北京西山靜福寺地區(qū)鳥類多度分布格局
2017-05-15 12:30:16陳卓琳賈麗麗關(guān)文彬
生態(tài)學(xué)報 2017年6期
陳卓琳,賈麗麗,關(guān)文彬
北京林業(yè)大學(xué),北京 100083
北京西山靜福寺地區(qū)鳥類多度分布格局
陳卓琳,賈麗麗,關(guān)文彬*
北京林業(yè)大學(xué),北京 100083
物種多度格局分析對理解群落結(jié)構(gòu)具有重要意義。采用分割線段模型對北京西山靜福寺地區(qū)鳥類群落物種數(shù)量關(guān)系進(jìn)行擬合研究,并運用秩相關(guān)性分析對模型進(jìn)行相關(guān)系數(shù)檢驗,結(jié)果表明:(1)該地區(qū)鳥類群落多度格局模型與分割線段模型顯著相關(guān),利用分割線段模型可以良好的擬合該地區(qū)鳥類群落多度格局;(2)鳥類多度分布格局具有時間和空間的雙重屬性;(3)月尺度上,1月與其他月份差異顯著,森林群落間差異不顯著;(4)季尺度上,冬季與春、秋兩季差異顯著,落葉闊葉林與針闊混交林差異顯著,針葉林與落葉闊葉林、針闊混交林差異都不顯著;(5)年尺度上,年變化差異不顯著。
鳥類群落;多度格局;分割線段模型
在生物多樣性研究中,除物種多樣性指數(shù)外,還包括“物種相對多度模型”。該模型是利用數(shù)學(xué)統(tǒng)計方法描述物種多度的分布[1],也是某些物種多樣性指數(shù)應(yīng)用的基礎(chǔ)[2-3]。物種的多度格局是由同一群落的多個物種不斷相互作用、相互影響的結(jié)果,同時也是物種關(guān)系和作用機制的表征與體現(xiàn)[4- 6]。如果說,多樣性指數(shù)是群落結(jié)構(gòu)的數(shù)量化表征,那么群落多度格局分析就是對群落性質(zhì)的進(jìn)一步闡釋,兩者相輔相成[3]。多度不僅是確定物種保護(hù)等級的基本依據(jù),而且在生物多樣性保護(hù)和管理上有重要意義。此外,對于認(rèn)識一個群落而言,多度格局比多樣性指數(shù)更加直觀地體現(xiàn)群落特征[7]。為了更好的分析多度格局,研究者根據(jù)生態(tài)學(xué)原理構(gòu)建了各種關(guān)于多度格局的模型[3,8]。例如基于生態(tài)位理論的幾何級數(shù)模型、分割線段模型、重疊生態(tài)位模型等[5,9-10]。此外也有用統(tǒng)計模型研究多度格局,例如對數(shù)級數(shù)模型、對數(shù)正態(tài)分布模型等,但有些模型的擬合效果雖然很好,但無助于闡釋物種的作用過程和群落的裝配機制[5-6]。鳥類研究在保護(hù)生態(tài)環(huán)境和生物多樣性等方面發(fā)揮著重要作用。我國有豐富的鳥類資源,鳥類物種占世界鳥類物種總數(shù)的13.1%。國內(nèi)外對鳥類群落組成、動態(tài)及生物多樣性的研究已非常深入。