TIPE3干擾質粒對SW480結腸癌細胞生長的作用及相關機制探討①
2017-04-10 11:02:43葉宇涵劉忠臣齊忠權莊國洪
中國免疫學雜志 2017年3期
葉宇涵 劉忠臣 齊忠權 莊國洪 殷 平
(福建醫科大學基礎醫學院,福州350004)
·生物治療·
TIPE3干擾質粒對SW480結腸癌細胞生長的作用及相關機制探討①
葉宇涵 劉忠臣②③齊忠權④莊國洪④殷 平⑤
(福建醫科大學基礎醫學院,福州350004)
目的:通過TIPE3干擾質粒轉染SW480結腸癌細胞,驗證干擾TIPE3表達對SW480結腸癌細胞生長的影響并探討相關機制。方法:構建TIPE3-shRNA-pSIREN-RetroQ干擾質粒,通過脂質體轉染法成功將干擾質粒導入SW480細胞,通過RT-PCR、Western blot檢測重組質粒的干擾效率。應用CCK-8方法檢測SW480細胞生存率。AnnexinV-FITC/PI流式細胞法檢測細胞凋亡。使用Western blot檢測細胞增殖、凋亡相關分子的表達情況。結果:成功設計、構建和篩選具有生物活性且干擾效率最佳的TIPE3-shRNA-pSIREN-RetroQ干擾質粒。CCK-8檢測證實干擾SW480結腸癌細胞TIPE3表達可以抑制細胞生長。流式結果顯示,TIPE3-shRNA3干擾組的凋亡率為(27.99±1.087)%,顯著高于正常細胞組(12.10±2.213)%及轉染空載質粒組(11.44±0.277 0)%。證實了降低TIPE3表達可以增加SW480對aDR5ScFv所誘導的細胞凋亡敏感性。Western blot結果顯示干擾TIPE3表達可以活化caspase3蛋白,降低p-AKT、p-PDK1、PCNA等分子的表達。結論:干擾TIPE3的表達對SW480結腸癌細胞具有促進凋亡,抑制增殖的作用。
TIPE3shRNA;結腸癌;凋亡;增殖
腫瘤壞死因子-α誘導蛋白8(Tumor necrosis factor-α induced protein 8,TNFAIP8或稱TIPE)家族是近年來發現的一組蛋白質,具有維持免疫平衡、調節腫瘤生長的作用[1]。哺乳動物TNFAIP8家族包括四個成員,即TIPE、TIPE1、TIPE2與TIPE3[2],它們在結構上具有高度同源性,均包含7個高度保守的α螺旋(α0~α6)以及由α螺旋組成的疏水性空腔[3]。TIPE3(又稱TNFAIP8L3)作為磷脂酰肌醇通路中的第二信使轉移蛋白促進腫瘤的發生[4]。TIPE3在N端具有獨特的19 個氨基酸序列,稱為N 端結構域(NT),TIPE 家族的其他成員沒有該結構域,刪除這段序列 TIPE3 將從促增殖蛋白質轉換為促死亡蛋白[5]。
TIPE3在食管癌、卵巢癌、胃癌等腫瘤細胞系中表達[5,6],降低TIPE3表達對腫瘤生長的研究有助于進一步揭示TIPE3在腫瘤發生發展中的可能機制。因此我們構建TIPE3-shRNA-pSIREN-RetroQ干擾質粒,降低TIPE3在結腸癌中的表達來研究TIPE3對其生長的影響,進而了解TIPE3在腫瘤增殖、凋亡中的作用,為結腸癌治療的分子治療提供新的靶點。
1 材料與方法
1.1 材料
1.1.1 主要試劑 T4 DNA Ligase、BamHⅠ限制性內切酶、EcoR Ⅰ限制性內切酶、PrimeScriptTMRT reagent Kit with gDNA Eraser(Perfect Real Time)、DL5000 DNA marker購自寶生生物工程(大連)有限公司;PCR 產物膠回收試劑盒購于天根生化科技(北京)有限公司; ExFectTMTransfection Reagent購自 Vazyme公司;CCK購于北京全式金有限公司;流式凋亡檢測試劑盒購于BD公司;PCR 引物合成由生工生物工程(上海)股份有限公司完成;質粒測序由上海英駿公司完成;辣根過氧化酶標記的山羊抗兔IgG購廈門英偉生物技術有限公司; TIPE3抗體購自藥明康德新藥開發有限公司;caspase3、caspase3-cleaved、AKT、pAKT、PDK、pPDK抗體購于abcam公司。
