性細胞比例。④Westernblot蛋白定量檢測:分別在分化第7天取蛋白,吸棄培養(yǎng)液,用預冷的PBS清洗2次,以徹底去除殘留的培養(yǎng)液。加入預冷的RIPA裂解細胞并轉(zhuǎn)移入預冷的離心管中,離心后將上清轉(zhuǎn)移至預冷的離心管中,用BCA對提取的蛋白進行定量。取蛋白樣品,加入蛋白上樣緩沖液,SDS-PAGE分離蛋白,濕轉(zhuǎn)法將蛋白質(zhì)轉(zhuǎn)移至PVDF膜上。將PVDF膜轉(zhuǎn)移至含有5%脫脂奶粉的TBST封閉液中封閉1h,加入一抗的稀釋液,4℃過夜,次日TBST洗膜3次,每次10min。將PVDF膜放入含稀釋二抗的10cm皿中,孵育1h,在增強化學發(fā)光反應混合液中反應2~3min后,拍片顯影,條帶灰度采用Imege J軟件進行分析MBP蛋白的相對灰度比值。
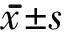
2 結(jié)果
2.1 LINGO-1在少突膠質(zhì)細胞分化過程中表達情況
分別采用少突膠質(zhì)細胞標志物A2B5及O4和LINGO-1抗體進行雙染色。結(jié)果表明在誘導分化后,部分脊髓神經(jīng)干細胞向少突膠質(zhì)細胞分化,細胞有雙極或三極突起,胞體較規(guī)則。LINGO-1抗體(圖1B、F)和A2B5抗體(圖1A)、O4抗體(圖1E)以及細胞核Hoechst染色(圖C、G)顯示分別共表達于培養(yǎng)分化細胞中(圖1D、H),說明LINGO-1在脊髓神經(jīng)干細胞分化成為少突膠質(zhì)細胞的過程中持續(xù)表達。
2.2 LINGO-1促進少突膠質(zhì)細胞分化成熟 為了進一步了解LINGO-1對少突膠質(zhì)細胞分化成熟的影響,我們在分化第7天應用O4抗體對神經(jīng)元進行免疫熒光染色。結(jié)果提示LINGO-1干擾組的少突膠質(zhì)細胞(圖2D-F)比對照組(圖2A-C)的數(shù)量更多,成熟度更高,主要表現(xiàn)在LINGO-1干擾組少突膠質(zhì)細胞分化更為徹底,細胞突起更長,并且有更多的片狀髓鞘結(jié)構(gòu)。定量分析成熟少突膠質(zhì)細胞比例顯示對照組為5.3%,而LINGO-1干擾組則高達17.4%,是對照組細胞的3.28倍,2組比較差異有統(tǒng)計學意義(P<0.05)。
2.3 LINGO-1對MBP蛋白表達的影響 相比對照組,LINGO-1干擾組的MBP表達量明顯升高(圖3),MBP條帶采用Image J軟件進行灰度值分析比較,轉(zhuǎn)化為數(shù)值進行量倍比較,進一步半定量分析顯示LINGO-1干擾組的MBP表達量為對照組的5.31倍(56.8±3.3、0.7±0.5,P<0.05)。

A.A2B5 B.LINGO-1 C.Hoechst D.Merge

E.O4 F.LINGO-1 G.Hoechst H.Merge
圖1LINGO-1在少突膠質(zhì)細胞分化過程中表達情況。LINGO-1抗體(B,F)與A2B5抗體(A)、O4抗體(E)以及細胞核Hoechst染色(圖C、G)共表達在培養(yǎng)細胞中(D,H)(×40)。

A.O4 B.Hoechst C.Merge

D.O4 E.Hoechst F.Merge
圖2LINGO-1對少突膠質(zhì)細胞分化成熟的影響:熒光倒置顯微鏡下觀察LINGO-1干擾組(D-F)和對照組(A-C)少突膠質(zhì)細胞的成熟狀況,綠色細胞表示O4陽性細胞(×40)。

