一株產菌核曲霉的分離鑒定及生物學特性研究
2017-04-14 07:37:52寧露娟韓建榮敖新宇
山西農業科學 2017年3期
寧露娟,韓建榮,敖新宇
(1.西南林業大學生命科學學院,云南昆明650224;2.山西大學生命科學學院,山西太原030006)
一株產菌核曲霉的分離鑒定及生物學特性研究
寧露娟1,韓建榮2,敖新宇1
(1.西南林業大學生命科學學院,云南昆明650224;2.山西大學生命科學學院,山西太原030006)
從20種森林土樣中共分離出78株曲霉,其中,從四川省南充市西充縣華光鄉柏樹根部土壤中分離得到產菌核曲霉N1菌株;通過對菌落形態、個體形態特征觀察及ITSrDNA序列分析,將N1菌株鑒定為黃曲霉原變種(Aspergillus flavus var.flavus)。生物學特性研究結果表明,CYA和CA為N1菌株產菌核的最適培養基,N1菌株的最適產菌核溫度為30℃,最適產菌核pH值是6;培養基的碳氮源對菌核生物量有一定影響,最優碳源為蔗糖,最優氮源為蛋白胨,培養基中含碳量在9.88~39.53 g/L、含氮量在0.42~0.84 g/L時,菌核生物量最大;無機鹽對N1菌株菌核生物量也具有明顯的影響作用。
曲霉;菌核;ITS;生物學特性
曲霉(Aspergillus)屬于半知菌亞門(Deuteromycotina)絲孢綱(Hyphomycetes)絲孢目(Moniliates)叢梗孢科(Moniliaceae),其廣泛分布在谷物、空氣、土壤和各種有機物品上,是引起多種物質霉腐的主要微生物之一,主要作用是釀酒、制醋等。曲霉在幼小而活力旺盛時,菌絲體產生大量的分生孢子梗,分生孢子梗頂端膨大成為頂囊,一般呈球形。頂囊表面長滿1層或2層輻射狀小梗(初生小梗與次生小梗),其中,最上層小梗瓶狀,頂端著生成串的球形分生孢子[1],孢子呈綠、黃、橙、褐、黑等顏色。這些都是菌種鑒定的依據[2]。曲霉屬的某些種能產生菌核結構。
菌核[3]是真菌生長到一定階段,菌絲體不斷地分化,相互糾結在一起形成的顏色較深而堅硬的菌絲體組織顆粒,同時它又是糖類和脂類等營養物質的儲藏體。菌核形態多樣,色澤和大小差異也很大,大菌核可達15 kg,而小菌核只有小米粒大小或者更小。
由于能形成菌核的絲狀真菌中有很多是很重要的植物病原體,因而其在農業科學方面引起了十分廣泛的研究。同時,在生物科學方面,絲狀真菌菌核的形成也引起了眾多學者的興趣,這是因為它們被看作是菌核形態變異簡單形式的代表,這使得它們成為研究菌核形態變異這個生物現象的重要模型,而菌核形態變異的機制也為代替傳統有毒殺菌劑提供了新的可能。通常,這些真菌在不利的環境條件下可以存活很長時間,具有較強的耐化學性和抗生物降解性[4-6]。產菌核真菌為了抵御來自自然環境中各種各樣導致氧脅迫的不良因素,就會形成菌核,并進化出一套有效的抗氧化系統來維持細胞內氧化還原的動態平衡[7]。因而,一般情況下,菌核具有較高的抗氧化活性,可以為利用產菌核曲霉開發相關天然抗氧化劑產品提供一定理論依據。
