雷公藤內酯醇對肝細胞色素P450的誘導及機制研究
2017-03-20 03:17:07萬子衿朱燕萍徐海榮
中國藥理學通報 2017年3期
關鍵詞:研究
虞 茜,萬子衿,朱燕萍,徐海榮,廖 凱,劉 峰,李 巍
(1. 揚州大學醫學院,2. 江蘇省中西醫結合老年病防治重點實驗室,江蘇 揚州 225001)
雷公藤內酯醇對肝細胞色素P450的誘導及機制研究
虞 茜1,2,萬子衿1,2,朱燕萍1,2,徐海榮1,2,廖 凱1,2,劉 峰1,李 巍1,2
(1. 揚州大學醫學院,2. 江蘇省中西醫結合老年病防治重點實驗室,江蘇 揚州 225001)
目的 探索雷公藤內酯醇(triptolide,TP)對肝細胞中細胞色素P450(CYPs)的誘導及可能機制。方法 TP處理大鼠原代肝細胞或HepG2細胞后,采用實時定量PCR和Western blot等方法檢測CYPs的表達水平變化,并結合特異性抑制劑或基因敲低的方法分析TP誘導CYPs的可能機制。結果 TP能夠分別誘導大鼠CYP1A2、2C7、2C11、2C12、2D2、2E1和3A1的mRNA表達,50 nmol·L-1時誘導倍數分別為18.5、2.2、31.2、3.2、21.5、88.3、34.0;100 nmol·L-1時誘導倍數分別為20.3、4.6、29.6、23.1、61.1、83.0、38.5。HepG2細胞中人CYP1A1、2B6、2C9、2C19、2D6、2E1和3A4也被TP誘導。大鼠原代肝細胞內核受體PXR和CAR的活性被TP下調;TP可上調肝細胞內p53的蛋白水平,p53活性抑制后TP對部分CYPs的誘導作用受到抑制。結論 TP可在肝細胞水平誘導CYPs表達,該誘導作用可能不通過核受體,p53可能參與了TP對部分CYPs的誘導作用。
雷公藤內酯醇;細胞色素P450;核受體;p53;肝細胞;HepG2
雷公藤內酯醇(triptolide, TP)是中國傳統中藥雷公藤(Tripterygiumwilfordii)的主要活性物質,具有抗炎、抗腫瘤和免疫抑制等多種藥理活性[1]。采用大鼠的整體實驗研究表明,TP可誘導大鼠體內幾種CYPs[2]。而體外肝細胞模型能否復制體內TP對CYPs的誘導作用,另一重要的代謝酶亞家族CYP2D能否被誘導,以及TP對CYPs的誘導作用機制未見報道。因此,本研究對大鼠原代肝細胞在TP作用后CYPs、核受體及轉錄因子p53的表達進行了分析。此外,為考察TP對CYPs的誘導作用是否可能在人體發生,本研究對人源的HepG2肝癌細胞在TP作用下CYPs的表達變化進行了分析。
1 材料與方法
1.1 試劑 雷公藤內酯醇(四川維克奇生物制品有限公司,純度≥98%,溶于二甲基亞砜DMSO),I型鼠尾膠原蛋白(生友生物技術有限公司),膠原酶IV型(Sigma),HBSS(+)和HBSS(-)(上海生工),Percoll(Pharmacia),0.4%臺盼藍染液(Solarbio),7-乙基香豆素(7-EC,Sigma),T4 Ligase(New England BioLAB),感受態大腸桿菌(DH5α,寶生物公司),反轉錄試劑盒(HiScript Q RT SuperMix for qPCR,Vazyme),Taqman法實時定量PCR試劑盒(FastStart Universal Probe Master,Roche),Taqman探針(Life technologies),SYBR Green法實時定量PCR試劑盒(FastStart Essential DNA Green Master,Roche),鼠抗CYP3A4(Millipore),兔抗p53(Proteintech),兔抗CAR(Proteintech),兔抗PXR(Proteintech),鼠抗GAPDH(KANGCHEN),山羊抗鼠IgG、山羊抗兔IgG(北京中杉金橋生物技術有限公司)。