但是,很少有從多度格局角度對鳥類群落進(jìn)行闡釋[11]。西山地區(qū)植被豐富,為森林鳥類提供天然棲息地,鳥類物種豐富,目前已有學(xué)者對該地鳥類群落的生物多樣性和種間聯(lián)結(jié)進(jìn)行研究[12-13]。本研究將從多度格局的角度,應(yīng)用多度格局模型,對北京西山靜福寺地區(qū)鳥類群落多度格局進(jìn)行研究,為研究和保護(hù)鳥類物種多樣性提供理論依據(jù)。
1 研究方法
研究地概況與鳥類調(diào)查方法參見文獻(xiàn)[12-13]。
1.1 物種多度指標(biāo)的選用
多度的測度分為絕對多度和相對多度。絕對多度是指群落中物種種數(shù)、頻度等指標(biāo)的絕對值;相對多度是指物種對群落總多度的貢獻(xiàn)大小,或稱為相對重要性百分率[14-15],同樣可以用來擬合各物種多度模型[16- 18]。May提出不同種的相對多度是群落模式的一個更為技術(shù)性的方面[19]。因此本文采用相對多度為指標(biāo)來擬合鳥類多度分布。觀測數(shù)據(jù)鳥類物種相對多度A為:
A=N/n×100%
(1)
式中,A為物種a的多度,N為物種a的個體數(shù),n為樣點中總個體數(shù)。
1.2 常用的多度格局模型
常用的多度格局模型可以劃分為兩類:統(tǒng)計類模型(如對數(shù)正態(tài)模型和對數(shù)級數(shù)模型等)和生態(tài)位模型(如幾何級數(shù)模型-生態(tài)位優(yōu)先占領(lǐng)假說和分割線段模型-隨機生態(tài)位假說)[3]。
對數(shù)正態(tài)模型是Preston[20]引入物種多度研究的對數(shù)正態(tài)分布形式:
S(R)=S0exp (-λ2R2)
(2)
式中,S(R)為第R個倍程物種的數(shù)量;S0為模擬模型中的總物種數(shù)的估計值;λ為正態(tài)分布曲線的寬度的倒數(shù),是一個參數(shù)。
對數(shù)級數(shù)模型是Fisher等在研究昆蟲物種多度分布時提出的[21],這一模型對有r個個體的中的頻率預(yù)測為:
(3)
式中,α代表了群落的特點,反映群落的多樣性;x是一個與調(diào)查樣方有關(guān)的參數(shù),其值與樣方大小有關(guān)。
幾何級數(shù)模型又稱生態(tài)位優(yōu)先模型,最早是由Motomura在1932年提出的,后經(jīng)Whittake[22-23]研究后才逐漸完善。模型中第i個種的多度所占比例期望值Pi為:
Pi=K×(1-K)i- 1
(4)
式中,i為觀察群落的物種數(shù);K對于每個群落是一定值。
分割線段模型又稱隨機生態(tài)位假說,是MacAthur在研究鳥類物種多度分布時提出的[24],模型中第i個物種的期望個體數(shù)所占比例Pi為:
(5)
式中,i為觀察群落的物種數(shù),S為物種總數(shù)。
1.3 數(shù)據(jù)處理