本試驗對20種不同森林土壤樣品中的產菌核曲霉進行了選擇性分離,并對分離到的菌株運用形態特征觀察、核糖體內轉錄間隔區(ITS)序列分析進行分類鑒定和生物學特性研究,旨在發現更多的產菌核曲霉,并初步確定其產菌核的適宜條件。
1 材料和方法
1.1 土樣采集
土壤樣品分別采自四川省南充市、云南省昆明市、山西省忻州市和太原市、貴州省都勻市和安順市、黑龍江省佳木斯市等20個地區針葉林根部表面10~20 cm處土壤。
1.2 分離和形態觀察培養基[8-9]
曲霉分離培養基為馬丁氏培養基。
采用查氏酵母膏瓊脂培養基(CYA)、麥芽汁瓊脂培養基(MEA)、查氏瓊脂培養基(CA)、馬鈴薯葡萄糖瓊脂培養基(PDA)、25%甘油硝酸鹽瓊脂培養基(G25N)作為形態觀察培養基。
1.3 產菌核曲霉的分離純化及形態觀察
將土壤樣品梯度稀釋,涂布法均勻涂布于馬丁氏平板上,每梯度3個平板,倒置于25℃的恒溫培養箱中培養3~5 d后挑取曲霉三點式接種于CA平板上,14 d后篩選出產菌核曲霉菌株,經多次分離純化得到純菌株,編號并保存于4℃冰箱。將產菌核曲霉三點式分別接種到CYA,MEA,CA,PDA和G25N平板上,于25℃培養箱內倒置培養14 d,按照文獻[1,10]的方法,觀察產菌核曲霉的菌落形態特征,顯微鏡檢其分生孢子頭、頂囊及菌核等個體形態特征。
1.4 產菌核曲霉的ITS序列分析與分類鑒定
配制CA培養基,待培養基凝固后向培養基表面平整地鋪入玻璃紙,在玻璃紙上接種產菌核曲霉,于25℃培養箱內倒置培養3~5 d,收集玻璃紙上的菌絲體,烘干備用。使用Ezup柱式真菌基因組DNA抽提試劑盒(上海生工生物工程股份有限公司)提取產菌核曲霉的DNA,通用引物ITS1和ITS4作為rDNA ITS區段PCR擴增的引物,PCR反應體系及PCR循環條件列于表1,2。擴增完成后,取0.5 μL PCR產物,與1.5 μL含有Mg2+的Loading Buffer混合,Marker 5 μL,1%瓊脂糖電泳,150 V,100 mA,20 min后用UV凝膠成像系統電泳觀察并保存圖片。PCR產物送至生工生物工程(上海)股份有限公司進行測序,將獲得的結果在GenBank數據庫進行BLAST比對和同源性分析。

表1 PCR反應體系

表2 PCR循環條件
1.5 產菌核曲霉的生物學特性研究[11-13]