圖3 LINGO-1對MBP蛋白表達的影響
3 討論
本實驗中成功從成年大鼠脊髓中提取培養(yǎng)了神經(jīng)干細胞,誘導分化后利用免疫熒光染色發(fā)現(xiàn)LINGO-1分別與A2B5及O4共表達,證實LINGO-1表達于少突膠質(zhì)細胞成熟的各階段。然后利用RNAi對LINGO-1的表達進行下調(diào),發(fā)現(xiàn)下調(diào)LINGO-1的表達可以促進脊髓神經(jīng)干細胞向少突膠質(zhì)細胞分化,并同時促進其成熟,進一步檢測成熟少突膠質(zhì)細胞表達的MBP蛋白時也發(fā)現(xiàn)LINGO-1干擾組MBP蛋白表達量明顯高于對照組。以上實驗結(jié)果為脊髓損傷后早期干預促進內(nèi)源性脊髓神經(jīng)干細胞向少突膠質(zhì)細胞分化并促進其成熟提供活體體外證據(jù),也為進一步促進脊髓損傷后神經(jīng)修復帶來希望。
脊髓損傷后大量神經(jīng)元壞死、神經(jīng)傳導束崩解,少突膠質(zhì)細胞包裹軸突形成的髓鞘分解,形成大量的髓鞘源性抑制因子(Myelin-Associated Inhibitory Factors,MAIFs),這其中包括Nogo-A、MAG和OMgp為代表抑制因子,其中Nogo-A通過其復合受體之一LINGO-1(LRR and Ig Domain Containing,Nogo Receptor Interacting Protein)介導了多種生物學效應,這包括阻止軸突再生及影響神經(jīng)元存活[6-7]。脊髓損傷后位于中央水管周圍的具有多種分化潛能的神經(jīng)干細胞被激活,但是大部分卻分化成為星形膠質(zhì)細胞,所以如何調(diào)控其分化便成為目前神經(jīng)修復研究的熱點問題。
本實驗中我們發(fā)現(xiàn)分離培養(yǎng)的脊髓神經(jīng)干細胞經(jīng)誘導分化后,LINGO-1分別與少突膠質(zhì)細胞標志物A2B5及O4共表達于分化細胞中。A2B5及O4同為少突膠質(zhì)細胞特異性標志物,但是二者的表達時間卻不盡相同[8]。A2B5在少突膠質(zhì)細胞的早期表達,即在少突膠質(zhì)-2型星形膠質(zhì)細胞(Oligodendrocyte-Type2 Astrocyte Progenitor,O-2A Progenitor)及成少突膠質(zhì)細胞中表達,而O4的表達則較為靠后,分別在成少突膠質(zhì)細胞、未成熟少突膠質(zhì)細胞及成熟少突膠質(zhì)細胞中表達。本實驗中發(fā)現(xiàn)LINGO-1可以分別與以上2個標志物分別共表達,這說明LINGO-1在由脊髓神經(jīng)干細胞分化為成熟少突膠質(zhì)細胞的整個過程中持續(xù)表達,也提示其可能會在這一過程中持續(xù)發(fā)揮重要作用。
為了驗證LINGO-1對少突膠質(zhì)細胞分化的作用,我們進一步下調(diào)LINGO-1的表達,結(jié)果發(fā)現(xiàn)干預后少突膠質(zhì)細胞分化成熟比例有明顯提高,達到17.4%,是對照組的3倍之多。此外LINGO-1干擾組的少突膠質(zhì)細胞還表現(xiàn)為細胞突起更長,并且有更多的片狀髓鞘結(jié)構(gòu)。L??v[9]也檢測過LINGO-1對皮層來源NSCs向少突膠質(zhì)細胞分化的影響,其發(fā)現(xiàn)下調(diào)LINGO-1后,少突膠質(zhì)細胞分化成熟有輕微的升高,但是并無統(tǒng)計學意義,這可能與不同來源的NSCs的本身特性有一定關(guān)系。
MBP蛋白是成熟少突膠質(zhì)細胞中表達的蛋白,是構(gòu)成髓鞘的主要成分之一,其表達量的多少可以間接反映出少突膠質(zhì)細胞的成熟情況,試驗中LINGO-1干擾組的MBP表達量明顯升高,為對照組的5倍之多,這與前面的實驗結(jié)果是一致的,都反應出LINGO-1對NSCs向少突膠質(zhì)細胞分化成熟的負性調(diào)控作用。
本研究沒有進一步檢測LINGO-1影響少突膠質(zhì)細胞成熟后對軸突成髓鞘作用的影響及分化成熟可能涉及的通路,這也是本研究下一步所努力的方向。明確LINGO-1在脊髓NSCs分化過程中在少突膠質(zhì)細胞中的表達特征及其對少突膠質(zhì)細胞分化成熟的影響可以使我們更加廣泛的了解LINGO-1的生物學功能,也為進一步促進脊髓損傷后神經(jīng)修復提供可能的作用靶點,為治療脊髓損傷帶來更多的希望。
[1] Mi S, Lee X, Shao Z, et al. LINGO-1 is a component of the Nogo-66 receptor/p75 signaling complex[J]. Nat Neurosci, 2004, 7(3): 221-228.
[2] GrandPré T, Nakamura F, Vartanian T, et al. Identification of the Nogo inhibitor of axon regeneration as a Reticulon protein[J]. Nature, 2000, 403(6768): 439-444.
[3] Ji B, Li M, Wu WT, et al. LINGO-1 antagonistpromotes functional recovery and axonal sprouting after spinal cord injury[J]. Mol Cell Neurosci, 2006, 33(3): 311-320.
[4] Zhao XH, Jin WL, Wu J, et al. Inactivation of glycogen synthase kinase-3beta and up-regulation of LINGO-1 are involved in LINGO-1 antagonist regulated survival of cerebellar granular neurons[J]. Cell MolNeurobiol, 2008, 28(5): 727-735.
[5] Sabelstr?m H, Stenudd M, Frisén J. Neural stemcells in the adult spinal cord[J]. ExpNeurol, 2014, 260(1): 44-49.
[6] Mi S, Hu B, Hahm K, et al. LINGO-1 antagonist promotes spinal cord remyelinationand axonal integrity in MOG-induced experimentalautoimmune encephalomyelitis[J]. Nat Med, 2007, 13(10): 1228-1233.
[7] WangF, Zhu Y. Theinteraction of Nogo-66 receptor with Nogo-P4 inhibits the neuronal differentiation of neural stem cell[J]. Neuroscience, 2008, 151(1): 74-81.
[8] Armstrong R. Isolation and Characterization of Immature Oligodendrocyte Lineage Cells[J]. Methods, 1998, 16(3): 282-292.
[9] L??v C, Fernqvist M, Walmsley A, et al. Neutralization of LINGO-1 during in vitro differentiation of neural stem cells results in proliferation of immature neurons[J]. PLoS One, 2012, 7(1): 29771-29789.