1.2 儀器 CO2培養箱(Thermo),FluorChem E多功能成像分析系統(Proteinsimple公司),熒光酶標儀(Biotech公司),LC/MS/MS(AB公司),電泳系統(Biored公司),實時定量PCR儀(Roche公司)。
1.3 動物 清潔級Wistar大鼠,♂,體質量(195±15)g,購自揚州大學醫學比較中心[動物合格證號:SCXK(蘇)2012-0004]。
1.4 大鼠原代肝細胞的分離與培養 大鼠原代肝細胞采取兩步膠原酶灌注法分離。Wistar大鼠用質量分數為1%的水合氯醛(ip)麻醉后暴露腹腔,門靜脈流入37 ℃灌注液(NaCl 0.142 mol·L-1、KCl 6.7 mmol·L-1、HEPES 0.01 mol·L-1、EDTA 0.05 mmol·L-1,pH=7.4,流速10 mL·min-1),同時剪開腹靜脈。肝臟灌注至黃白色后換成膠原酶溶液[0.5 g·L-1溶于HBSS(+),37 ℃]灌注15 min。移出肝臟用HBSS(-)清洗,于HBSS(-)中去除肝葉表面的膜并輕挑肝臟,采用200目細胞篩過濾細胞懸液至50 mL離心管中。4 ℃、126×g離心細胞懸液1 min兩次。用體積分數為28%的Percoll重懸細胞后4 ℃、126×g離心10 min,移去上清。用DMEM培養基(含有10%胎牛血清、100 kU·L-1青霉素和100 mg·L-1鏈霉素、MEM NEAA 2 mmol·L-1、HEPES 10 mmol·L-1)重懸細胞,0.4%臺盼藍染液1 ∶1稀釋后細胞計數,每孔4×105接種于Ⅰ型鼠尾膠原蛋白包被的6孔板。于37 ℃、5% CO2培養箱中培養12 h后用TP (0、50、100 nmol·L-1)處理,每12 h更換含藥培養基。
1.5 p53敲低的HepG2細胞的建立 由于HepG2細胞直接轉染效率較低,采用慢病毒建立p53敲低的HepG2細胞株。合成p53 shRNA編碼序列(序列在Sigma mission shRNA的基礎上修改所得,見Tab 1)后進行退火反應,形成帶有粘性末端的雙鏈序列。采用T4 Ligase將上述退火產物與BbsI酶切后的MSCV-puro-BbsI進行連接反應,連接產物轉化至感受態大腸桿菌,挑選陽性克隆并測序鑒定,獲得質粒分別命名為MSCV-puro-532和MSCV-puro-533。分別將上述構建好的質粒或陰性對照MSCV-puro-SCR與pCMV-Gag-Pol 和pCMV-VSV-G(DNA含量分別為1.8、0.6、0.6 μg) 共轉染至293T細胞,24 h后使用293T細胞培養基感染HepG2細胞。培養48 h后更換含2 mg·L-1嘌呤霉素的培養基,再培養72 h,之后用1 mg·L-1嘌呤霉素維持培養,獲得的細胞命名為HepG2-532、HepG2-533和HepG2-SCR。