1.5.1 最適培養基測定 配制CYA,MEA,CA,PDA和G25N培養基,將產核曲霉的單個菌核三點式接種法接種到上述5種平板上,于25℃培養箱內倒置培養14 d,觀察菌落生長情況,測量菌落大小,測定菌核干質量,確定產菌核的最適培養基。
1.5.2 最適產菌核溫度測定[8]將產菌核曲霉單個菌核三點式接種到CYA平板上,培養溫度分別設定為15,20,25,30,35℃,培養14 d后收集菌核并測定菌核干質量,確定產菌核的最適溫度。
1.5.3 最適產菌核pH值測定 用1 mol/L NaOH和1 mol/L HCl調節CYA的pH值,使其pH值分別為4,5,6,7,8,9,10。將產菌核曲霉的單個菌核三點式接種到配制好的CYA平板上,倒置于30℃培養箱內培養14 d,觀察記錄菌落的生長狀況并收集測定菌核干質量,確定產菌核的最適pH值。
1.5.4 最適碳源、氮源測定 分別用葡萄糖、乳糖、麥芽糖、可溶性淀粉和果糖代替CA中的蔗糖;分別用硝酸銨、硫酸銨、氯化銨、蛋白胨和酵母膏代替CA中的硝酸鈉。將產菌核曲霉的單個菌核三點式接種到配制好的平板上,倒置于30℃培養箱內培養14 d并測定菌核干質量,確定最適宜產菌核的碳、氮源。
1.5.5 最適碳源、氮源濃度測定 在CA中分別加入蔗糖11.735,23.47,46.94,93.88,187.76 g/L,使得CA中含碳量分別為4.94,9.88,19.76,39.53,79.06 g/L,C/N分別為10∶1,20∶1,40∶1,80∶1,160∶1。在CA中分別加入硝酸鈉10.23,5.11,2.56,1.70,1.02 g/L,使得CA中含氮量分別為1.69,0.84,0.42,0.28,0.17 g/L,C/N分別為7.5∶1,15∶1,30∶1,45∶1,75∶1。將產菌核曲霉的單個菌核三點式接種到配制好的平板上,倒置于30℃培養箱內培養14 d并測定菌核干質量,確定產菌核的最適碳源、氮源濃度和C/N。
1.5.6 無機鹽利用試驗 將CA中的磷酸氫二鉀、硫酸鎂、氯化鉀和硫酸亞鐵按照不同組合進行添加,將產菌核曲霉的單個菌核三點式接種到配制好的平板上,倒置于30℃培養箱內培養14 d并測定菌核干質量,觀察不同無機鹽組合形式對產菌核曲霉生長情況及菌核生物量的影響作用。
1.6 菌核干質量測定
用自來水沖洗平板,將曲霉菌株的孢子沖去并刮洗所形成的菌核,將菌核反復沖洗干凈后放置于60℃烘箱烘干,記錄菌核干質量。
1.7 數據分析
以上試驗均設3次重復,數據用SPSS 17.0進行處理;取3次數據的平均值,結果以平均值±標準差表示;用Duncan多重比較法[14]進行多個均數間兩兩比較的顯著性檢驗。
2 結果與分析
2.1 產菌核曲霉的分離結果
