姜荷花種球抗寒生理生化特征及促抗寒藥劑效果
2017-04-26 02:28:23劉建新徐笑寒丁華僑
浙江農業學報 2017年4期
劉建新,徐笑寒,丁華僑
(浙江省農業科學院 花卉研究開發中心,浙江 杭州 311202)
姜荷花種球抗寒生理生化特征及促抗寒藥劑效果
劉建新,徐笑寒,丁華僑
(浙江省農業科學院 花卉研究開發中心,浙江 杭州 311202)
姜荷花是國內新興的一種熱帶花卉,但中國絕大部分地區因冬季寒冷導致種球不能自然越冬保存。為了明確姜荷花種球的抗寒生理,以便找出進一步提升種球抗寒性的方法,用不同低溫脅迫處理姜荷花清邁粉種球,分析其各種生理生化指標,并開展了種球的抗寒藥劑篩選試驗。結果表明,姜荷花種球在9 ℃開始表現出逆境脅迫;由抗氧化酶系統和滲透調節系統的變化可知,種球分別在5和1 ℃調動了自身的抗逆系統來抵御環境的不良影響,在-3 ℃,各抗逆相關指標基本上降低到最低點。抗寒藥劑篩選試驗表明,各藥劑處理(包括脫落酸、水楊酸、蕓苔素內酯、6-BA、多效唑等)均對增強姜荷花種球的抗寒性有一定作用。根據細胞受損程度分析,1 mg·L-1ABA浸泡種球處理24 h的保護效果最好;其次,0.08 mg·L-16-BA處理對膜透性有較好的保護作用,但膜脂的氧化程度較高。
姜荷花;清邁粉;種球;抗寒性;脫落酸
姜荷花(CurcumaalismatifoliaGagnep)是國際上流行的主要姜荷屬花卉種類,為多年生球根草本花卉,原產于泰國清邁等地,主要作為佛教用花。由于不育苞片形似荷花且為姜科,故稱為姜荷花。具備荷花和郁金香雙重之美,又稱熱帶郁金香(Summer tulip)或暹羅郁金香(Siam tulip)。目前,國外有關姜荷花的研究主要集中在地理生態分布、采后生理、花發育、花期調控、組織培養、同工酶多態性、遺傳多樣性、基因克隆、遺傳轉化等。國內姜荷花相關研究相對較少,且主要集中在生產應用方面,如姜荷花的栽培、組培、切花保鮮[1]、矮化[2]、園林應用[3-4]等。至于抗寒性方面,林金水等[5]對姜荷花植株的抗寒性進行了研究,但有關姜荷花種球的抗寒性研究一直未見報道。由于姜荷花原產地在熱帶地區,種球一般都需要較高的溫度才能順利越冬,溫度較低會導致種球凍壞。在中國的絕大部分地區,姜荷花種球都無法自然越冬,因此,有必要使用抗寒相關藥劑來提升種球的抗寒能力。本研究選用了具有促抗寒潛力的脫落酸(abscisic acid,ABA)、水楊酸(salicylic acid,SA)、多效唑(paclobutrazol,PP333)、6-BA和油菜素內酯(brassinolide,BR)作為研究對象。ABA是抗寒基因表達的信號分子,對植物抗寒力的調控起重要作用,能提高植物的抗逆性[6-8]。SA是一種植物內源信號物質和植物激素,能抑制氣孔開放,從而使葉片具有抗蒸騰的作用,進而減少植物的水分蒸騰,提高抗逆性[9-10];PP333是一種高效、低毒的植物生長調節劑,可以抑制細胞縱向生長、縮短莖節、降低株高、改善植物群體結構、提高作物抗寒性、耐旱性和光合活性等[11-12]。6-BA的主要生理效應除促進細胞分裂、芽分化、細胞擴大和延緩葉片衰老外[13],也可提高植物的抗逆性[10]。如0.5 mg·L-1的6-BA誘導香根草能提高香根草的抗寒性,使得香根草能在低溫下生長良好[14]。BR浸種能增強花生對低溫的忍耐能力[15-16]。
本實驗通過研究姜荷花種球的抗寒生理,明確種球能耐受的低溫閾值,并采用具有抗寒潛力的各種藥劑處理種球,以期找出能提升姜荷花種球抗寒性的方法,為中國姜荷花種球生產提供支持。
1 材料與方法
1.1 試驗材料
采用浙江農業科學院花卉研究開發中心生產的大小一致、無病蟲害及外傷的清邁粉姜荷花種球。取樣時,從種球的中間位置縱切開,從中切取完整小片(包含外中內各層組織,確保取樣代表性)。MDA含量、SOD活性、POD活性、CAT活性、脯氨酸含量、可溶性糖含量、蛋白質含量的測定用試劑盒均購自蘇州科銘生物技術有限公司。