1.6 熒光實時定量PCR反應 TP處理大鼠原代肝細胞或HepG2細胞48 h后,TRIzol法提總RNA進行反轉錄。Taqman探針法檢測大鼠CYP1A2、2B1、2D2、2E1、3A1和3A9的mRNA的表達變化。SYBR Green法檢測大鼠的CYP2C6、2C7、2C12、PXR和CAR,以及人CYP1A1、2B6、2C9、2C19、2E1和3A4的mRNA的表達變化,SYBR Green引物序列見Tab 1。
1.7 Western blot檢測蛋白表達水平 TP(0,50和100 nmol·L-1)處理大鼠原代肝細胞后,RIPA裂解液提取總蛋白,BCA法測蛋白濃度,以50 μg上樣,電泳轉膜,封閉1 h,分別加一抗CYP3A4(1 ∶1 000)、p53(1 ∶1 000)、CAR(1 ∶1 000)、PXR(1 ∶1 000)和GAPDH(1 ∶1 000)于4 ℃孵育過夜,洗膜后加二抗,室溫孵育1 h。ECL顯色,用FluorChem E多功能成像分析系統成像。
1.8 CYPs活性檢測 全CYPs活性采用7-EC檢測。大鼠原代肝細胞2×105接種于12孔板,用TP處理72 h后,更換含7-乙基香豆素的培養基繼續孵育30 min,收集培養基,剩余細胞采用BCA法測蛋白濃度。培養基加等體積乙醇沉淀蛋白,13 000×g離心后,采用熒光酶標儀,于350 nm下激發,檢測450 nm發射光,該結果采用每孔蛋白含量進行標準化。大鼠原代肝細胞2×106接種于培養皿,采用TP(0、50、100 nmol·L-1)處理72 h后,細胞刮刀取全部細胞離心并用PBS清洗兩次,細胞重懸于磷酸鉀緩沖液(100 mmol·L-1,pH 7.4)中,玻璃勻漿器研磨后于9 000×g離心,所得上清為S9。睪酮(250 μmol·L-1)或丁呋洛爾(70 μmol·L-1)分別與含MgCl2(5 mmol·L-1)、NADPH(1 mmol·L-1)和S9(0.225 g·L-1)的磷酸鉀緩沖液(100 mmol·L-1,pH 7.4)于37 ℃下孵育30 min,反應產物6β-羥基睪酮和1-羥基丁呋洛爾采用LC/MS/MS檢測[3]。
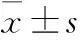

Tab 1 Primers used in the real-time quantitative PCR reactions and p53 shRNA
2 結果
2.1 TP誘導大鼠原代肝細胞中CYPs的表達和活性 TP處理大鼠原代肝細胞后,qRT-PCR檢測CYPs mRNA的變化。該結果與整體實驗的趨勢相似[2],TP可誘導大鼠原代肝細胞內的CYP1A2、2C11、2E1、3A1的mRNA的表達,50 nmol·L-1下的誘導倍數分別為18.5、31.2、88.3、34.0,100 nmol·L-1下的誘導倍數分別為20.3、29.6、83.0、38.5(Fig 1A~1D)。該結果提示,大鼠原代肝細胞可能成為研究TP對CYPs誘導的體外模型。應用該模型,研究發現CYP2C7、2C12和2D2的mRNA水平也被TP誘導,50 nmol·L-1下的誘導倍數分別為2.2、3.2和21.5,100 nmol·L-1下的誘導倍數分別為4.6、23.1和61.1(Fig 1E~1G)。Western blot實驗結果顯示,TP可導致CYP3A的蛋白水平上調(Fig 2),提示了蛋白上調與mRNA上調一致。