計算每個物種的相對多度,以物種從常見到稀有的順序為橫坐標(biāo),以物種相對多度(頻度)為縱坐標(biāo),繪制生態(tài)模型擬合圖。利用R語言3.2.3軟件Vegan數(shù)據(jù)包(http://mirror.bjtu.edu.cn/cran/)進(jìn)行模型比較。利用SPSS 19.0軟件與模型值進(jìn)行相關(guān)性分析檢驗,計算觀測值與模型值的相關(guān)性。對圖像線性回歸,構(gòu)建秩相關(guān)方程,利用方程斜率分析物種多度格局變化。Spearman秩相關(guān)系數(shù)Rs屬于非參數(shù)檢驗,表達(dá)式[25]為:
(6)
dk=(Xik-Xi)(Xjk-Xj)
(7)
式中,Xik、Xjk分別為中i、j在樣方k中的秩。
所繪圖像與對數(shù)曲線類似,計算曲線的斜率參數(shù)。方程在x0點的斜率即該方程在x0點的導(dǎo)數(shù),斜率為k=b/(x0ln10)[26]。
2 結(jié)果與分析
對54組多度數(shù)據(jù)分別進(jìn)行對數(shù)正態(tài)分布模型、對數(shù)級數(shù)模型、幾何級數(shù)模型和分割線段模型擬合度檢驗。得到各模型的平均偏差值。統(tǒng)計模型中,對數(shù)正態(tài)分布模型偏差值為0.063,對數(shù)級數(shù)模型偏差值為0.103。可見統(tǒng)計模型中,對數(shù)正態(tài)分布模型更適合描述該地多度格局。生態(tài)位模型中,幾何級數(shù)模型偏差值為0.404,分割線段模型偏差值為0.174。可見生態(tài)位模型中,分割線段模型更適合描述該地多度格局。由于生態(tài)位模型傾向于揭示群落多度格局的機制。而統(tǒng)計模型主要是描述,一般很難把生態(tài)學(xué)過程與之聯(lián)系起來[3]。如幾何級數(shù)模型擬合直線的斜率代表了群落的復(fù)雜性[7]同樣,分割線段模型擬合的曲線斜率在一定程度上反映了群落均勻度的變化趨勢[27]。用生態(tài)位模型更能直觀的體現(xiàn)群落的結(jié)構(gòu),所以本文選用生態(tài)位模型擬合研究地多度格局,確定分割線段模型為本文使用的主要模型。
2.1 鳥類多度格局月尺度分析
按鳥類物候,對2014年3月到2015年2月,一個生物年,共12個月數(shù)據(jù)進(jìn)行計算處理,得到以下結(jié)果。
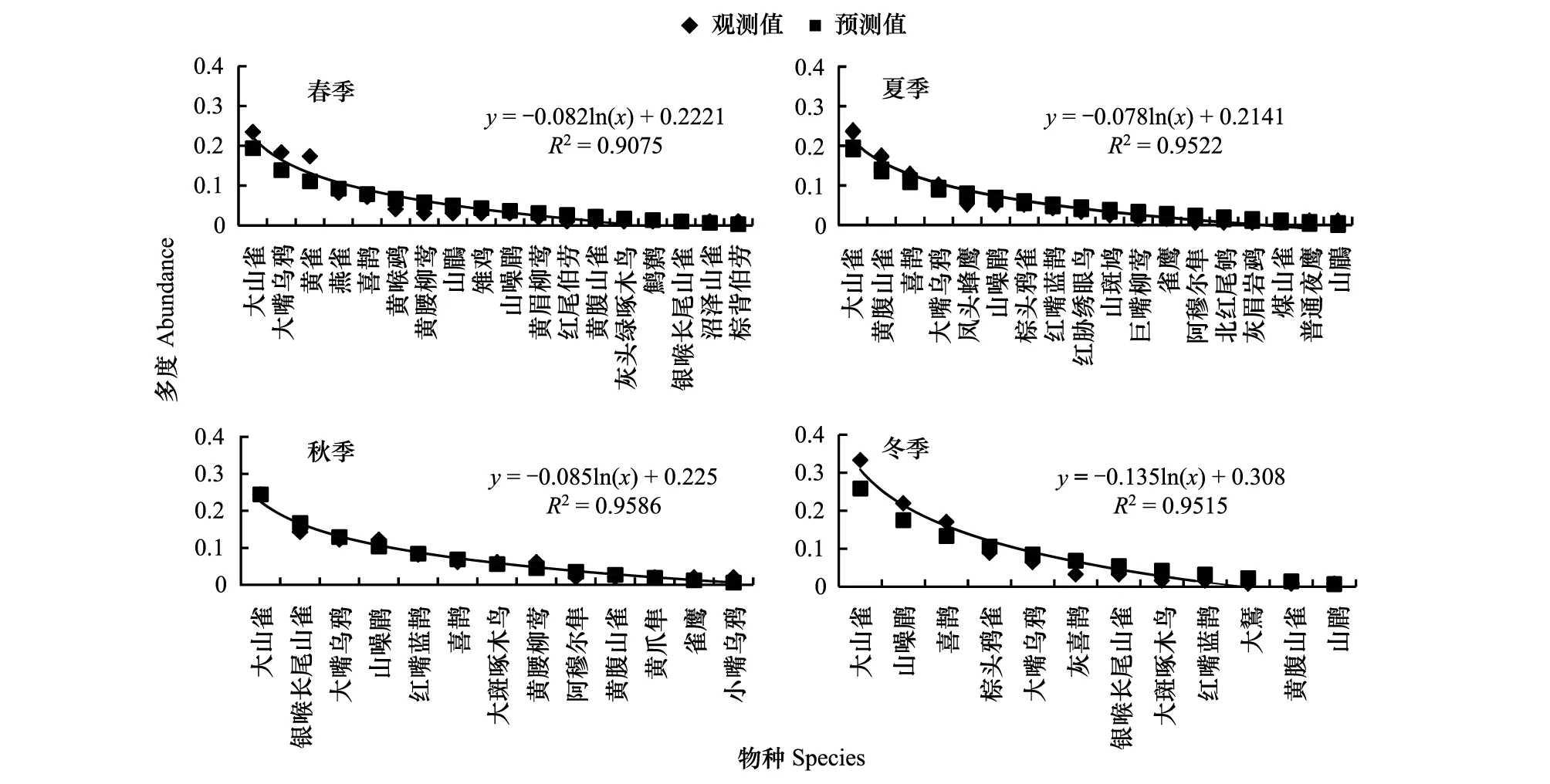
側(cè)柏針葉林模型擬合檢驗中,觀測值與預(yù)測值的相關(guān)系數(shù)r分別為0.982,0.866,0.918,0.982,0.975,0.909,0.960,0.979,0.892,0.974,0.976,0.860。T檢驗P值均小于0.01。觀測值與期望值屬于極顯著相關(guān)。說明側(cè)柏針葉林中,鳥類多度格局適用分割線段模型進(jìn)行擬合。由圖1可以看出,物種分布比較稀疏,相互之間競爭比較弱,即生態(tài)位分化程度較大,符合隨機生態(tài)位假說。該群落中,喜鵲(Picapica)、大山雀(Parusmajor)等相對多度最大,占有大部分生態(tài)位,優(yōu)勢度明顯,可做為側(cè)柏針葉林中的優(yōu)勢物種,其余各物種數(shù)量雖有差別,但不顯著。