1.1.2 載體和菌種 shRNA 載體pSIREN-RetroQ 由廈門大學醫學院金光輝老師饋贈,DH5α菌株購于北京全式金公司,引物及干擾片段由廈門大學醫學院高淑彬老師設計,上海 Sangon 合成,人結腸癌細胞HT-29、人結腸腺癌細胞SW480、人腎胚細胞HKE293細胞株由廈門大學醫學院抗癌研究中心保存。
1.2 方法
1.2.1 TNFAIP8干擾質粒的構建
1.2.1.1 RNA 干擾片段的設計與合成 取pSIREN-RetroQ為shRNA 載體。轉化提取質粒pSIREN-RetroQ后,用限制性內切酶BamHⅠ和EcoRⅠ對質粒 pSIREN-RetroQ進行雙酶切,雙酶切后采用TIANGEN Universal DNA 純化回收試劑盒純化回收DNA溶液。
登陸 GeneBank 查找并獲取人源性 TIPE3基因序列,從TIPE3的ORF 中選擇評分最高的靶序列位點三處,分別命名為TIPE3 shRNA1;TIPE3 shRNA2;TIPE3 shRNA3(GCGAGATCTTTGATGAGCT;ACAAGACTCATCCCCTTAT;GAGCAAAATAGCCAGCAAA)。利用SiRNA Construct Builder 軟件,構建TIPE3 干擾片段序列(含黏性末端)。將該序列送至上海Sangon 合成DNA單鏈,退火形成互補片段。酶切后的膠回收的質粒和退火片段用T4連接酶連接。
1.2.1.2 TIPE3-shRNA-pSIREN-RetroQ重組質粒的轉化、鑒定及測序 將連接產物轉化至感受態中。經培養后挑取飽滿單克隆菌落進行培育。并進行初篩、復篩以排除假陽性克隆的出現。重組質粒的初篩鑒定引物具體序列如下(設計的引物送上海Songon合成):Forward primer=GGGCAGGAAGAGGGCCTAT;TIPE3-shRNA1 reverse primer=ACGCGTAAA-AAAGCGAGA;TIPE3-shRNA2 reverse primer=ACG-CGTAAAAAAACAAGA;TIPE3-shRNA3 reverse primer=ACGCGTAAAAAAGAGCAA;復篩采用質粒手冊中的通用引物MSCV_primer和MSCV_rev_primer。
行菌液 PCR 鑒定陽性克隆,所得產物用1.5%的瓊脂糖凝膠電泳,在Bio-Rad凝膠成像儀上鑒定。將陽性菌落送上海英俊公司測序。
1.2.2 重組干擾質粒的干擾效率檢測及最佳干擾效率質粒的篩選
1.2.2.1 檢測人結腸癌細胞HT-29、人結腸腺癌細胞SW480 細胞株TIPE3表達情況 復蘇HT-29、SW480細胞,待細胞長至70%~80%后收集細胞,制備RNA樣品,并逆轉錄為cDNA,行PCR檢測TIPE3在兩株結腸癌細胞中的基因水平表達情況。β-actin上游引物5′-AGCGAGCATCCCCCAAAGTT-3′;下游引物:5′-G GGCACGAAGGCTCATCATT-3′。TIPE3上游引物:5′-CGCAGCATGGA TTCGGATT-3′;下游引物:5′-TCAAAGATCTCGCTGCTGGTGT-3′。PCR 反應條件為:(1)94℃、5 min;(2)94℃、0.5 min;(3)56℃、0.5 min;(4)72℃、0.5 min;(5)72℃、5 min;其中(2)~(4)循環35次。產物存于4℃。所得產物用1.5%的瓊脂糖凝膠電泳,在Bio-Rad凝膠成像儀上鑒定。行Western blot檢測TIPE3在兩株結腸癌細胞中蛋白水平的表達情況。PBS洗滌2次。棄上清,取細胞沉淀,加入含有蛋白酶抑制劑的蛋白裂解液裂解細胞并提取蛋白。行 SDS-PAGE電泳并轉印至PVDF膜,5%脫脂奶粉室溫封閉1 h后,滴加一抗(1∶1 000),4℃孵育過夜,滴加二抗(1∶10 000),室溫孵育1 h,ECL顯色。用已知高表達TIPE3的HEK293細胞作為對照[6]。