1.2 試驗方法
1.2.1 低溫脅迫處理
目前,生產中姜荷花種球一般在15 ℃左右的環境越冬,但在中國中部及南部大部分地區,冬季氣溫一般最低可以達到-3~-2 ℃(包括浙江省)。本試驗以24 ℃為常溫對照組,以-3 ℃為最低點,每隔4 ℃設1個基點,將種球分別置于24、9、5、1、-3 ℃暗處理48 h,然后取出存放于-80 ℃超低溫冰箱備用。
1.2.2 藥劑處理
將姜荷花種球用不同藥劑分別浸泡處理24 h,用清水沖洗干凈,再1 ℃低溫脅迫48 h,然后取出存放于-80 ℃超低溫冰箱凍存備用。試驗藥劑包括:1 mg·L-1脫落酸(ABA)、11 mg·L-1或80 μmol·L-1水楊酸(SC)、0.01 mg·L-1油菜素內酯(BR)、0.08 mg·L-16-BA、100 mg·L-1多效唑(PP333),各藥劑濃度設定參照其他作物相關研究中最常見的濃度,并作適當調整。另設對照組(CK):清水浸泡24 h后,1 ℃低溫脅迫48 h,凍存備用。
1.2.3 生理生化指標檢測
相對電導率的測定:將種球切塊,用去離子水室溫下浸泡12 h,沸水加熱30 min后再冷卻至室溫。采用電導儀測定沸水加熱前后的電導率,然后再計算出相對電導率。
束縛水的測定采用阿貝折光儀法[17]。用折光儀測定經蔗糖透析前后的濃度,再計算出束縛水的含量。
MDA含量、SOD活性、POD活性、CAT活性、脯氨酸含量、可溶性糖含量、蛋白質含量的測定均按照試劑盒說明書方法進行。所有測定采用同批次處理種球,進行3次平行試驗。
1.3 數據分析
試驗數據用Excel 2007軟件分析并作圖。
2 結果與分析
2.1 低溫脅迫下姜荷花種球的生理生化變化
2.1.1 種球的相對電導率、MDA和束縛水含量
如圖1-A所示,姜荷花種球在24 ℃的相對電導率最低,9 ℃時電導率顯著升高,說明9 ℃時種球已處于受低溫脅迫的狀態;與9 ℃相比,1、5 ℃時,種球的相對電導率略有降低,-3 ℃時,種球的相對電導率升高,表明-3 ℃時姜荷花種球細胞膜的受損最嚴重。如圖1-B所示,除對照組外,束縛水含量隨著溫度的降低而逐漸升高,在-3 ℃達到最高。低溫脅迫處理后,MDA含量明顯增加,在-3 ℃時達到最高,表明此時細胞膜脂過氧化程度最高,受損最嚴重(圖1-C)。結合這3個指標可知,種球在9 ℃已經處于逆境脅迫狀態,在試驗溫度范圍內,-3 ℃時姜荷花種球的受脅迫程度達到最高。
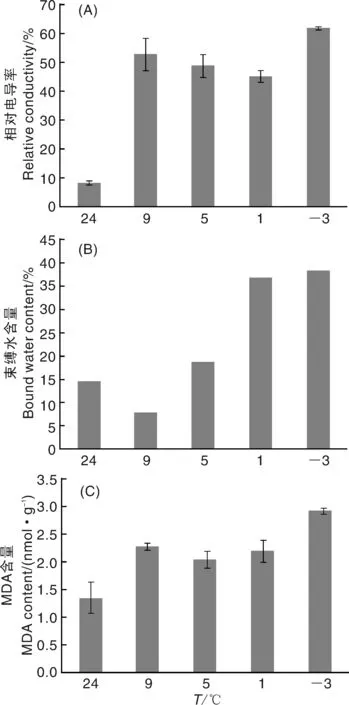
圖1 低溫脅迫下姜荷花種球的相對電導率、束縛水含量和MDA含量Fig.1 Relative conductivity, bound water content and MDA content of C. alismatifolia bulbs under low temperature stress
2.1.2 種球的SOD、POD和CAT活性
由圖2-A可知,低溫脅迫時,姜荷花種球SOD活性沒有表現出明顯的規律性,與對照相比,在5、-3 ℃時,SOD活性顯著增加,9和1 ℃時SOD活性與對照相當。在9~-3 ℃的低溫脅迫處理過程中,POD活性呈先增加后降低的趨勢,1 ℃達到最高,在-3 ℃時降到最低(圖2-B)。與POD活性類似,在9~-3 ℃的低溫脅迫處理過程中,CAT活性也呈先增加后降低的趨勢,1 ℃時CAT活性最高(圖2-C)。由POD和CAT活性變化可知,-3 ℃時,姜荷花種球已經失去抵抗逆境的能力。綜上,9 ℃時,SOD、POD和CAT活性均沒有表現出明顯的脅迫響應,而在5 ℃時,3個抗氧化酶的活性都大幅增加,說明溫度降至5 ℃時,種球已經開始啟動抗逆響應機制。
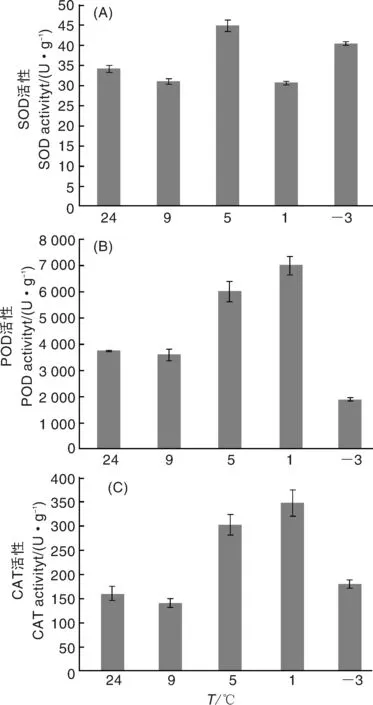
圖2 低溫脅迫下姜荷花種球的抗氧化酶活性Fig.2 Antioxidant enzyme activities of C. alismatifolia bulbs under low temperature stress