大鼠原代肝細胞在TP處理72h后,采用全CYPs底物7-EC共孵育,7-EC的生成速率明顯上調,提示TP可能誘導大鼠原代肝細胞內的CYPs活性(Fig 3A)。制備S9進行體外代謝反應,結果與整體實驗趨勢一致[2],TP導致6β-羥基睪酮的生成速率加快(Fig 3B),提示該體外原代肝細胞模型可作為TP對CYPs誘導的研究模型。由于本研究首次報道CYP2D2的mRNA水平被誘導,因此對TP處理大鼠原代肝細胞后的S9代謝丁呋洛爾為1-羥基丁呋洛爾的代謝速率進行分析,發現1-羥基丁呋洛爾的生成速率上升約1.7倍(Fig 3C),提示TP在表達和酶活性水平均可誘導CYP2D2。
2.2 核受體CAR和PXR的表達 核受體是CYPs表達調控的重要分子,因此采用qRT-PCR及Western blot對TP作用后核受體CAR和PXR的表達量進行分析。結果顯示在TP作用后CAR和PXR的mRNA水平下調(Fig 4A、4B),50 nmol·L-1TP處理下,CAR和PXR的mRNA水平分別為對照組的0.8和0.3倍,而100 nmol·L-1TP處理后分別為0.2和0.1倍,但是蛋白水平變化不明顯(Fig 2)。
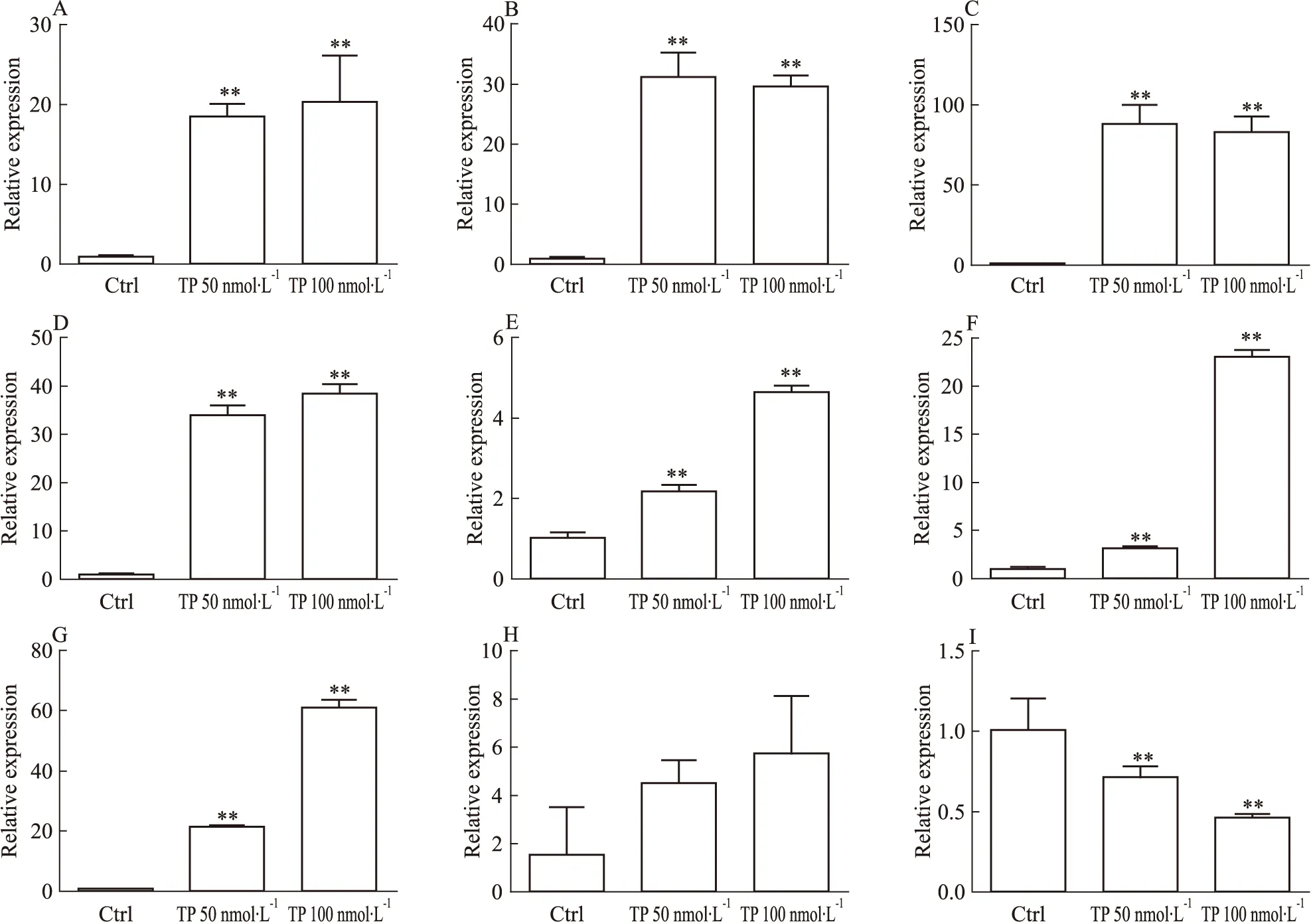
Fig 1 Changes of mRNA level of CYP1A2(A),CYP2C11(B),CYP2E1(C),CYP3A1(D),CYP2C7(E),CYP2C12(F),CYP2D2(G),CYP2B1(H) and CYP3A9(I) in rat primary hepatocytes after triptolide treatment
**P<0.01vscontrol group
Fig 2 Western blot results(A) and relative band density of CYP3A(B), PXR(C), CAR(D) andp53(E) in rat primary hepatocytes after triptolide treatment
該結果提示TP可能不通過誘導核受體CAR和PXR而提高CYPs的表達,但由于核受體表達量的變化不能完全反映其活性,而其下游靶基因的表達水平可間接反映核受體活性。為進一步檢測PXR與CAR的活性的可能變化,我們檢測了其下游靶基因P-糖蛋白(P-gp)的mRNA水平。結果表明,TP作用下P-gp的mRNA水平下調(Fig 4C),在50、100 nmol·L-1的TP作用下,分別為對照組的0.6和0.4倍,提示了PXR和CAR活性水平的下調。該結果表明,TP可能不活化核受體PXR和CAR,因此核受體可能并不參與TP對CYPs的誘導。
2.3 轉錄因子p53的表達 據報道p53可能誘導CYPs的轉錄調控,進而導致CYPs活性上調[4-6]。在50、100 nmol·L-1的TP處理CYPs后,p53蛋白水平上調(Fig 2)。為進一步檢測p53在TP調控CYPs中的作用,將p53抑制劑Pifithrin-α hydrobromide與TP共同作用于大鼠原代肝細胞后,提取總RNA并檢測CYPs的mRNA水平變化。結果顯示,100 nmol·L-1的TP對大鼠CYP1A2、2D2、2E1和3A1誘導倍數分別為100.5、130.6、426.4、420.0倍;加入p53抑制劑后,誘導倍數分別為17.2、22.3、111.1、22.0。該結果表明p53抑制后,TP對細胞內CYP1A2、2D2、2E1和3A1的誘導能力降低。但p53抑制劑對TP誘導CYP2C7的作用無明顯影響。p53抑制劑導致TP對CYP2C11、2C12的誘導能力提高(Fig 5),100 nmol·L-1的TP對上述兩酶的誘導倍數分別為29.6和13.0,加入p53抑制劑后,誘導倍數分別為45.9和34.8。該結果提示,p53可能在TP對CYPs誘導作用中發揮作用。