1.2.2.2 人結腸腺癌細胞SW480細胞轉染及檢測干擾效率 采用ExFectTMTransfection Reagent轉染試劑將TIPE3-shRNA-pSIREN-RetroQ重組質粒轉染至 SW480 細胞。選擇細胞狀態好且處于生長對數期的SW480細胞作為轉染對象。將Transfection Reagent加入分別含pSIREN-RetroQ、GFP-pSIREN-RetroQ、TIPE3-shRNA-pSIREN-RetroQ等質粒的不完全培養基中制備成轉染復合物。轉染復合物與 SW480細胞混勻后將之置于恒溫培養箱中進行轉染。48 h后利用熒光顯微鏡觀察GFP-pSIREN-RetroQ轉染效率。取各組轉染細胞制備RNA樣品,并逆轉錄為cDNA,行PCR對TIPE3的表達進行驗證和分析。行Western blot檢測轉染TNFAIP8-shRNA-pSIREN-RetroQ重組質粒的SW480細胞TIPE3干擾效率及TIPE3蛋白的表達。
1.2.3 CCK檢測轉染TIPE3-shRNA-pSIREN-RetroQ干擾質粒SW480 細胞的存活率 行CCK檢測轉染細胞生長情況:取3.0×103細胞接種到96孔板中,置于37℃、5%CO2飽和濕度的培養箱中培養24、48 h后,加入10%Cell Counting Kit-8(CCK-8)的完全培養基,4 h后,置于450nm波長測其吸光度。
1.2.4 AnnexinV-FITC/PI流式細胞檢測aDR5ScFv誘導轉染TIPE3干擾質粒SW480 細胞的凋亡情況。用流式法檢測 aDR5ScFv誘導細胞凋亡情況。在消化前4 h加入終濃度150 μg/ml 的 aDR5ScFv培養基。用不含EDTA的0.25%的胰酶消化細胞,計數1×106個細胞,用遇冷的PBS 1 000 r/min,4℃離心10 min棄上清洗滌2次后,100 μl binding buffer重
懸細胞,分別加入5 μl的PI和Annexin V-FITC 避光室溫孵育15 min,在1 h內上機完成檢測。
1.2.5 檢測轉染TIPE3干擾質粒的SW480 細胞相關分子pro-caspase3、cleaved caspase3、AKT、p-AKT、PDK、p-PDK、PCNA等表達活化 將轉染干擾質粒48 h后的SW480細胞提取蛋白,另設正常細胞組,空載細胞組,行Western blot檢測caspase3、AKT、PDK、PCNA等相關分子的表達情況。探討TIPE3干擾質粒影響細胞增殖、凋亡的可能機制。
1.3 統計學處理 全部數據使用SPSS13.0、GraphPad Prism5和Excel軟件做統計學處理。P<0.05表示差異有統計學意義。
2 結果
2.1 TIPE3-shRNA-pSIREN-RetroQ 干擾質粒陽性克隆初篩、復篩鑒定 從連接轉化后所得菌落制備PCR模板,以初篩引物進行菌液PCR初篩鑒定。取初篩為陽性的菌種作為PCR模板,以質粒pSIREN-RetroQ的通用引物MSCV_primer和MSCV_rev_primer為引物,進行復篩,將篩選為陽性的菌種送上海英俊公司測序。復篩鑒定1、2為TIPE3-shRNA1陽性克隆。鑒定3、4為TIPE3-shRNA2陽性克隆。鑒定5、6為TIPE3-shRNA3陽性克隆。復篩鑒定的PCR產物大小為410 bp(圖1)。
2.2 TIPE3-shRNA-pSIREN-RetroQ測序結果比對 測序結果經DNAMAN軟件比對,與設計靶序列一致(圖2)。
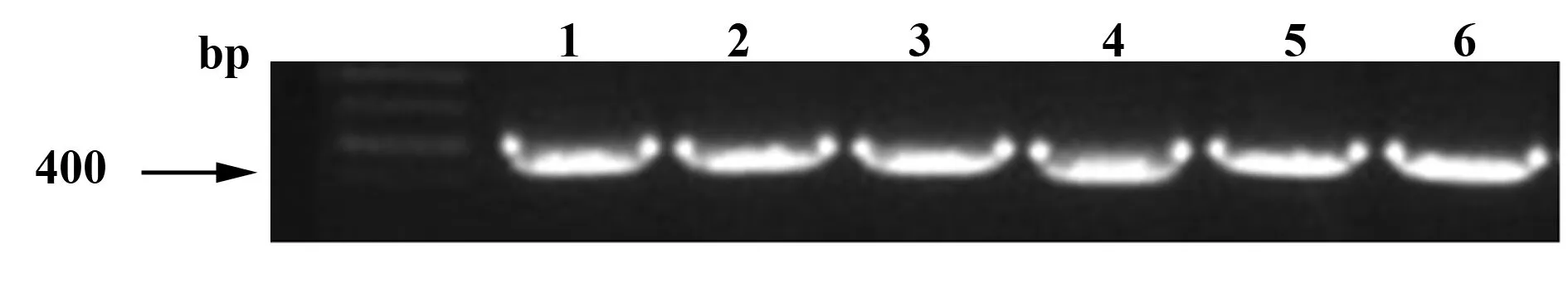
圖1 重組質粒陽性克隆復篩鑒定Fig.1 Rescreen positive recombinant plasmid clonesNote: 1,2.Positive clones of TIPE3-shRNA1;3,4.Positive clones of TIPE3-shRNA2;5,6.Positive clones of TIPE3-shRNA3.