2.1.3 種球的脯氨酸、可溶性糖和可溶性蛋白含量
如圖3-A所示,在24~-3 ℃,隨著溫度的降低,脯氨酸含量先增加后降低,5 ℃時達到最高,然后逐漸下降,-3 ℃時達到最低。與對照組相比,各處理的可溶性糖含量變化不明顯,1 ℃時可溶性糖含量最高,-3 ℃時可溶性糖含量最低(圖3-B)。在9~-3 ℃,隨著溫度的降低,可溶性蛋白含量逐漸增加,1 ℃時達到最高,溫度低于1 ℃時可溶性蛋白含量又下降(圖3-C)。

圖3 低溫脅迫下姜荷花種球細胞滲透調節系統的變化Fig.3 Changes of osmotic adjustment system of C. alismatifolia bulbs cell under low temperature stress
2.2 藥劑處理對姜荷花種球抗寒性的影響
2.2.1 不同藥劑處理對低溫脅迫下種球的相對電導率、MDA和束縛水含量的影響
從圖4-A中可以看出,1 ℃處理48 h后,對照組種球的相對電導率最高,各藥劑處理組種球的相對電導率均顯著低于對照,其中,6-BA處理的種球相對電導率下降幅度最大。如圖4-B所示,1 ℃處理48 h后,與對照組相比,各藥劑處理組種球的束縛水含量均有不同幅度的增加,其中SC和6-BA處理的增加幅度最大。1 ℃處理48 h后,與對照組相比,6-BA處理的種球MDA含量顯著增加,ABA處理的種球MDA含量顯著下降,SC、BR和PP333處理的種球MDA含量略有下降,但差異不顯著(圖4-C)。
醫療體制逐步改革,法律法規逐漸完善,尊重和保護病人權益的呼聲越來越大,導致醫患矛盾加劇,病人往往不配合床邊教學。病人周轉快、住院天數縮短等也使得可供選擇的床邊教學病人減少。
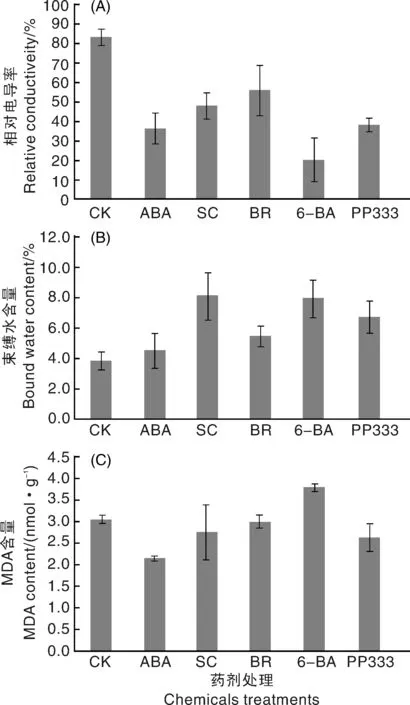
圖4 低溫脅迫下藥劑處理后姜荷花種球的相對電導率、束縛水含量和MDA含量Fig.4 Effects of different chemicals treatments on relative conductivity, bound water content and MDA content of C. alismatifolia bulbs under low temperature stress
2.2.2 不同藥劑處理對低溫脅迫下種球SOD、POD和CAT活性的影響
如圖5-A所示,1 ℃處理48 h后,與對照組相比,各藥劑處理的姜荷花種球SOD活性均顯著增強,其中,6-BA處理的姜荷花種球SOD活性增加幅度最大,其次為BR和PP333處理。除6-BA處理外,各處理的POD活性均較對照組有顯著增加,SC處理的增加幅度最大,其次是BR、PP333和ABA(圖5-B)。除6-BA處理外,各處理的CAT活性均有顯著增加,ABA和PP333處理的姜荷花種球CAT活性最高(圖5-C)。由此可知,姜荷花種球受低溫脅迫時,不同藥劑處理的效果不同,如ABA處理可以提高CAT活性,SC處理可以提高POD和CAT活性,BR處理可同時提高SOD、POD和CAT活性;6-BA處理可以顯著提高姜荷花種球SOD活性,但同時抑制了POD和CAT活性。
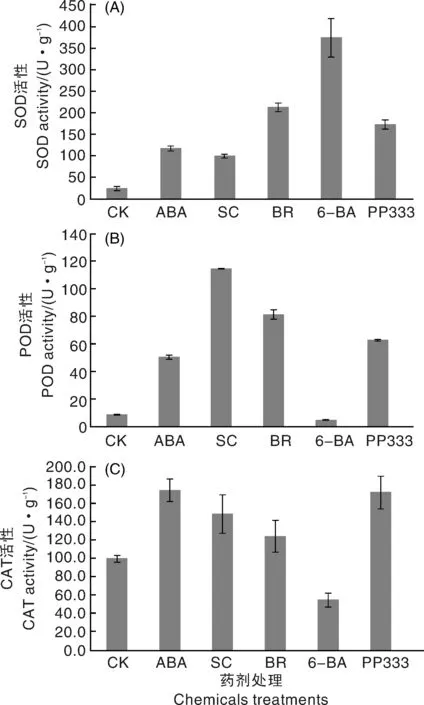
圖5 低溫脅迫下藥劑處理對姜荷花種球抗氧化酶活性的影響Fig.5 Effects of different chemicals treatments on antioxidant enzyme activities of C. alismatifolia bulbs under low temperature stress