從20種土壤樣品中共分離得到78株曲霉菌株,其中,產菌核霉菌一株,命名為N1。該菌株分離自四川省南充市西充縣華光鄉的柏樹根部土壤樣品,采集地點東經106°1′4.27″、北緯37°48′12.78″,海拔395 m。
2.2 N1菌株的菌落形態和個體形態觀察結果
N1菌株在CYA培養基上于25℃培養6 d后,菌落直徑為58~63 mm,菌落中央灰綠色呈絮狀突起,菌落表面聚集大量墨綠色分生孢子,菌落邊緣為白色菌絲,孢子底部間隙處有黑色菌核產生,菌核表面有滲出液,菌落邊緣可見大量白色未成熟菌核(圖1-A);培養基背面可見菌落腐蝕培養基形成放射狀溝紋,菌落中心形成菌核處顯現黑褐色斑點(圖1-A′)。培養第14天,菌落直徑59~69 mm,菌落表面均勻分布完全成熟的黑色菌核,滲出液消失(圖2-A);培養基背面均勻分布大量黑褐色斑點,有放射狀溝紋(圖2-A′)。
Nl菌株在MEA培養基上于25℃培養6 d后,菌落直徑為52~58 mm,深綠色絮狀突起,菌落中央可見少量黑色菌核,無滲出液,菌落邊緣為白色菌絲夾雜白色未成熟菌核(圖1-B);菌落背面平坦,顯現大量白色斑點,菌落邊緣分布密集(圖1-B′)。培養第14天,菌落直徑為53~64 mm,菌落表面均勻分布完全成熟的黑色菌核及未完全成熟的深褐色菌核,未成熟菌核有滲出液(圖2-B);培養基背面平坦,較第6天時顏色變深呈淺褐色,白色斑點變少(圖2-B′)。
N1菌株在PDA培養基上于25℃培養6 d后,菌落直徑為43~48 mm,菌落中央為環形白色絮狀突起,菌落表面聚集大量深綠色分生孢子,菌落中心可見少量深褐色未成熟菌核,有滲出液,菌落邊緣為白色菌絲夾雜白色未成熟菌核(圖1-C);培養基背面平坦,菌落邊緣分布白色斑點(圖1-C′)。培養第14天,菌落直徑為52~60 mm,菌落邊緣及菌落中心絮狀環帶處分布少量完全成熟的黑色菌核及未完全成熟的深褐色菌核,未成熟菌核有滲出液(圖2-C);培養基背面平坦,較第6天時顏色略深呈淺黃褐色(圖2-C′)。
N1菌株在CA培養基上于25℃培養6 d后,菌落直徑為37~42 mm,菌落表面中心聚集黃綠色分生孢子,菌落表面形成大量黑褐色菌核,伴隨有淺褐色滲出液,菌落中心滲出液形成小液珠,菌落邊緣可見白色菌絲夾雜少量未成熟菌核(圖1-D);培養基背面平坦,顏色為淺黃褐色(圖1-D′)。培養第14天,菌落直徑為39~48 mm,菌落中心聚集大量黃綠色分生孢子,菌落邊緣可見白色菌絲,滲出液消失(圖2-D);菌落背面平坦,顏色較第6天時加深呈淡黃褐色(圖2-D′)。
N1菌株在G25N培養基上于25℃培養6 d,生長緩慢,菌落直徑為11~15 mm,呈長毛狀突起,菌落中心可見少量淡黃色分生孢子,未見菌核產生(圖1-E);培養基背面可見菌落中心形成環狀溝紋(圖1-E′)。培養第14天,菌落直徑為24~33 mm,深黃綠色菌絲呈絮狀突起,菌落邊緣為黃色菌絲,未產生菌核(圖2-E);菌落背面可見放射狀溝紋(圖2-E′)。