圖2 TIPE3-shRNA-pSIREN-RetroQ重組質粒的測序圖譜和序列比對Fig.2 Sequence comparison of TIPE3-shRNA-pSIREN-RetroQ recombinant plasmid
2.3 檢測人結腸癌細胞HT-29、人結腸腺癌細胞SW480細胞株TIPE3表達情況 通過PCR及Western blot比較人腎胚細胞HEK293、人結腸癌細胞HT-29、SW480中TIPE3表達特點。文獻報道TIPE3在HEK293細胞高表達,與HEK293細胞相比,TIPE3在人結腸癌細胞HT-29、SW480也為高表達,且在表達上沒有差異。由此,我們選擇SW480作為干擾細胞株(圖3)。
2.4 人結腸腺癌細胞系SW480 細胞培養及轉染 經脂質體轉染48 h后,轉染了GFP-pSIREN-RetroQ質粒的SW480細胞,在100倍熒光顯微鏡下觀察,視野下大量綠色熒光蛋白表達。對比自然光光鏡下SW480細胞數量,可見轉染GFP-pSIREN-RetroQ質粒轉染效率達70%。相對較高的轉染效率為后面對所構建干擾質粒干擾效果的檢測提供有力基礎(圖4)。
2.5 TIPE3-shRNA-pSIREN-RetroQ質粒干擾效率檢測 收集轉染48 h后的SW480細胞,提取RNA并逆轉錄為cDNA,檢測TIPE3的基因表達情況。與正常細胞(泳道1)相比,轉染TIPE3-shRNA1、TIPE3-shRNA2、TIPE3-shRNA3質粒(泳道2~4)的SW480細胞,TIPE3的表達量均降低,且轉染TIPE3-shRNA3 質粒干擾效率最高(圖5)。轉染48 h后提取SW480細胞蛋白進行Western blot檢測。TIPE3-shRNA3干擾組中TIPE3表達量明顯降低,低于空載質粒和TIPE3-shRNA1/2干擾質粒轉染組。泳道1~5分別為正常細胞、pSIREN-RetroQ、TIPE3-shRNA1、TIPE3-shRNA2和TIPE3-shRNA3質粒轉染組。綜上測試結果,構建的TIPE3-shRNA3-pSIREN-RetroQ干擾質粒在蛋白和RNA水平均能較好地干擾TIPE3的表達(圖6)。
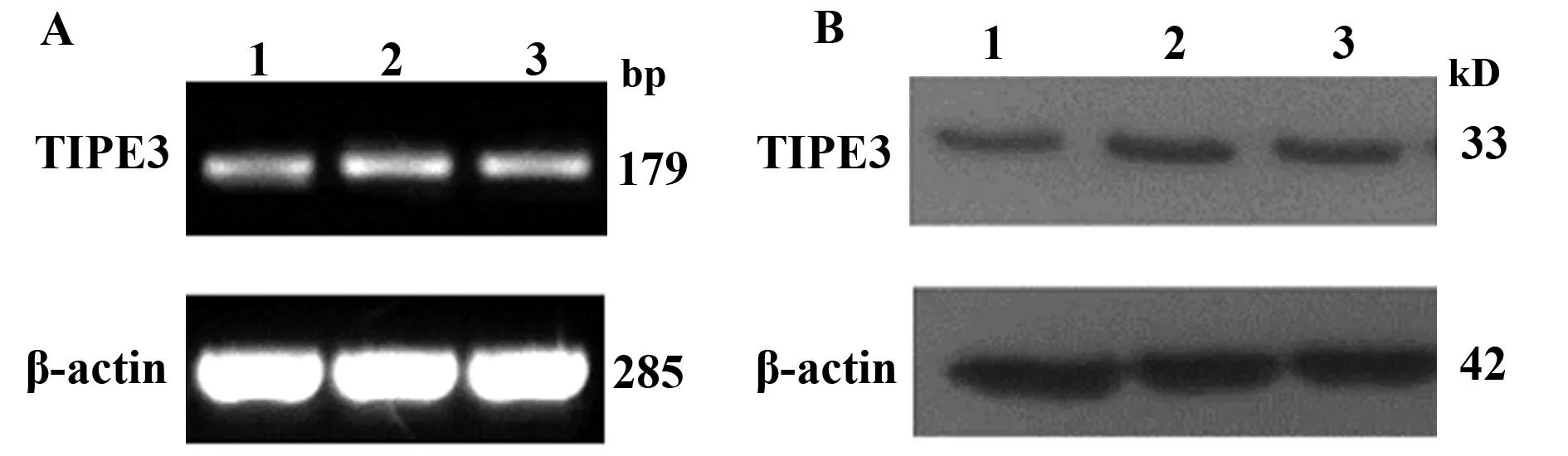
圖3 SW480、HT-29與HEK293細胞TIPE3表達情況的比較 Fig.3 Comparison of expression of TIPE3 in HEK293,SW480 and HT-29Note: A.RT-PCR;B.Western blot;1.HEK293 cell;2.SW480 cell;3.HT-29 cell.