2.4 TP誘導人肝癌細胞HepG2細胞株中CYPs表達 由于人與大鼠CYPs的誘導調控機制可能存在種屬的差異,因此本研究采用HepG2細胞以考察TP是否也可通過p53信號途徑誘導CYPs的表達。在HepG2-SCR 細胞中,50 nmol·L-1TP誘導CYP1A1、2B6、2C9、2C19、2D6、2E1和3A4的mRNA表達,分別為48.0、20.8、114.0、67.8、36.4、6.2和21.0倍。因為HepG2細胞中核受體PXR和CAR表達極低或缺失[8-9],該結果提示在人源肝細胞模型中,TP對CYPs的誘導作用可能也可不通過這兩個核受體。在p53敲低的HepG2-532和HepG2-533細胞中,CYP2C9和CYP3A4的誘導在兩株細胞中均被抑制(Fig 6),提示TP在HepG2細胞中對這兩個酶的誘導作用可能通過p53。
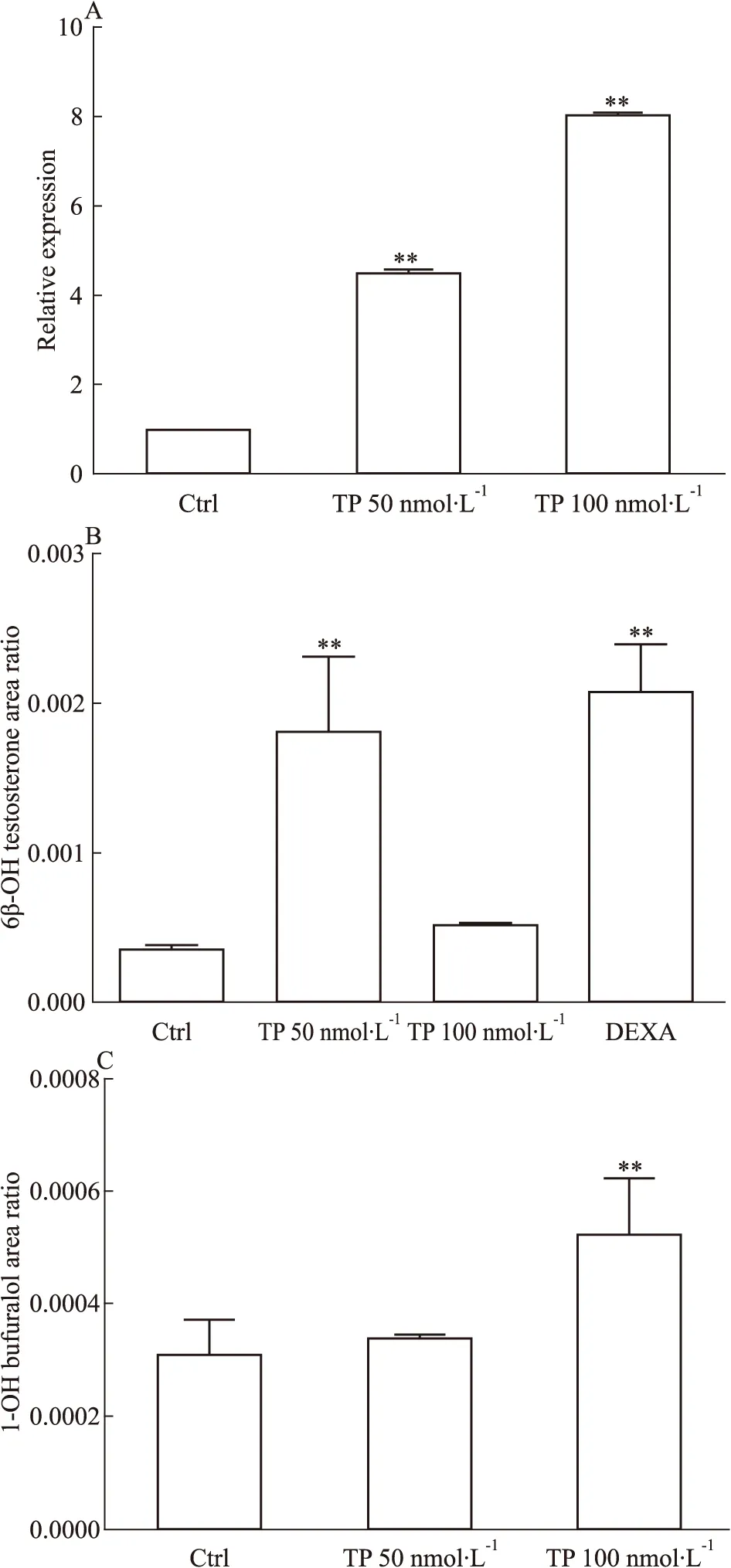
Fig 3 Changes of metabolic rate of 7-EC(A),testosterone(B) and bufuralol(C) in rat primary hepatocytes after triptolide treatment
**P<0.01vscontrol group
3 討論
前期研究表明,TP灌胃后大鼠肝內的CYP1A2、2C11、2E1、3A1可被誘導[2]。但體內CYPs的調控可能是復雜的過程,其表達可能與體內的微環境變化相關,包括內源性的氧化類固醇、膽汁酸、雄激素、孕激素、腎上腺素和炎癥因子等。為進一步確認TP對CYPs的誘導作用是否可在肝細胞水平上發生或依賴于體內環境的變化,本研究采用大鼠原代肝細胞對TP的誘導作用進行了檢測。結果發現大鼠原代肝細胞內CYP1A2、2C11、2E1、3A1的表達均被TP誘導,與體內實驗結果一致。因此本研究結果提示,大鼠原代肝細胞可成為TP對CYPs誘導研究的體外模型。應用大鼠原代肝細胞的模型,本研究發現TP也可誘導CYP2C7、2C12、2D2的表達。應用HepG2細胞的實驗結果表明,TP可誘導人CYP1A1、2B6、2C9、2C19、2D6、2E1、3A4的表達。結果表明,TP對CYPs的誘導作用可發生在肝細胞水平,大鼠原代肝細胞和人HepG2細胞均可成為研究TP對CYPs的誘導機制的體外模型。
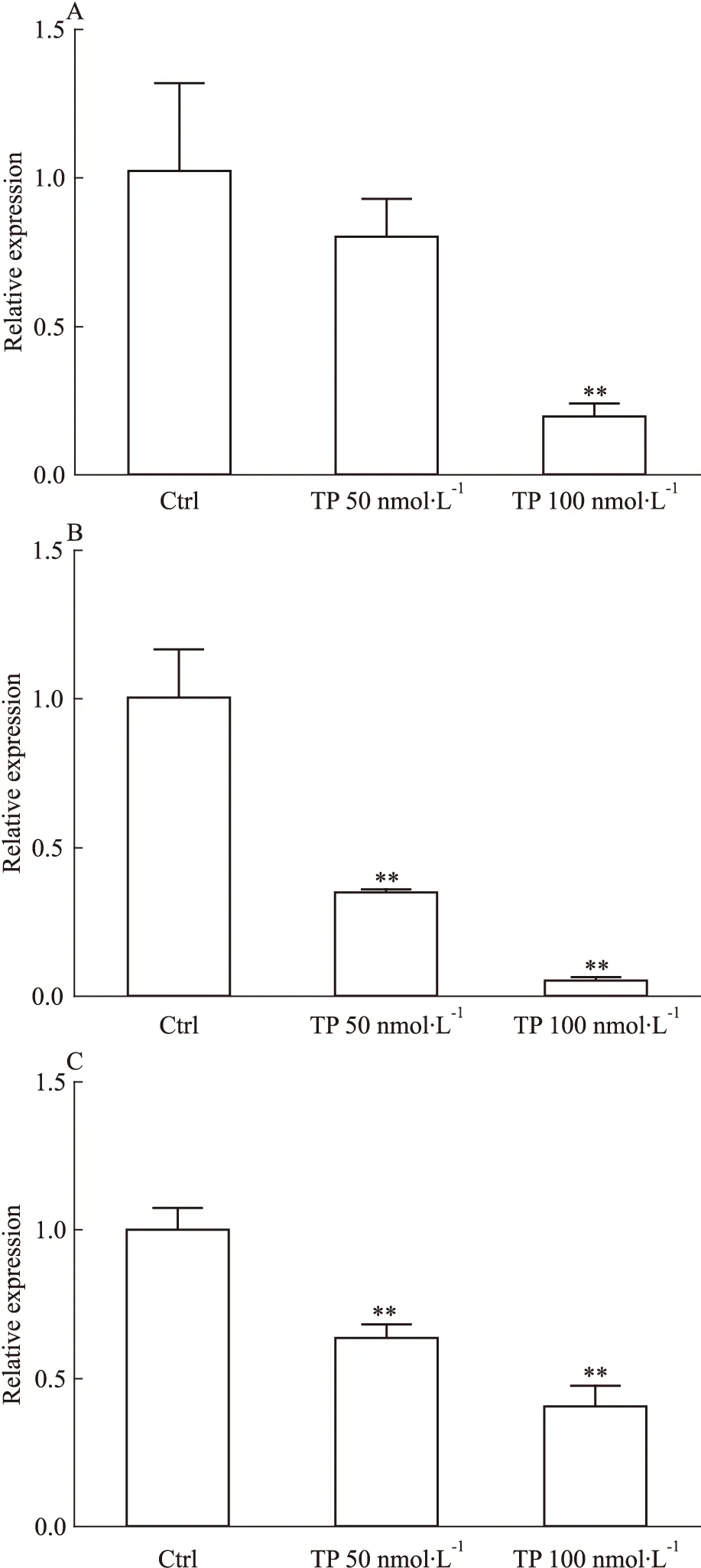
Fig 4 Changes of mRNA level of CAR(A),PXR(B) and P-gp(C) in rat primary hepatocytes after triptolide treatment
**P<0.01vscontrol group
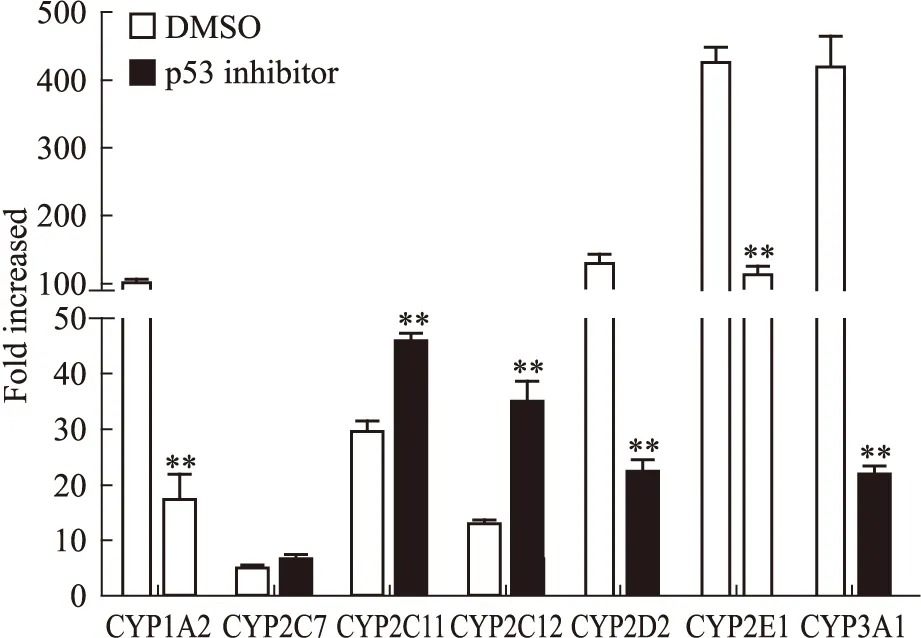