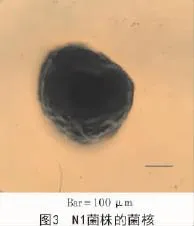
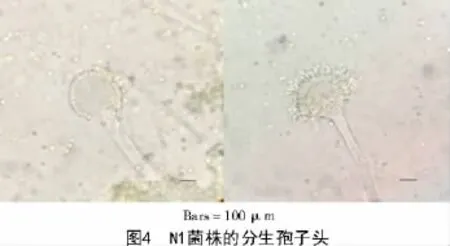
N1菌株在CYA,MEA,PDA及CA培養基上生成的菌核多為近球形(圖3),直徑為476~520 μm。N1菌株的分生孢子頭呈輻射形,291~342 μm;分生孢子梗直徑10.8~13.3 μm,壁粗糙,具小刺;頂囊球形或燒瓶形,直徑29.7~33.5 μm,產孢結構單層或雙層;分生孢子為球形或近球形,3.5~3.8 μm(圖4)。


2.3 N1菌株的ITS序列分析與分類鑒定
N1菌株的rDNA ITS區段PCR擴增得到1條單一的目的條帶(圖5),N1菌株的ITS序列長度大于500 bp。
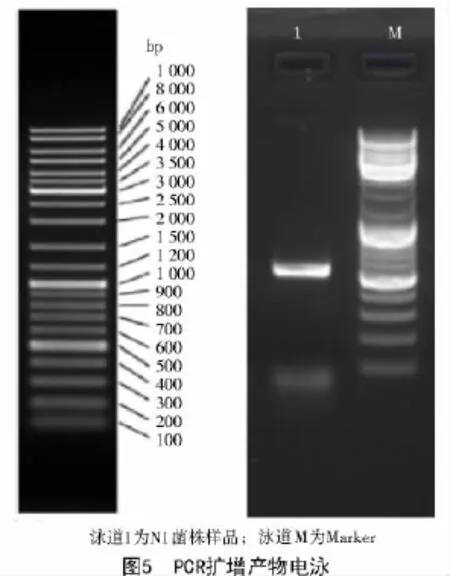
N1菌株rDNA ITS區段測序后,經編輯,長度為568 bp,在GenBank數據庫進行比對顯示,N1菌株ITS序列與黃曲霉原變種和米曲霉原變種的同源性都為100%。結合N1菌株的形態學特征,根據中國真菌志[8]中的描述,并將N1菌株鑒定為黃曲霉原變種(Aspergillus flavus var.flavus)。登錄號:KX572367。
2.4 N1菌株的生物學特性確定
2.4.1 最適培養基 N1菌株在CYA,MEA,PDA上生長迅速,培養6 d時,菌落直徑都在60 mm左右,在CA上菌核出現及成熟的時間最短,而在G25N培養基上生長最緩慢并且沒有菌核產生。從表3可以看出,在CYA和CA上形成的菌核量較大,所以本試驗條件中,將CYA和CA作為N1菌株的最適培養基及最適產菌核培養基。
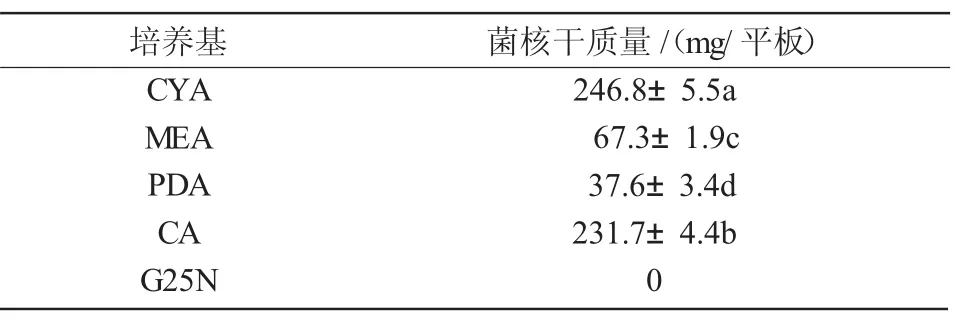
表3 N1菌株在CYA,MEA,PDA,CA,G25N培養14 d的菌核生物量
2.4.2 最適產菌核溫度 N1菌株在15~35℃溫度范圍內均能生長,在25~35℃生長較快,30℃時生長最快。從表4可以看出,20~35℃范圍內,N1菌株均能形成菌核,30℃時形成菌核的時間最短,3 d即有成熟菌核產生,且菌核生物量最高;15~20℃菌株生長緩慢,20℃時9~10 d出現菌核;15℃時未形成菌核。可見,溫度過低不利于菌落生長及菌核的形成,N1菌株最適產菌核溫度為30℃。
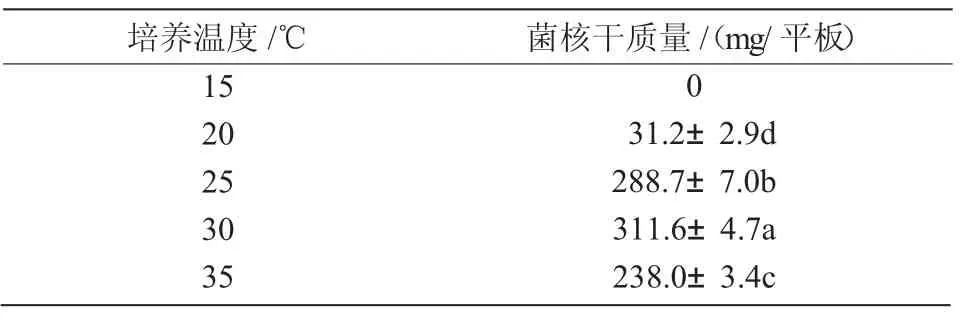
表4 不同培養溫度對N1菌株菌核生物量的影響