圖4 同一視野下熒光顯微鏡觀察GFP-pSIREN-RetroQ 轉染SW480細胞Fig.4 SW480 cell observed by fluorescence after GFP-pSIREN-RetroQ transfection at same field of visionNote: A.Observation under natural light;B.Observation under blue light.
2.6 CCK-8檢測TIPE3-shRNA-pSIREN-RetroQ干擾質粒轉染后SW480細胞的生長 為了進一步驗證干擾TIPE3的表達對結腸癌細胞的影響,分別在轉染后24、48 h對結腸癌細胞行CCK-8檢測,結果表明無論是24 h還是48 h ,與正常的細胞相比,空載組的細胞存活率遠高于實驗組不同干擾序列組,表明干擾TIPE3后細胞生長能力受限。且48 h時,TIPE3-shRNA3表現出最強的抑制作用,細胞存活率與TIPE3蛋白干擾時間成負相關。說明構建的TIPE3干擾質粒具有抑制腫瘤細胞生長的作用,并呈時間依賴性(圖7)。
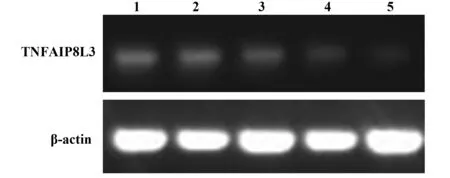
圖5 應用RT-PCR檢測構建質粒的干擾效果 Fig.5 Detection of interference of recommend plasmid by RT-PCRNote: 1.Normal SW480 cell;2.SW480 cell transfected with pSIREN-RetroQ empty control plasmid;3.SW480 cell transfected with TIPE3-shRNA1 plasmid;4.SW480 cell transfected with TIPE3-shRNA2 plasmid;5.SW480 cell transfected with TIPE3-shRNA3 plasmid.
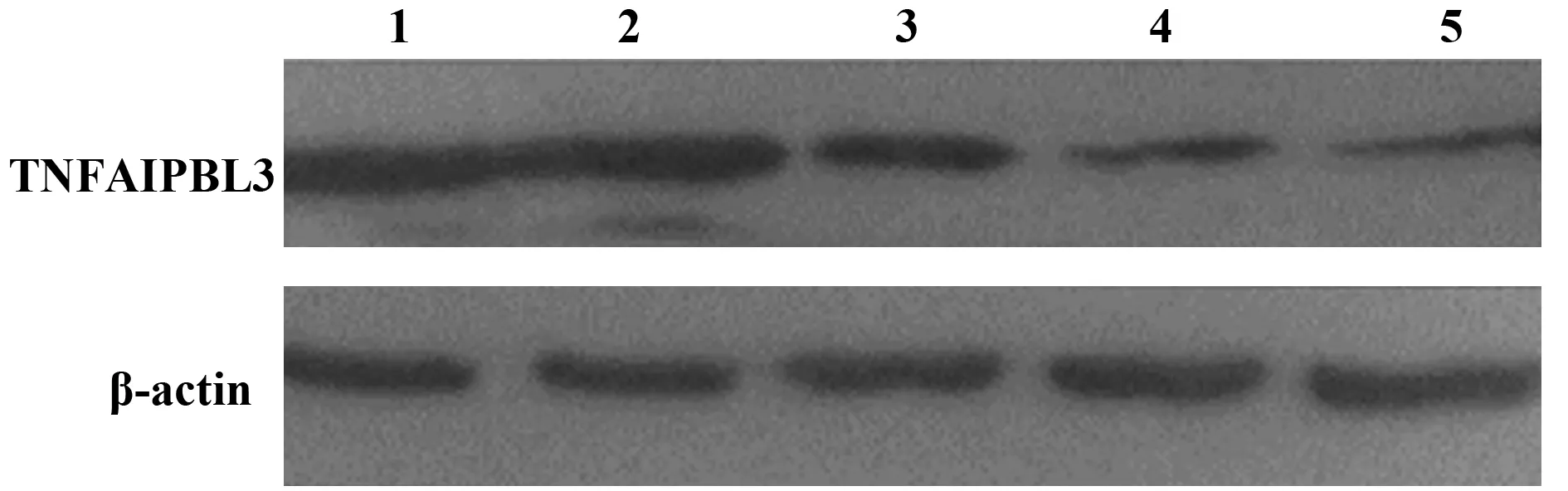
圖6 TIPE3-shRNA質粒轉染SW480細胞蛋白水平表達的檢測Fig.6 Detection of protein expression of TNFAIP8L3-shRNA plasmid transfected SW480 cellsNote: 1.Normal SW480 cell;2.SW480 cell transfected with pSIREN-RetroQ empty control plasmid;3.SW480 cell transfected with TIPE3-shRNA1 plasmid;4.SW480 cell transfected with TIPE3-shRNA2 plasmid;5.SW480 cell transfected with TIPE3-shRNA3 plasmid.