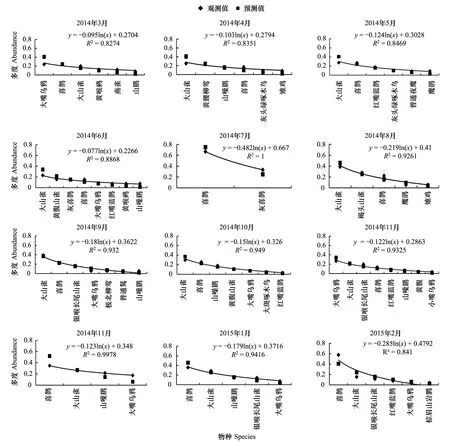
圖1 側(cè)柏針葉林中鳥類多度分割線段模型擬合Fig.1 Broke stick model fitting of bird abundance in coniferous forest
針闊混交林模型擬合檢驗中,相關(guān)系數(shù)r分別為0.756,0.921,0.978,0.941,0.980,0.942,0.969,0.990,0.930,0.945,0.945,0.945。T檢驗P值大部分小于0.01,3月、12月、1月、2月的P值大于0.05,相關(guān)性不顯著。這4個月處于早春或是冬季,鳥類種類、數(shù)量都小于其他月份。此外,從14年10月開始,所選調(diào)查點附近進(jìn)行了靜福寺遺跡挖掘工作,也對之后的鳥類調(diào)查帶來負(fù)面的影響。導(dǎo)致野外調(diào)查獲得數(shù)據(jù)量小,無法得出理想的擬合曲線。
由圖2可以看出:該群落中物種分布比較稀疏,物種的資源分割是隨機的,物種間競爭比較弱,符合隨機生態(tài)位假說。群落中,喜鵲、大山雀占有大部分生態(tài)位,優(yōu)勢度明顯,是群落中的優(yōu)勢物種,其余各物種數(shù)量雖有差別,但不顯著。并且如黃腹山雀(Parusvenustulus)、黃腰柳鶯(Phylloscopusproregulus)等小型林鳥的數(shù)量比重有所增加。
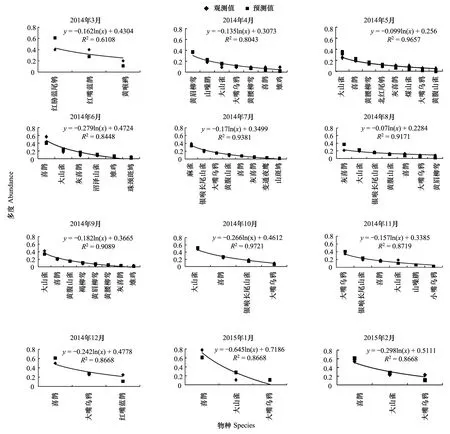
圖2 針闊混交林中鳥類多度分割線段模型擬合Fig.2 broke stick model fitting of bird abundance in conifer and broadleaf mixed forest
落葉闊葉林模型擬合檢驗中,相關(guān)系數(shù)r值分別為0.976,0.956,0.922,0.951,0.975,0.901,0.966,0.955,0.961,0.977,0.977,0.964.T檢驗P值均小于0.01。觀測值與期望值成極顯著相關(guān),說明該群落中資源的分割是隨機的,物種間競爭較弱,生態(tài)位分化較大,重疊程度小,符合隨機生態(tài)位假說。由圖3可以看出:落葉闊葉林中鳥類豐富度大于前兩種森林群落。該群落中,以大山雀為代表的小型鳥類占有大部分生態(tài)位,可做為闊葉落葉林中的優(yōu)勢物種。喜鵲的相對多度小于前兩種森林群落。在冬季時,以山噪鹛(Garrulaxdavidi)為代表的越冬鳥類所占比重增加。其余各物種數(shù)量雖有差別,但不顯著,且有相近的多度,各物種間能根據(jù)需要取其所需資源和諧共存。