Fig 5 Changes of mRNA level of CYP1A2, CYP2D2, CYP2E1, CYP3A1, CYP2C7, CYP2C11 and CYP2C12 when rat primaryhepatocytes were co-treated with triptolide(100 nmol·L-1) andp53 inhibitor(pifithrin-α hydrobromide, 20 μmol·L-1)
**P<0.01vsDMSO group
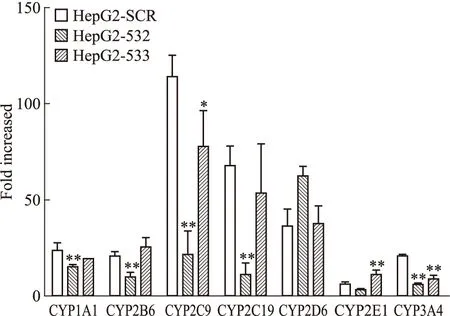
Fig 6 Changes of mRNA level of CYP1A1, CYP2B6, CYP2C9,CYP2C19, CYP2D6, CYP2E1 and CYP3A4 inHepG2-SCR, HepG2-532 and HepG2-533 cellsafter 50 nmol·L-1 triptolide treatment
**P<0.01vsHepG2-SCR group
據報道,TP能夠被人和大鼠體內CYP3A代謝為低毒的羥基化產物[7],并可能抑制CYPs的活性,提示TP與其他藥物共服可能誘導藥藥相互作用(DDI)。但值得注意的是,TP對CYP1A2和CYP3A4的IC50分別為14.18、8.36 μmol·L-1[8],但在服用雷公藤多苷片的個體中,Cmax約在182~441 nmol·L-1[9],因此在正常用藥的情況下,發生直接基于TP抑制代謝酶的DDI的可能性較低。而本研究發現在50~100 nmol·L-1的濃度下,TP可誘導CYPs在大鼠原代肝細胞和人肝癌細胞中的CYPs表達,包括對臨床藥物代謝居前3位的CYP3A、2D、2C。鑒于雷公藤常與非甾體抗炎藥(主要代謝酶為CYP2C)聯合用于治療類風濕性關節炎[10],本研究結果提示TP與非甾體類抗炎藥共服,產生基于代謝酶誘導的DDI的可能性可能高于基于代謝酶抑制的DDI。
核受體是CYPs調節的關鍵分子,NR1I家族的PXR和CAR可誘導包括CYP1、CYP2和CYP3家族在內的多種CYPs的表達[11-12]。但在本研究中,TP作用后PXR和CAR的蛋白表達水平均無明顯變化,且這兩個核受體下游基因P-gp的表達水平也下調,表明PXR和CAR的活性下調;同時TP可誘導缺乏PXR和CAR這兩種核受體表達的HepG2細胞中的CYPs[13-14],該結果表明,TP對CYPs的誘導作用可能不依賴核受體PXR和CAR的作用。
據報道,p53通過結合p53反應元件誘導CYP1A1、CYP2A6和CYP3A4基因的表達[4-6]。我們前期研究表明在LNCaP細胞內TP作用可以上調p53蛋白[15]。在大鼠原代肝細胞中,我們發現TP作用后p53的蛋白水平也發生了上調,進一步采用p53抑制劑后,CYP1A2、2D2、2E1、3A1的上調作用被抑制。而在HepG2細胞中,CYP3A4和2C9的上調也被抑制。該結果提示,TP對部分CYPs的誘導可能是通過p53發生的,但不同種屬間可能存在調控差異。
綜上,本研究發現大鼠原代肝細胞可能成為研究TP對CYPs的誘導作用的體外模型;TP可在nmol·L-1級的濃度下誘導大鼠原代肝細胞和人肝癌細胞HepG2中CYPs的表達,其對部分CYPs的誘導作用可能是通過核受體p53的上調實現的,核受體PXR和CAR可能在該過程中不發揮作用;TP對CYPs的誘導作用可能引發同服藥物的相互作用的可能性。