為分析各藥劑處理對抗氧化酶系統的總體效用,將SOD、POD和CAT活性各按1∶1∶1均等權重累加,將對照組的總抗氧化酶活性定為1.0,構建1個不同藥劑處理的總抗氧化酶活性圖(圖6)。由圖6可知,與對照組相比,各處理的總抗氧酶活性均明顯增加,BR、SC、6-BA、PP333和ABA處理組的總抗氧酶活性分別是對照組的6.2、6.0、5.4、5.2和4.0倍。說明各藥劑處理均能促進抗氧化酶的生成,但生成的的抗氧化酶種類各有側重。
2.2.3 不同藥劑處理對低溫脅迫下種球脯氨酸、可溶性糖和可溶性蛋白含量的影響
1 ℃處理48 h后,除PP333處理外,各藥劑處理的姜荷花種球脯氨酸含量均較對照組高,其中,ABA和BR處理組脯氨酸含量最高(圖7-A)。與對照組相比,各藥劑處理的可溶性糖含量均升高,其中ABA處理的可溶性糖含量最高(圖7-B)。如圖7-C所示,除6-BA處理外,其他藥劑處理的可溶性蛋白含量均較對照高,其中SC處理的蛋白質含量最高,其次為BR、ABA處理。
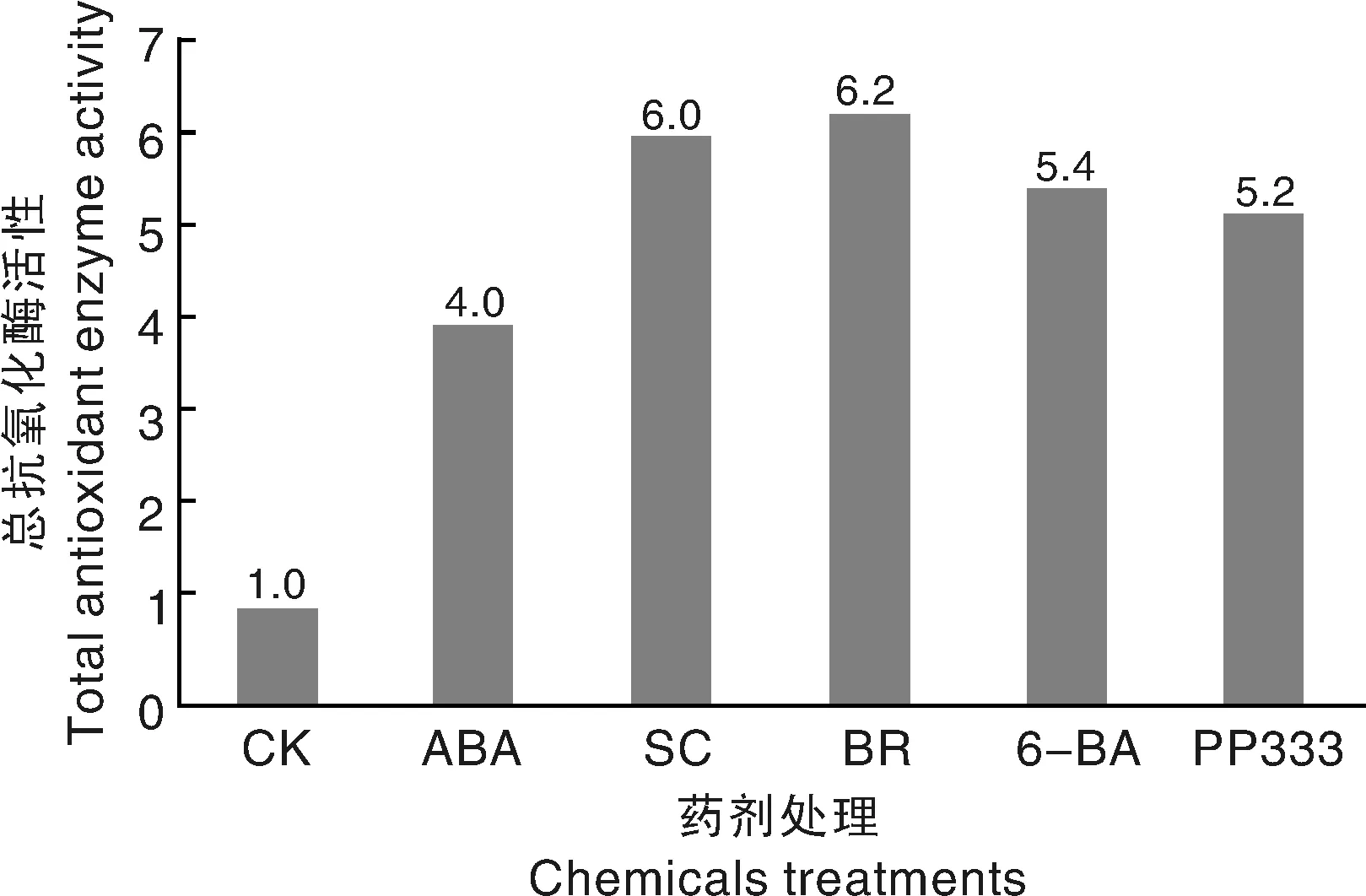
圖6 低溫脅迫下不同藥劑處理對種姜荷花種球抗總氧化酶活性的影響Fig.6 Effects of different chemicals on total antioxidant enzyme activity of bulbs under low temperature stress

圖7 低溫脅迫下不同藥劑處理對種球細胞滲透調節系統的影響Fig.7 Effects of different chemicals treatments on cell osmotic regulation of C. alismatifolia bulbs under low temperature stress
3 結論與討論
3.1 低溫脅迫下姜荷花種球的生理生化變化
植物受到低溫脅迫時,細胞膜系統最先受到影響[18],質膜透性增大,電解質外滲,導致電導率會有不同程度的增加。膜透性的大小反映了細胞膜受損的程度,相對電導率與膜受傷害程度呈正相關,電導率越高說明細胞膜受到的傷害越大[19]。束縛水是指植物體內和原生質膠體緊密結合的水分,其含量相對增加可以增強植物的抗寒能力[20]。MDA含量是膜脂過氧化的指標,可以反映植物膜脂過氧化程度及對逆境反應的強弱[21-22]。本研究中,9 ℃時,姜荷花種球的相對電導率和MDA含量均顯著增加,說明9 ℃左右時,姜荷花種球已經受到脅迫了。逆境條件下,植物本身具備抵抗逆境的防御系統,抗氧化酶系統和滲透調節系統是植物常用的2套防御系統。抗氧化酶系統是清除活性氧和自由基的保護酶系統,可清除脅迫條件下植物體積累的活性氧自由基,降低自由基對膜系統造成的傷害,保護細胞膜結構和功能[23-24],抗氧化酶包括過氧化氫酶、過氧化物酶、超氧化物歧化酶等[17]。滲透調節系統是逆境條件下,植物通過滲透調節保證一定的膨壓來維持細胞正常的生理功能[25]。遭受環境低溫脅迫時,大多數植物可在體內積累脯氨酸、可溶性蛋白及可溶性糖等物質[26],來降低細胞液的滲透勢,防止細胞過度失水,提高植物的抗寒性[27]。由抗氧化酶系統和滲透調節系統變化可知,5和1 ℃時,姜荷花種球已經調動了自身的抗逆系統來抵御低溫,-3 ℃時,各抗逆相關指標基本上降低到最低點,表明此時的低溫已超出種球的耐受極限。
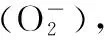
3.2 不同藥劑處理對姜荷花種球抗寒性的影響
不同藥劑處理對姜荷花種球抗氧化酶的影響不同,如6-BA處理可顯著提高SOD活性,而POD和CAT活性卻低于對照。由SOD、POD和CAT的總體活性可知,各藥劑處理均能明顯提高總抗氧化酶活性,其中以BR處理的效果最顯著。