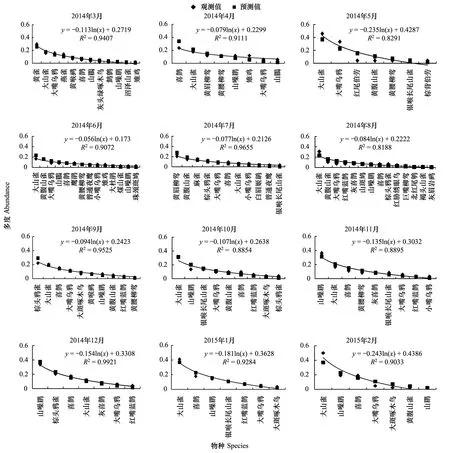
圖3 落葉闊葉林中鳥類多度分割線段模型擬合Fig.3 broke stick model fitting of bird abundance in broadleaved deciduous forest

圖4 3種林型擬合方程斜率月變化Fig.4 slopes of fitting equation in three forest types in every month
由圖1—圖3可以看出:在調(diào)查的鳥類群落中,落葉闊葉林物種豐富度最大,針葉林次之,針闊混交林最小。3種森林群落中大部分物種的相對多度值較小,僅少數(shù)幾個種的相對多度值較大,群落組成以少數(shù)種為主。
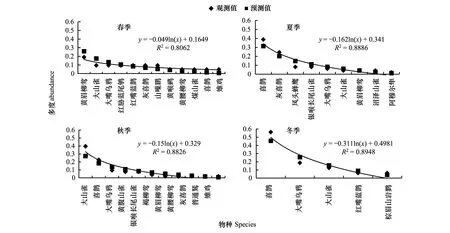
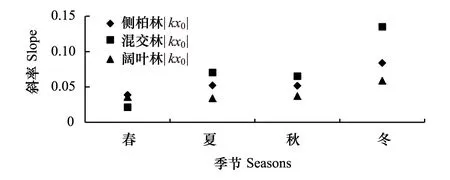
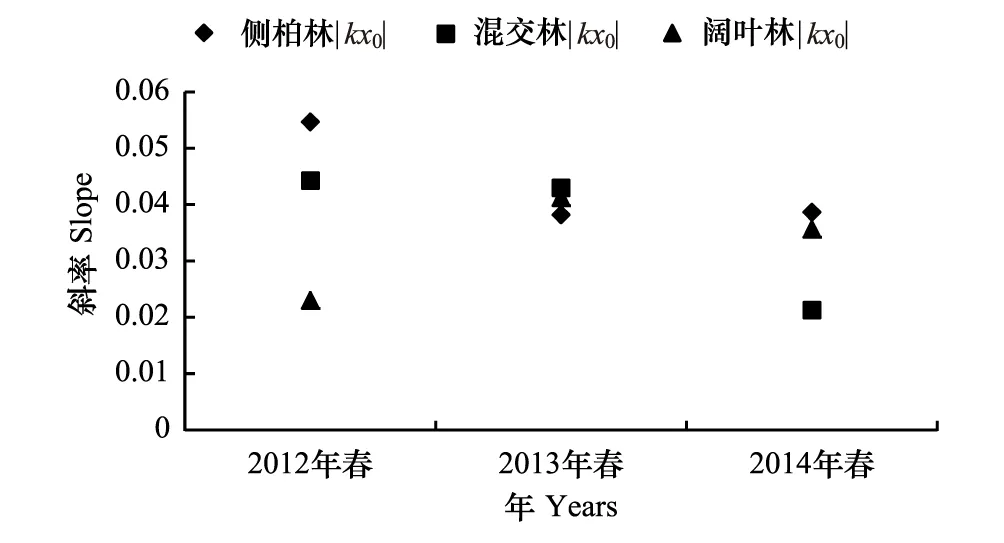
種-多度曲線中,斜率是圖像重要的參數(shù),它直觀的反應(yīng)圖像的變化速率。回歸方程曲線,大多數(shù)R>R0.02,屬于顯著相關(guān),只有混交林中3月、12月、1月、2月R 根據(jù)文獻(xiàn)[12-13]數(shù)據(jù),按本文方式分析12年鳥類群落多度格局,結(jié)果表明:單因素方差分析,12個月數(shù)據(jù)總體差異不顯著,11月與6月結(jié)果存在顯著差異,8月與5、6月結(jié)果差異顯著。按森林群落做單因素方差分析,3種森林群落間差異顯著,闊葉林與針葉林和針闊混交林差異顯著。 2.2 鳥類多度格局季尺度分析 季節(jié)劃分按鳥類春、秋遷徙期,繁殖期,越冬期劃分。 側(cè)柏針葉林四季模型擬合檢驗中,觀測值與預(yù)測值的相關(guān)系數(shù)r分別為0.902、0.950、0.980、0.917。T檢驗P值均小于0.01。觀測值與期望值屬于極顯著相關(guān)。說明季尺度上,側(cè)柏針葉林中,鳥類多度格局適用分割線段模型進(jìn)行擬合,群落中物種間競爭較弱,生態(tài)位上分布相對均勻,符合隨機生態(tài)位假說。由圖5可以看出,該群落中,喜鵲、大山雀、大嘴烏鴉(Corvusmacrorhynchos)相對多度最大,占有大部分生態(tài)位,優(yōu)勢度明顯,可作為側(cè)柏針葉林中的優(yōu)勢物種,四季各林型中優(yōu)勢種變化不大,其余各物種數(shù)量雖有差別,但不顯著,物種分布較為均勻。 