2.4.3 最適pH值 N1菌株在pH值為4~10的范圍內均能生長且產生菌核,其中,pH值為4時培養基呈液體,有菌核產生;pH值為5時培養基呈半固體,菌落生長迅速,產生大量菌核;pH值為6~8時菌株生長最快,菌核成熟時間最短;pH為9和10時,菌落表面分生孢子很少,菌核成熟速度較快。pH值為6~7時,菌核生物量最大(表5),說明中偏酸性的pH條件有利于N1菌株生長發育且形成大量菌核。
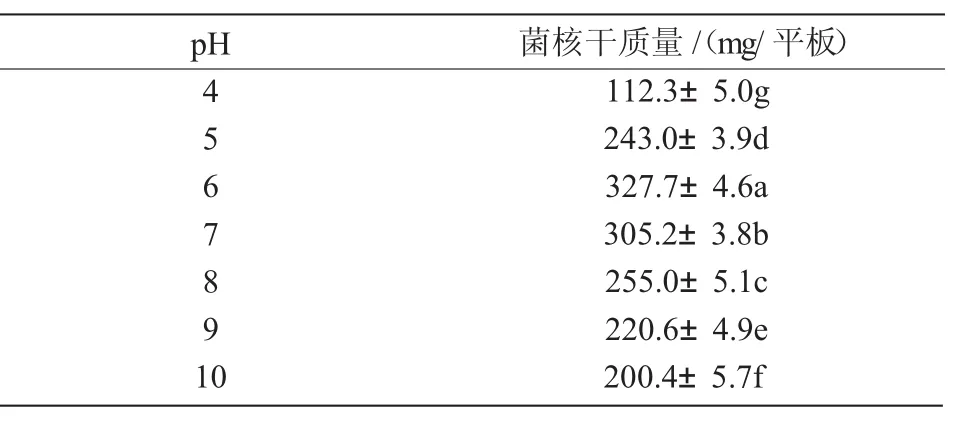
表5 不同pH值對N1菌株菌核生物量的影響
2.4.4 碳氮源利用 由表6可知,CA中不同的碳源、氮源對菌核生物量有一定影響,在試供的6種碳源中,使N1菌株菌核生物量最高的是蔗糖,最低的是乳糖;在6種氮源中,使N1菌株菌核生物量最高的是蛋白胨,當硫酸銨或氯化銨作為培養基的唯一氮源時,菌落生長緩慢,且無菌核產生。所以在本試驗條件下,葡萄糖為N1菌株最適碳源,蛋白胨為N1菌株最適氮源。

表6 不同碳源、氮源對N1菌株菌核生物量的影響
從表7可以看出,當CA中氮源濃度保持不變、改變碳源濃度時,菌核生物量有所變化,不同碳源濃度均能使N1菌株形成菌核,在含碳量為9.88~39.53 g/L,C/N為(20∶1)~(80∶1)范圍內,菌核生物量較高;同樣,在碳源濃度保持不變、改變氮源濃度的情況下,N1菌株的菌核生物量也各不相同,當含氮量為0.42~0.84 g/L,C/N為(30∶1)~(15∶1)時,N1菌株可以形成大量菌核。綜上可知,含碳量及含氮量過高或過低時,都不利于菌核形成;培養基中含碳量在9.88~39.53 g/L、含氮量在0.42~0.84 g/L時,N1菌株可以形成大量菌核。2.4.5 無機鹽利用 由表8可知,無機鹽對N1菌株菌核生物量具有明顯的影響作用,除CA原配方外,在不同的添加組合中,K2HPO4+MgSO4,K2HPO4+MgSO4+KCl,K2HPO4+MgSO4+FeSO4菌核生物量較大,說明培養基中必須同時有K2HPO4和MgSO4才能產生大量菌核,表明K2HPO4和MgSO4是N1菌株產菌核所必需的無機鹽,在沒有MgSO4的條件下,FeSO4與K2HPO4同時存在亦能產生少量菌核,而其余添加組合均未形成菌核。
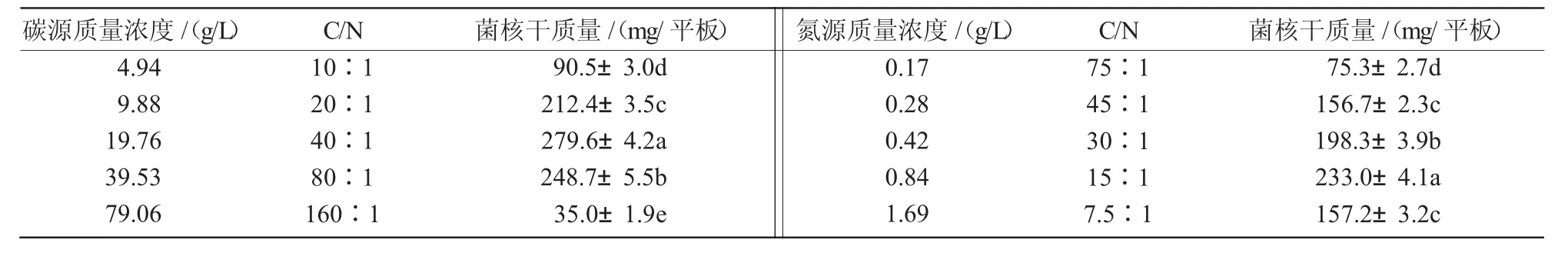
表7 不同碳源濃度和氮源濃度對菌核生物量的影響

表8 無機鹽對N1菌株菌核生物量的影響
3 討論
N1菌株在CYA,PDA,CA培養基上生長時,未成熟菌核表面有滲出液,當菌核完全成熟后滲出液消失。霉菌菌株滲出液中含有無機鹽離子、蛋白質、葡萄糖及一些酶類[15],由此猜想,N1菌株滲出液的某些物質是否與菌核發育有關,尚待深入研究。