從相對電導率、MDA含量指標來看,1 mg·L-1ABA對種球的保護效果最好;0.08 mg·L-16-BA處理雖然對姜荷花種球膜透性有較好的保護作用(相對電導率最低),但其膜脂氧化程度卻較高(MDA含量最高);11 mg·L-1SC處理的種球抗寒性較0.01 mg·L-1BR和100 mg·L-1PP333好,BR和PP333處理效果相當。由姜荷花種球抗氧化酶系統及滲透調節系統可知,ABA主要通過滲透調節系統來提高種球抗寒性,抗氧化酶系統的作用沒有其他藥劑處理明顯,這與ABA處理產生的MDA含量低有一定關系。與此相反,6-BA處理的種球膜脂氧化較嚴重(MDA含量較高),總抗氧化酶活性也相當高,約是對照組的5.4倍,但滲透調節系統變化不明顯。此外,雖然1 mg·L-1ABA和0.08 mg·L-16-BA對種球的保護效果較好,但由于這個濃度是參照其他植物的研究而確定的,對姜荷花種球未必是最理想的濃度,因此,后續試驗需要針對姜荷花種球進行濃度篩選,以確保獲得最佳的抗寒效果。
[1] 劉建新, 丁華僑, 李明江, 等. 姜荷屬花卉的切花保鮮試驗[J]. 浙江農業科學, 2015, 56(6): 890-892. LIU J X, DING H Q, LI M J, et al. Study on fresh-keeping technology ofCurcumacut flowes [J].JournalofZhejiangAgriculturalSciences, 2015, 56(6): 890-892. (in Chinese)
[2] 丁華僑, 劉建新, 王煒勇, 等. 烯效唑對姜荷花的矮化效果及組織解剖研究[J].浙江農業學報, 2013, 25(5): 975-979. DING H Q, LIU J X, WANG W Y, et al. Dwarfing effect of uniconazole onCurcumaalismatifoliaand its tissue anatomy [J].ActaAgriculturaezhejiangensis, 2013, 25(5): 975-979. (in Chinese with English abstract)
[3] 丁華僑, 劉建新, 鄒清成, 等. 姜荷屬花卉資源及園林應用分析[J]. 現代園林, 2014, 11(8): 85-88. DING H Q, LIU J X, ZOU Q C, et al. Germplasm resources and garden applications ofCurcumalinnaeus[J].ModernLandscapeArchitecture, 2014, 11(8): 85-88. (in Chinese with English abstract)
[4] 丁華僑, 劉建新, 王煒勇, 等. 不同植物生長延緩劑對姜荷花的矮化效果[J]. 浙江農業科學, 2013 (5): 559-562 DING H Q, LIU J X, WANG W Y, et al. Dwarfing effect of different plant growth retardants onCurcumaalsimatifolia[J].JournalofZhejiangAgriculturalSciences, 2013 (5): 559-562. (in Chinese)
[5] 林金水, 陸鑾眉, 趙倩倩. 姜荷花抗寒性的研究初探[J]. 福建熱作科技, 2013, 38(2): 1-5. LIN J S, LU L M, ZHAO Q Q. Preliminary study on the cold resistance ofCurcumaalsimatifolia[J].FujianScienceandTechnologyofTropicalCrops, 2013, 38(2): 1-5. (in Chinese)