圖5 側(cè)柏針葉林中鳥類四季多度分割線段模型擬合Fig.5 Broke stick model fitting of bird abundance in coniferous forest in four seasons 針闊混交林四季模型擬合檢驗中,觀測值與預(yù)測值的相關(guān)系數(shù)r分別為0.908、0.958、0.958、0.962。T檢驗P值均小于0.01。觀測值與期望值顯著相關(guān),符合隨機生態(tài)位假說。由圖6可以看出:春季時,該群落中,黃眉柳鶯(Phylloscopusinornatus)、大山雀相對多度最大,占有大部分生態(tài)位,優(yōu)勢度明顯,可作為優(yōu)勢種;夏季優(yōu)勢種為喜鵲、灰喜鵲(Cyanopicacyana);秋季優(yōu)勢種為大山雀、喜鵲;冬季優(yōu)勢種為喜鵲、大嘴烏鴉。因鳥類居留型不同,各季節(jié)鳥類群落優(yōu)勢種也存在差異。 圖6 針闊混交林中鳥類四季多度分割線段模型擬合Fig.6 Broke stick model fitting of bird abundance in conifer and broadleaf mixed forest in four seasons 落葉闊葉林四季模型擬合檢驗中,觀測值與預(yù)測值的相關(guān)系數(shù)r分別為0.959、0.984、0.984、0.983。T檢驗P值均小于0.01。觀測值與期望值顯著相關(guān),群落中物種分布符合隨機生態(tài)位假說。由圖7可以看出,該群落中,春季時,大山雀、大嘴烏鴉相對多度最大,占有大部分生態(tài)位,優(yōu)勢度明顯,可作為該群落的優(yōu)勢種;夏季優(yōu)勢種為大山雀、黃腹山雀;秋季優(yōu)勢種為大山雀、銀喉長尾山雀(Aegithaloscaudatus);冬季優(yōu)勢種為大山雀、山噪鹛。 圖7 闊葉落葉林中鳥類四季多度分割線段模型擬合Fig.7 broke stick model fitting of bird abundance in broadleaved deciduous forest in four seasons 回歸曲線,R>R0.02,屬于顯著相關(guān),且F檢驗的P值均小于0.01。以k×x0為縱軸,季節(jié)為橫軸得到圖8。由圖8可知,針闊混交林鳥類群落多度格局的季變化最大,其次是側(cè)柏林,落葉闊葉林變化最小。對季變化圖像斜率進(jìn)行雙變量方差分析。 以季節(jié)為固定因子,森林群落類型為隨機因子,主體間效應(yīng)檢驗,總體差異不顯著,交互作用均不顯著。多重比較分析,冬季與春、秋兩季差異顯著;冬季與夏季差異不顯著;春、夏、秋三季間差異不顯著。 以森林群落類型為固定因子,季節(jié)為隨機因子,主體間效應(yīng)檢驗,總體差異顯著,交互作用不顯著。落葉闊葉林與針闊混交林差異顯著;針葉林與落葉闊葉林、針闊混交林差異均不顯著。 