韓建榮等[16-18]分離得到能產生橙紅色菌核的湯姆青霉PT95菌株和能產生橘紅色菌核的湯姆青霉Q1菌株,其產生的菌核內可以積累類胡蘿卜素。N1菌株所產生的黑色菌核是否也積累某種生物活性物質,還需要對N1菌株菌核中的色素進行分離提取和鑒定。
菌核具有較強的抵抗不良環境的能力,菌核的形成有助于該真菌在沒有能力降低由不良環境條件導致氧脅迫的情況下度過不良的生長環境。CHET等[19]做了諸多環境因素對菌核萌發影響的實驗,結果證明,菌絲體在與化學物質相接觸或與其他微生物相互作用時,容易導致菌核的形成。GEORGIOU等[20-21]對菌核分化進行了深入研究,證明了真菌的菌核分化過程中伴隨著高度的脂質過氧化,活性氧(ROS)可以誘導菌核的分化。我們在培養基中添加能導致氧脅迫的外源物質,是否會對N1菌株的菌核分化有所影響以及是否可以進一步提高菌核生物量,還有待進一步研究。
[1]齊祖同.曲霉的分類鑒定[J].釀酒科技,1981,3(7):18-20.
[2]武漢大學,復旦大學.微生物學 [M].北京:人民教育出版社,1980.
[3]ROBERT K ANTIBUS.Formation and structure of sclerotia and scleotium-specific proteins in Hygrophoropsis aurantiaca[J].Mycologia,1989,81(6):905-913.
[4]KUO M J,ALEXANDER M.Inhibition of the lysis of fungi by melanins[J].Journal ofBacteriology,1967,94:624-629.
[5]COLEY-SMITH J R,COOKE R C.Survival and germination offungal sclerotia[J].Annual Reviews ofPhytopathology,1971,9:65-92.
[6]WILLETTS H J.The survival of fungal sclerotia under adverse environmental conditions[J].Biological Reviews,1971,46:387-407.
[7]EVANS J D,ARONSTEIN K,CHEN Y P,et a1.Immune pathways and defence mechanisms in honey bees Apis mellifera[J].Insect Molecular Biology,2006,15(5):645-656.