[6] 趙春江, 康書江, 王紀華, 等. 植物內源激素與不同基因型小麥抗寒性關系的研究[J]. 華北農學報, 2000, 15(3): 51-54. ZHAO C J, KANG S J, WANG J H, et al. Study on relations between plant endogenous hormones and cold resistance in wheat [J].ActaAgricuhturaeBoreahi-Sinica, 2000, 15(3): 51-54. (in Chinese with English abstract)
[7] 盧少云, 郭振飛. 草坪草逆境生理研究進展[J]. 草業學報, 2003, 12(4): 7-13. LU S Y, GUO Z F. Physiological responses of turfgrass to abiotic stresses [J].ActaPrataculturaeSinia, 2003, 12(4): 7-13. (in Chinese with English abstract)
[8] 劉海卿, 方園, 武軍艷, 等. 低溫脅迫下內源ABA、GA及比值對白菜型和甘藍型冬油菜抗寒性的響應[J]. 中國生態農業學報, 2016, 24(11): 1529-1538. LIU H Q, FANG Y, WU J Y, et al. Response of endogenous ABA and GA to cold resistance ofBrassicarapaL. andBrassicanapusL. [J].ChineseJournalofEco-Agriculture, 2016, 24(11): 1529-1538. (in Chinese with English abstract)
[9] 李才生, 秦燕, 宗盼. 外源水楊酸對玉米幼苗生理特性的影響[J]. 玉米科學, 2010, 18(3): 98-104. LI C S, QIN Y, ZONG P. The effect of exogenous salicylic acid on physiological characteristics of maize seedling [J].JournalofMaizeSciences, 2010, 18(3): 98-104. (in Chinese with English abstract)
[10] 方基建. 磷鉀及6_BA處理對絲瓜幼苗抗寒性的影響[D]. 合肥: 安徽農業大學, 2010. FANG J J. Effect of phosphorus, potassium and 6-BA treatments on cold resistance of towelgourd (luffa) seedling [D]. Hefei: Anhui Agricultural University, 2010. (in Chinese with English abstract)
[11] 劉波, 劉志雄. 多效唑對幾個狗牙根品種抗寒性的影響[J]. 安徽農學通報, 2007, 13(12): 36-37. LIU B, LIU Z X. Effects of paclobutrazal on physiological response of several bermudagrass cultivars to cold stress [J].AnhuiAgriculturalScienceBulletin, 2007, 13(12): 36-37. (in Chinese with English abstract)
[12] 黃國弟. 乙烯利、多效唑在生產中的應用[J]. 廣西熱帶農業, 2005 (3): 17. HUANG G D. The application of ethephon and paclobutrazol in production [J].GuangxiTropicalAgriculture, 2005 (3): 17. (in Chinese)
[13] 王忠. 植物生理學[M]. 北京: 中國農業出版社, 2000.