圖8 3種林型擬合方程斜率季變化 Fig.8 Slopes of fitting equation in three forest types in four seasons 根據(jù)文獻(xiàn)[12-13]數(shù)據(jù),按本文方式分析12a鳥類群落多度格局,結(jié)果表明:以季節(jié)為固定因子,森林群落為隨機因子,主體間效應(yīng)檢驗,總體差異顯著,交互作用不顯著,季節(jié)差異不顯著;以森林群落類型為固定因子,季節(jié)為隨機因子,主體間效應(yīng)檢驗,總體差異極顯著,森林群落間差異極顯著,闊葉林與其他森林群落有顯著差異,針葉林與混交林差異不顯著。 2.3 鳥類多度格局年尺度分析 查看較早的實驗調(diào)查數(shù)據(jù)(2012年、2013年調(diào)查數(shù)據(jù))整理出兩年完整的春季鳥類調(diào)查數(shù)據(jù),利用2012、2013年春季數(shù)據(jù),重復(fù)上述工作,得到圖9。 2012、2013年春季模型擬合檢驗中,觀測值與預(yù)測值的相關(guān)系數(shù)r分別為0.952、0.977、0.891、0.963、0.933、0.896。T檢驗P值均小于0.01。觀測值與期望值顯著相關(guān),群落物種分布符合隨機生態(tài)位假說。兩年數(shù)據(jù)相比,群落物種組成沒有明顯變化,多度最大的優(yōu)勢種有所變化。 圖9 3種森林群落鳥類2012年和2013年春季多度分割線段模型擬合Fig.9 broke stick model fitting of bird abundance in three forest types in spring of 2012 and 2013 回歸曲線,R>R0.001,屬于高度相關(guān),且F檢驗的P值均小于0.01。以k×x0為縱軸,年為橫軸得到圖10。由圖10可知,3種森林群落中,群落多度格局均有明顯的年變化。斜率做單因素方差分析的到P>0.05差異性不顯著,說明3種森林群落年變化不明顯。 圖10 3種森林群落春季擬合方程斜率年變化 Fig.10 slopes of fitting equation in three forest types in spring of the years of 2012,2013,2014 3.1 結(jié)論 分割線段模型可以較好的擬合該研究區(qū)域鳥類群落的數(shù)量關(guān)系,并從4個季節(jié)、3種森林群落兩個角度對西山靜福寺地區(qū)鳥類群落多度格局進(jìn)行分析闡釋,說明鳥類群落的多度格局具有時間與空間尺度的雙重屬性。通過擬合的曲線,可以直觀地了解鳥類群落的組成、豐富度、鳥類在林中分布的狀況、群落中鳥類四季遷飛的變化。 3.1.1 時間尺度上鳥類群落多度格局 分別從月、季、年3種不同的時間尺度上對西山靜福寺地區(qū)鳥類群落多度格局進(jìn)行分析:月尺度上,針闊混交林中鳥類多度格局變化明顯,側(cè)柏林與闊葉落葉林中多度格局有所變化,但不明顯,各林型中鳥類因物候期的變化,鳥類種類與種數(shù)也在變化;季尺度上,冬季格局與春、秋兩季差異顯著,春、夏、秋三季格局差異不顯著。