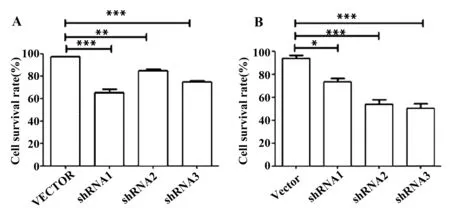
圖7 CCK-8檢測shRNA干擾TIPE3表達對SW480結腸癌細胞存活率的影響Fig.7 CCK-8 detect shRNA impact of SW480 cell survival rateNote: A.24 h;B.48 h,*.P< 0.05;**.P<0.01;***.P< 0.000 1.
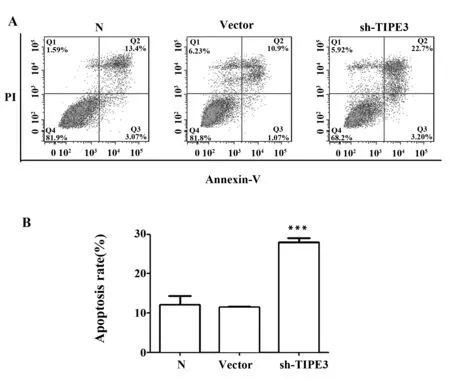
圖8 流式檢測aDR5ScFv誘導SW480細胞的凋亡Fig.8 Flow cytometry detect aDR5ScFv induced SW480 cells apoptosisNote: ***.P< 0.000 1.
2.7 流式檢測 TIPE3-shRNA3-pSIREN-RetroQ 干擾質粒對細胞凋亡的影響 本實驗室前期實驗結果已證實aDR5ScFv(抗人死亡受體5單鏈抗體)可以通過結合DR5受體誘導腫瘤細胞的凋亡[7,8]。流式細胞術檢測結果表明終濃度 150 μg/ml的 aDR5ScFv 作用6 h后,正常SW480細胞凋亡率為(12.10±2.213)%,轉染空載質粒的SW480細胞凋亡率為(11.44±0.277)%,TIPE3-shRNA3干擾組的凋亡率為(27.99±1.087)%。與前兩者相比干擾組細胞凋亡增加明顯,差異具有統計學意義(P<0.05)。而正常SW480細胞組與轉染空載質粒的SW480細胞組相比,差異無統計學意義(P>0.05)。比較正常組、空載組、干擾組細胞早期凋亡(Q3象限)發現,三者差異無統計學意義(圖中未顯示)。說明構建的TIPE3干擾質粒能夠提高結腸癌細胞SW480對aDR5ScFv的敏感性,主要通過增加中晚期凋亡促進腫瘤細胞凋亡的作用(圖8)。
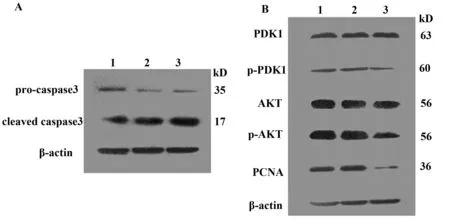
圖9 Western blot檢測caspase3、PDK1、AKT、PCNA等蛋白的表達Fig.9 Detection of expression of caspase3,PDK1,AKT and PCNA by Western blotNote: 1.Normal SW480 cell;2.SW480 cell transfected with empty control plasmid;3.SW480 cell transfected respectively with TIPE3-shRNA3 plasmid.