(致謝:感謝轉化醫學實驗中心的老師和同學技術指導和幫助。)
[1] 張 軍,陳 玟,劉史佳,等.LC/APCI/MS/MS測定Beagle犬口服雷公藤片后血漿中雷公藤甲素[J].中國藥理學通報,2013,29(12):1765-8.
[1] Zhang J, Chen M, Liu S J, et al. LC/APCI/MS/MS analysis for plasma concentration of triptolide in Beagle dogs following oral administration of tripterygium tablets[J].ChinPharmacolBull, 2013, 29(12):1765-8.
[2] Han F M, Peng Z H, Wang J J, Chen Y.Invivoeffect of triptolide combined with glycyrrhetinic acid on rat cytochrome P450 enzymes[J].YaoXueXueBao,2013,48(7):1136-41.
[3] Yu C, Shin Y G, Kosmeder J W, et al.Liquid chromatography/tandem mass spectrometric determination of inhibition of human cytochrome P450 isozymes by resveratrol and resveratrol-3-sulfate[J].RapidCommunMassSpectrom, 2003, 17(4) :307-13.
[4] Wohak L E, Krais A M, Kucab J E, et al.Carcinogenic polycyclic aromatic hydrocarbons induce CYP1A1 in human cells via a p53-dependent mechanism[J].ArchToxicol, 2016, 90(2):291-304.
[5] Goldstein I, Rivlin N, Shoshana OY, et al.Chemotherapeutic agents induce the expression and activity of their clearing enzyme CYP3A4 by activating p53[J].Carcinogenesis, 2013, 34(1):190-8.
[6] Hu H, Yu T, Arpiainen S, et al. Tumour suppressor protein p53 regulates the stress activated bilirubin oxidase cytochrome P450 2A6[J].ToxicolApplPharmacol, 2015, 289(1):30-9.
[7] Li W, Liu Y, He Y Q, et al.Characterization of triptolide hydroxylation by cytochrome P450 in human and rat liver microsomes[J].Xenobiotica, 2008, 38(12):1551-65.