[14] YANG B B, WU G J, MA Z R, et al. Efficient regeneration system and agrobacterium-mediated transformation ofVetiveriazizanioides(L.) Nash [J].PakistanJournalofBotany, 2008, 4(2): 911-921.
[15] 董登峰, 李楊瑞, 江立庚, 等. 長效油菜素內酯TS303和二氫茉莉酸丙酯增強花生抗寒能力[J]. 廣西植物, 2008, 28(5): 675-680. DONG D F, LI Y R, JIANG L G, et al. Effects of long-lasting brassinosteroid TS303 and propyl dihydrojasmonate on enhancing peanut resistance to chilling [J].Guihaia, 2008, 28(5): 675-680. (in Chinese with English abstract)
[16] 許紹惠, 韓忠環, 何若韞. 油菜素內酯對白皮松幼苗抗寒性的生理效應[J]. 沈陽農業大學學報, 1991, 22(2): 123-127. XU S H,HAN Z H, HE R Y. Physiological effects of brassinolide (BR) on cold resistance inPinusbungeanaseedlings [J].JournalofShenyangAgriculturalUniversity, 1991, 22(2): 123-127. (in Chinese with English abstract)
[17] 王依, 靳娟, 羅強勇, 等. 4個釀酒葡萄品種抗寒性的比較[J]. 果樹學報, 2015 (4): 612-619. WANG Y, JIN J, LUO Q Y, et al. Comparison of cold resistance for 4 grapewine cultivar [J].JournalofFruitScience, 2015 (4): 612-619. (in Chinese with English abstract)
[18] 李瓊, 劉國道, 郇樹乾. 鹽脅迫下六種禾本科牧草幼苗葉片膜脂過氧化作用及其與耐鹽性的關系[J]. 家畜生態學報, 2005, 26(5): 63-67. LI Q, LIU G D, XUN S Q. The activities of protective enzymes of grass seedlings to long salinity and their relationship to salinity tolerance [J].JournalofDomesticAnimalEcology, 2005, 26(5): 63-67. (in Chinese with English abstract)
[19] 雷紅靈, 胡雪雷, 吳永堯. 硒對恩施碎米薺葉片抗氧化酶活性的影響[J]. 華中科技大學學報, 2010, 3(38): 78-85. LEI H L, HU X L, WU Y Y. Effect of selenium on the antioxidant enzyme activity ofCardamineenshiensis’ leaves [J].JournalofHuazhongUniversityofScienceandTechnology, 2010, 3(38): 78-85. (in Chinese with English abstract)
[20] 孫玉潔, 王國槐. 植物抗寒生理的研究進展[J]. 作物研究, 2009, 2(5): 293-297. SUN Y J, WANG G H. The research progress of plant cold-resistance physiology [J].CropResearch, 2009, 2(5): 293-297. (in Chinese)
[21] 梅映學, 魏瑋, 張詩婉, 等. 干旱鍛煉對鹽脅迫下水稻幼苗根系抗氧化酶活性的影響[J]. 浙江農業學報, 2016, 28(8): 1304-1308. MEI Y X, WEI W, ZHANG S W, et al. Effect of PEG pretreatment on antioxidant enzymes activity under salt stress in root of rice seeding [J].ActaAgriculturaeZhejiangensis, 2016, 28(8): 1304-1308. (in Chinese with English abstract)[22] 劉錦川, 云錦鳳, 張磊. 氯化鈉脅迫下3種披堿草屬牧草生理特性的研究[J]. 草地學報, 2010, 18(5): 694-697. LIU J C, YUN J F, ZHANG L. Physiological characteristics of threeElymusgrass under NaCl stress [J].ActaAgrectirSinica, 2010, 18(5): 694-697. (in Chinese with English abstract)
[23] 朱政, 蔣家月, 江昌俊, 等. 低溫脅迫對茶樹葉片SOD、可溶性蛋白和可溶性糖含量的影響[J]. 安徽農業大學學報, 2011, 38(1): 24-26. ZHU Z, JIANG J Y, JIANG C J, et al. Effects of low temperature stress on SOD activity, soluble protein content and soluble sugar content inCamelliasinensisleaves [J].JournalofAnhuiAgriculturalUniversity, 2011, 38(1): 24-26. (in Chinese with English abstract)
[24] 房用, 李秀芬, 慕宗昭. 茶樹抗寒性研究進展[J]. 經濟林研究, 2004, 22(2): 69-72. PANG Y, LI X F, MU Z Z, et al. Development of research on anti-frigidity of tea tree [J].EconomicForestResearches, 2004, 22(2): 69-72. (in Chinese with English abstract)
[25] 閆洪奎, 劉祥, 王廣東,等. 低鉀脅迫下耐低鉀玉米可溶性蛋白、可溶性糖和鉀含量的變化及其關系[J]. 玉米科學, 2012, 20(6): 81-84. YAN H K, LIU X, WANG G D, et al. Changes and relation of soluble protein, soluble sugar and potassium in low-potassium tolerant maize under low-potassium condition [J].JournalofMaizeSciences, 2012, 20(6): 81-84. (in Chinese with English abstract)