2.8 Western blot檢測細胞凋亡、增殖相關蛋白分子的表達情況 應用Western blot檢測caspase家族成員:pro-caspase3與cleaved caspase3的表達情況。結果可見,與正常細胞組及空載細胞組比較,SW480細胞干擾組其cleaved caspase3明顯升高。caspase3作為細胞凋亡的執行者,活化的cleaved caspase3表達增高表明降低TIPE3表達可以促進了細胞凋亡(圖9A)。并且在Total PDK1、AKT表達無明顯差異的情況下,干擾質粒組p-PDK1、p-AKT蛋白質的表達明顯下調,而正常細胞組與空載質粒組表達量沒有明顯差異。結果提示干擾TIPE3表達可能降低PDK1、AKT磷酸化,進而抑制磷脂酰肌醇信號通路,使結腸癌細胞增殖減少。同時細胞增殖狀態的PCNA蛋白減少明顯,進一步證實了結腸癌細胞生長受到抑制(圖9B)。
3 討論
超過50%的人類腫瘤細胞有異常的磷脂酰肌醇信號活化,雖然一大部分原因可以由PTEN、Ras或者PI3KCA的突變做出解釋[9,10],但肯定存在其他的機制上調了該信號的活化。TIPE3作為磷脂酰肌醇轉運蛋白,可以使腫瘤細胞中的磷脂酰肌醇信號的異常活躍,能夠直接調節Ptdlns(3,4,5)P3、Ptdlns(4,5)P2的含量,闡述了另外一種腫瘤發生、發展的機制,可為腫瘤治療提出了新的靶點。但現在對TIPE3的研究成果很少,PubMed中針對TIPE3的試驗研究有且只有2篇。TIPE3對腫瘤的增殖、遷移、侵襲的機制有待進一步探討,為了更好地研究其對腫瘤細胞的影響,我們構建了TIPE3-shRNA-pSIREN-RetroQ干擾質粒。
shRNA是常被用于沉默靶基因的短發夾RNA序列[11],有與內源性mRNA一致的兩個短反向重復序列,是RNA干擾的靶點。我們成功構建了質粒TIPE3-shRNA1-pSIREN-RetroQ、TIPE3-shRNA2-pSIREN-RetroQ和TIPE3-shRNA3-pSIREN-RetroQ。由于載體不具有綠色熒光標記,我們用具有GFP標簽的GFP-pSIREN-RetroQ質粒作為判斷轉染效率的標準。我們通過RT-PCR、Western blot篩選發現,TIPE3-shRNA2和TIPE3-shRNA3都可以干擾TIPE3的表達,而TIPE3-shRNA3質粒具有最佳的干擾效率,可以顯著降低TIPE3在細胞中的表達。所以,我們將應用TIPE3-shRNA3質粒干擾TIPE3在SW480中的表達來探討TIPE3對結腸癌細胞生長的影響。
我們通過CCK-8檢測發現,在24 h與48 h的時間點,與正常細胞相比,空載組的細胞存活率達到90%;轉染了不同干擾序列的細胞組與正常細胞組及空載組相比差異顯著,差異具有統計學意義(P<0.05)。經RT-PCR、Western blot驗證轉染48 h的細胞存活率與24 h相比差異明顯。以上結果表明干擾TIPE3表達具有抑制腫瘤細胞生存的作用。文獻報道TIPE3促進腫瘤細胞的生長[5]。我們的實驗結果與文獻報道TIPE3在腫瘤生長方面起正向調控作用相符。干擾該基因表達,腫瘤生存受到抑制。
腫瘤的生長、存活與腫瘤凋亡、腫瘤增殖密不可分。為了驗證干擾TIPE3表達,降低腫瘤生存率是通過促進腫瘤凋亡還是抑制腫瘤增殖發揮作用,我們選擇具有干擾效果最強的TIPE3-shRNA3質粒轉染的細胞,并進行流式細胞術檢測。結果發現終濃度150 μg/ml 的aDR5ScFv作用6 h,干擾組細胞的凋亡率增加明顯,與正常細胞和空載組細胞相比差異顯著。Western blot結果顯示,干擾組中caspase3活化增強。caspase家族參與的細胞凋亡信號轉導,其中caspase3是主要的凋亡執行者[12]。干擾TIPE3在SW480細胞中的表達可以增強SW480細胞對aDR5ScFv誘導的凋亡敏感性,并且,干擾TIPE3表達可以通過激活caspase通路增加SW480細胞的凋亡。
眾所周知,PDK1/AKT通路作為磷脂酰肌醇信號轉導中的重要節點,調節細胞生長、增殖、分化、遷移等[13,14]。Western blot結果證實干擾TIPE3表達可以抑制AKT、PDK磷酸化、降低磷脂酰肌醇信號轉導,減少腫瘤細胞增殖。PCNA作為細胞增殖蛋白,在結腸癌分期、預后判斷具有重要意義[15],PCNA蛋白表達下調,表明細胞增殖受到抑制,我們的結果進一步證明干擾TIPE3表達抑制SW480細胞增殖。
綜上所述,TIPE3對調控腫瘤生長具有重要作用,主要通過影響腫瘤細胞增殖、凋亡發揮效果。干擾其表達為以后臨床分子靶向治療提供新的思路。
[1] Freundt EC,Bidere N,Lenardo MJ.A different TIPE of immune homeostasis[J].Cell,2008,133(3):401-402.
[2] Sun H,Gong S,Carmody RJ,etal.TIPE2,a negative regulator of innate and adaptive immunity that maintains immune homeostasis[J].Cell,2008,133(3):415-426.
[3] Zhang X,Wang J,Fan C,etal.Crystal structure of TIPE2 provides insights into immune homeostasis[J].Nat Struct Mol Biol,2009,16(1):89-90.
[4] Fayngerts SA,Wu J,Oxley CL,etal.TIPE3 is the transfer protein of lipid second messengers that promote cancer[J].Cancer Cell,2014,26(4):465-478.
[5] Moniz LS,Vanhaesebroeck B.A new TIPE of phosphoinositide regulator in cancer[J].Cancer Cell,2014,26(4):443-444.