[8] Zhang H, Ya G, Rui H. Inhibitory effects of triptolide on human liver cytochrome P450 enzymes and P-glycoprotein[J].EurJDrugMetabPharmacokinet, 2016.[Epub ahead of print]
[9] 李 穎,汪永忠,羅 歡,等. 類風濕關節炎患者雷公藤甲素血清濃度測定及其藥動學研究[J]. 中國中醫藥信息雜志,2014,21(1):85-7.
[9] Li Y, Wang Y Z, Luo H, et al. Determination of serum concentration of triptolide in patients with rheumatoid arthritis and its pharmacokinetics study[J].ChinJInfTraditChinMed,2014, 21(1):85-7.
[10]焦 娟,唐曉頗,員 晶,等.復方雷公藤外敷劑對類風濕關節炎患者關節疼痛的影響[J].中國中西醫結合雜志,2016,36(1):29-34.
[10]Jiao J, Tang X P, Yuan J, et al. Effect of external applying compound tripterygium wilfordii Hook F. on joint pain of rheumatoid arthritis[J].ChinJIntegrTraditWestMed, 36(1):29-34.
[11]Chai X, Zeng S, Xie W. Nuclear receptors PXR and CAR: implications for drug metabolism regulation, pharmacogenomics and beyond[J].ExpertOpinDrugMetabToxicol, 2013, 9(3):253-66.
[12]周 濤,王宇光,馬增春,等.銀杏內脂B通過激活孕烷X受體誘導CYP3A4的表達[J].中國藥理學通報,2014,30(7):926-31.
[12]Zhou T, Wang Y G, Ma Z C, et al.Ginkgolide B induces CYP3A4 expression through activation of human pregnane X recptor[J].ChinPharmacolBull, 2014, 30(7):926-31.
[13]Naspinski C, Gu X, Zhou G D, et al. Pregnane X receptor protects HepG2 cells from BaP-induced DNA damage[J].ToxicolSci, 2008, 104(1):67-73.
[14]Fahmi O A, Raucy J L, Ponce E, et al. Utility of DPX2 cells for predicting CYP3A induction-mediated drug-drug interactions and associated structure-activity relationships[J].DrugMetabDispos,2012,40(11):2204-11.
[15]Li W, Liu Y, Li X X, et al. MAPKs are not involved in triptolide-induced cell growth inhibition and apoptosis in prostate cancer cell lines with different p53 status[J].PlantaMedica,2011,77(1):27-31.
Investigation of inductive effect of triptolide on cytochrome P450s in rat hepatocytes and HepG2 cells and possible mechanism
YU Xi1,2,WAN Zi-jin1,2,ZHU Yan-ping1,2, XU Hai-rong1,2,LIAO Kai1,2, LIU Feng1, LI Wei1,2
(1.CollegeofMedicine,YangzhouUniversity,YangzhouJiangsu225001,China;2.JiangsuKeyLaboratoryofIntegratedTraditionalChineseandWesternMedicineforPreventionandTreatmentofSenileDiseases,YangzhouJiangsu225001,China)
Aim To investigating the induction of CYPs in hepatocytes or HepG2 cells by triptolide(TP) and the possible mechanism.Methods After TP treatment, the expression of CYPs in rat primary hepatocytes or human HepG2 cells was detected by real-time PCR and Western blot assays.Specific inhibitors or gene knockdown method were employed to analyze the possible mechanism.Results The expression of CYP1A2, 2C7, 2C11, 2C12, 2D2, 2E1 and 3A1 in rat primary hepatocytes was induced by TP. The fold was 19, 2, 31, 3, 21, 88 and 34 at 50 nmol·L-1,respectively while at 100 nmol·L-1it was 20, 5, 30, 23, 61, 83 and 38,respectively. In HepG2 cells, the expression of human CYP1A1, 2B6, 2C9, 2C19, 2D6, 2E1 and 3A4 was also induced by TP. The activities of nuclear receptor PXR and CAR were inhibited.TP upregulated p53 expression, and the induction of several CYPs caused by TP was blocked when p53 was inhibited.Conclusions TP induces CYPs expression in rat hepatocytes and HepG2 cells.Nuclear receptors may not be involved in TP induced CYPs, while the mechanism may partly attribute to p53.
triptolide(TP);cytochrome P450(CYPs);nuclear receptor;p53;hepatocytes; HepG2
時間:2017-3-4 11:50
http://kns.cnki.net/kcms/detail/34.1086.R.20170304.1150.030.html
2016-11-07,
2016-12-10
國家自然科學基金資助項目(No 81303109);揚州市省高校自然科學基金面上項目(No 14KJB310026);揚州市自然科學基金青年科技人才項目(No YZ2014020);揚州大學大學生創新計劃(No x2015764)
虞 茜(1991-),女,碩士生,研究方向:抗炎藥物,E-mail:625428660@qq.com; 李 巍(1982-),女,博士,講師,研究方向:藥物代謝,通訊作者,E-mail: weili@yzu.edu.cn
10.3969/j.issn.1001-1978.2017.03.015
A
1001-1978(2017)03-0366-07
R-332;R284.1;R322.47;R345.99;R392.11;R977.3;R977.6
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