[26] 朱金方, 陸兆華, 夏江寶, 等. 鹽旱交叉脅迫對檉柳幼苗滲透調節物質含量的影響[J]. 西北植物學報, 2013, 33(2): 357-363. ZHU J F, LU Z H, XIA J B, et al. Changes of osmotic adjusting substances in leaves ofTamarixchinensisseedlings under salt and drought stress[J].ActaBotanicaBoreali-OccidentaliaSinica, 2013, 33(2): 357-363. (in Chinese with English abstract)
[27] 閆世江, 張繼寧, 劉潔, 等. 茄子幼苗耐低溫性生理機制研究[J]. 西北植物學報, 2011, 31(12): 2498-2502. YAN S J, ZHANG J N, LIU J, et al. Physiological mechanism of chilling tolerance in eggplant seedling [J].ActaBotanicaBoreali-OccidentaliaSinica, 2011, 31(12): 2498-2502. (in Chinese with English abstract)
[28] 邱乾棟, 呂曉貞, 臧德奎, 等. 植物抗寒生理研究進展[J]. 山東農業科學, 2009 (8): 53-57. QIU Q D, LU X Z, ZANG D K, et al. Research progress on plant physiology of cold resistance[J].ShandongAgriculturalSciences, 2009 (8): 53-57. (in Chinese with English abstract)
[29] 田延亮, 蔡梓林, 季明芳. 油菜超氧物歧化酶活性及其同工酶的研究[J]. 中國油料作物學報, 1988 (1): 31-35. TIAN Y L, CAI Z L, JI M F. A study on the superoxide activity and its isoenzymes in rape [J].ChineseJournalofOilCropSciences, 1988 (1):31-35. (in Chinese)
(責任編輯 侯春曉)
Physiological and biochemical characteristics ofCurcumaalismatifoliabulbs and effects of some chemicals on its cold resistance
LIU Jianxin, XU Xiaohan,DING Huaqiao
(ResearchandDevelopmentCentreofFlower,ZhejiangAcademyofAgriculturalSciences,Hangzhou311202,China)
Curcumaalismatifoliais a new kind of tropical flowers. However, winter in most areas in China is too cold to preserve bulbs naturally. In order to clarify cold-resistance physiology ofCurcumaalismatifoliabulbs, and to find out methods to enhance bulbs cold-resistance, various physiological and biochemical indexes of bulbs which were treated with different low temperature stress were detected, and cold-resistance chemicals screening experiment for bulbs were carried out. The results of physiological and biochemical tests showed thatCurcumaalismatifoliabulbs began to be stressed at 9 ℃. According to changes of antioxidant enzyme system and osmotic adjustment system, these bulbs mobilized their own stress-resistant system to resist adverse effects at 5 and 1 ℃, respectively, While various stress-resistant related indexes reduced to the lowest point at -3 ℃, which might be beyond the tolerance limit of bulbs. The cold-resistant chemicals screening experiment showed that all chemicals treatments including abscisic acid, salicylic acid, brassinolide, 6-BA, paclobutrazol could enhance cold tolerance of bulbs. According to cell damage degree analysis, 1 mg·L-1ABA immerged bulbs for 24 h had the best protective effect to bulbs. 0.08 mg·L-16-BA treatment also could protect membrane permeability, but the oxidation degree of membrane lipid was very high.
Curcumaalismatifolia; Chiang Mai Pink; bulb; cold resistance; abscisic acid
http://www.zjnyxb.cn
10.3969/j.issn.1004-1524.2017.04.09
2016-11-03
浙江省農業科學院青年人才培養項目(重大課題主持能力培養類);浙江省農業科學院農業科技發展專項;浙江省農業科學院科技產業化資金項目;杭州市科技計劃引導項目(20163501Y78)
劉建新(1980—),男,湖南祁陽人,助理研究員,博士,主要從事花卉育種和栽培研究。E-mail: liujianxin2000@aliyun.com
S682.2+9
A
1004-1524(2017)04-0575-08
浙江農業學報ActaAgriculturaeZhejiangensis, 2017,29(4): 575-582
劉建新, 徐笑寒, 丁華僑. 姜荷花種球抗寒生理生化特征及促抗寒藥劑效果[J]. 浙江農業學報, 2017, 29(4): 575-582.