[8]沈萍,范秀榮,李廣武.微生物學實驗[M].北京:高等教育出版社,1996.
[9]趙文婧,張澤請,龍丹丹,等.湯姆青霉PT95和Q1菌株產菌核培養基篩選[J].山西大學學報,2013,36(4):622-627.
[10]齊祖同,孔華忠.中國真菌志(第五卷)[M].北京:科學出版社,1997.
[11]李智偉.一株產菌核青霉的分離鑒定及生物學特性研究[D].太原:山西大學,2010.
[12]徐軍,崔麗霞,韓建榮.無機鹽和碳氮源對青霉PT95類胡蘿產率的影響[J].微生物學通報,2003,30(4):77-81.
[13]韓建榮,李智偉,王宇,等.一株產菌核青霉的生物學特性研究[J].山西大學學報,2009,32(4):634-639.
[14]杜榮騫.生物統計學[M].北京:高等教育出版社,1985.
[15]韓建榮,姚鵬,戚敬,等.青霉PT95菌株滲出液的產生及理化性質的研究[J].山西大學學報,2007,30(2):274-278.
[16]韓建榮,王肖娟,原香娥.青霉PT95菌株菌核內產類胡蘿卜素的研究[J].微生物學通報,1998,25(6):319-321.
[17]韓建榮,徐軍.青霉PT95菌株固態發酵產生類胡蘿卜素的研究[J].微生物學報,1999,39(2):148-153.
[18]張澤清.氧脅迫和硒對湯姆青霉菌核分化和抗氧化性的影響研究[D].太原:山西大學,2014.
[19]CHET I,HENIS Y.Sclerotial morphogenesis in fungi[J].Annual Review ofPhytopathology,1975,13:169-192.
[20]GEORGIOU C D,PETROPOULOU K P.Effect of the antioxidant ascorbic acid on sclerotial differentiation in Rhizoctonia solani[J]. Plant Pathology,2001,50:594-600.
[21]SIDERI M,GEORGIOU C D.Differentiation and hydrogen peroxide production in Sclerotium rolfsii are induced by the oxidizing growth factors,light and iron[J].Mycologia,2000,92:1033-1042.
Study on Isolation and Identification of One Aspergillus Strain Producing Sclerotia and Analysis of Its Biological Characteristics
NINGLujuan1,HANJianrong2,AOXinyu1
(1.College ofLife Science,Southwest Forestry University,Kunming 650224,China;2.College ofLife Science,Shanxi University,Taiyuan 030006,China)
Seventy-eight Aspergillus strains were isolated from 20 forest soil samples.One of them could produce sclerotia,which was isolated from the cypress trees soil in Huaguang township,Xichong county,Nanchong city,Sichuan province,named N1.By observing the colonial,individual characteristics and the analysis of ITSrDNA sequence,N1 strain was initially identified to Aspergillus flavus var.flavus.The result ofbiological characteristics showed that the suitable medium for N1 strain producing sclerotia was CYA and CA.The optimum culture temperature for strain N1 was 30℃and the optimum pH was 6.Carbon and nitrogen sources in medium had certain effect on sclemtial biomass of strain N1.The best carbon source was sucrose and the best nitrogen source was peptone.The medium containing 9.88~39.53 g/L carbon and 0.42~0.84 g/L nitrogen,the sclerotia biomass of strain N1 was the largest.Inorganic salts also had obvious effects on sclerotial biomass ofstrain N1.
Aspergillus;sclerotium;ITS;biological characteristics
TQ925+.7
A
1002-2481(2017)03-0365-07
10.3969/j.issn.1002-2481.2017.03.12
2016-10-19
云南省優勢特色重點學科生物學一級學科建設項目(50097505)
寧露娟(1991-),女,山西太原人,在讀碩士,研究方向:生物化學與分子生物學。敖新宇為通信作者。