[6] Cui J,Hao C,Zhang W,etal.Identical expression profiling of human and murine TIPE3 protein reveals links to its functions[J].J Histochem Cytochem,2015,63(3):206-216.
[7] 孟慶宇,程小峰,張佳鍇,等.抗人DR5單鏈抗體誘導HepG2細胞凋亡的實驗研究[J].免疫學雜志,2010,26(7):585-593.
[8] 孟慶宇,程曉峰,張佳鍇,等.抗人死亡受體5單鏈抗體ZF1對鼠H22肝癌細胞的作用分析[J].中國免疫學雜志,2010,26(9):801-809.
[9] Yuan TL,Cantley LC.PI3K pathway alterations in cancer:variations on a theme[J].Oncogene,2008,27(41):5497-5510.
[10] Vivanco I,Sawyers CL.The phosphatidylinositol 3-kinase AKT pathway in human cancer[J].Nat Rev Cancer,2002,2(7):489-501.
[11] Paddison PJ,Caudy AA,Bernstein E,etal.Short hairpin RNAs(shRNAs)induce sequence-specific silencing in mammalian cells[J].Genes Dev,2002,16(8):948-958.
[12] Porter AG,Janicke RU.Emerging roles of caspase-3 in apoptosis[J].Cell Death Differ,1999,6(2):99-104.
[13] Itoh N,Semba S,Ito M,etal.Phosphorylation of Akt/PKB is required for suppression of cancer cell apoptosis and tumor progression in human colorectal carcinoma[J].Cancer,2002,94(12):3127-3134.
[14] Lee SH,Kim HS,Park WS,etal.Non-small cell lung cancers frequently express phosphorylated Akt;an immunohistochemical study[J].APMIS,2002,110(7-8):587-592.
[15] al-Sheneber IF,Shibata HR,Sampalis J,etal.Prognostic significance of proliferating cell nuclear antigen expression in colorectal cancer[J].Cancer,1993,71(6):1954-1959.
[收稿2016-10-19 修回2016-12-27]
(編輯 張曉舟)
Effect and mechanism of TIPE3 interference plasmid on SW480 colorectal cancer growth
YEYu-Han,LIUZhong-Chen,QIZhong-Quan,ZHUANGGuo-Hong,YINPing.
TheSchoolofBasicMedicalSciences,FujianMedicalUniversity,Fuzhou350004,China
Objective:To study the effect of interference TIPE3 on the colon cancer cell growth by transfecting SW480 colon cancer cells with the TIPE3 interference plasmid were detected.Methods: Transfecting the constructed TIPE3-shRNA-pSIREN-RetroQ plasmid to SW480 cells.To determine the highest interference efficiency plasmid,the mRNA and protein levels of recombined plasmid were detected by RT-PCR and Western blot separately and tested the cell proliferation with CCK8.Meanwhile,apoptosis rate of SW480 cells was determined by flow cytometry assay with AnnexinV-FITC/PI.To further determined the effects of recombined plasmid on cell development,the level of protein involved in proliferation and apoptosis were detected by Western blot.Results: The most effecient interference plasmids were successfully constructed.We found that the cell survival rate decreased when interference TIPE3 gene expressing in colorectal cancer cells.Flow cytometry indicated that interefering the expression of TIPE3 would increase the sensitivity of SW480 cell to apoptosis induced by aDR5ScFv.The results of Western blot showed that low expression of TIPE3 would activate caspase3 and downregulate the expression of p-AKT,p-PDK1 and PCNA.Conclusion: Interference TIPE3 could promote apoptosis and inhibit proliferation in SW480 colon cancer cells.
TIPE3 shRNA;Colon cancer;Apoptosis;Proliferation
10.3969/j.issn.1000-484X.2017.03.013
①本文為國家自然科學基金(81272720)和福建省屬公益類科研院所基本科研專項項目(2016R1102)資助。
葉宇涵(1991年-),女,在讀碩士,主要從事結腸癌分子病理學研究,E-mail:yeyuhan2688@hotmail.com。
及指導教師:莊國洪(1969年-),女,博士,副教授,主要從事腫瘤免疫研究,E-mail:zghxmu@163.com。 殷 平(1963年-),男,博士,教授,主要從事炎癥及腫瘤分子病理學研究,E-mail:yinping2002@163.com。
R735.3
A
1000-484X(2017)03-0378-06
②共同第一作者。
③同濟大學第十人民醫院普通外科,上海200072。
④廈門大學抗癌研究中心,器官移植研究所,廈門361004
⑤廈門大學附屬中山醫院病理科,廈門361004。
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
文苑(2018年22期)2018-11-19 02:54:14
西南國防醫藥(2016年7期)2016-12-01 06:01:15
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
海峽科技與產業(2016年3期)2016-05-17 04:32:12
中國衛生標準管理(2015年1期)2016-01-14 03:41:26