3種林型多度格局差異不顯著,除針闊混交林春季優(yōu)勢種為黃腰柳鶯,其他時期優(yōu)勢種均是本地留鳥(大山雀、喜鵲、灰喜鵲);年尺度上,3年春季多度格局相比較,無明顯變化。 3.1.2 空間尺度上鳥類群落多度格局 對西山靜福寺地區(qū)3種森林群落中鳥類群落多度格局的比較,可以看出:闊葉落葉林中,鳥類的豐富度最高,針闊混交林次之,針葉林與混交林相似;群落中大山雀、喜鵲、灰喜鵲多度處于最多地位,出現(xiàn)頻度最高,視為優(yōu)勢物種;其余鳥類多度相差不大,比較接近,物種分布較為均勻。落葉闊葉林中鳥類群落多度格局與針葉林、針闊混交林存在顯著差異,針葉林與針闊混交林中鳥類多度格局無顯著差異。 3.2 討論 本文從多度角度對西山靜福寺地區(qū)鳥類多度格局進(jìn)行研究,補充并豐富了該地區(qū)鳥類群落的研究。與種間聯(lián)結(jié)相似[12],該地區(qū)鳥類群落多度格局同樣具有時間和空間雙重屬性。 分割線段模型BSM是描述群落中缺乏相對重要性極強的物種,群落中物種并不豐富,但在生態(tài)位上的分布相對均勻,此分布符合隨機生態(tài)位假說[24],即群落中物種的資源分割是隨機的,物種間的生態(tài)位不重疊這樣一種情況的數(shù)學(xué)模型,該模型廣泛應(yīng)用于種群多度格局的研究。馬克平等用分割線段模型擬合北京東靈山地區(qū)9個森林群落種-多度數(shù)據(jù),效果不佳[28],而張金屯擬合美國紐約州闊葉林種-多度格局則效果很好,認(rèn)為此模型既適合于簡單群落,又適合于復(fù)雜群落結(jié)構(gòu)的研究[29],吳承禎等也用它擬合了觀光木群落種-多度數(shù)據(jù)[17],趙洪峰認(rèn)為該模型通常應(yīng)用于某些種群穩(wěn)定、生活史長、均質(zhì)的小群落,如森林中營巢的鳥類群落[30]。在鳥類群落中,鳥類的種類和數(shù)量會隨著季節(jié)的不同、植被的地帶性變化而有所改變。通過BSM模型擬合,我們可以直觀地認(rèn)識一個鳥類群落的多度分布、豐富度、均勻度等基本屬性,能更清楚的認(rèn)識一個鳥類群落的結(jié)構(gòu)人為干擾在一定程度上影響群落結(jié)構(gòu)及物種組成[31]。鳥類對人類活動的響應(yīng)主要體現(xiàn)在驚飛距離的差異,林地生境的鳥類的驚飛距離更小,人類活動對林地鳥類的影響尤為明顯[32]。在野外調(diào)查的過程中,2014年10月開始西山靜福寺地區(qū)進(jìn)行一次為期兩個月的文物保護(hù)挖掘工作,位置位于調(diào)查區(qū)域的針闊混交林地帶。挖掘工作破壞了該區(qū)域的原有景觀。而對于一個生境來說,其景觀結(jié)構(gòu)對其生物多樣性有至關(guān)重要的影響[33]。挖掘工作影響了鳥類數(shù)據(jù)的調(diào)查,導(dǎo)致數(shù)據(jù)的缺失或不完整,在擬合針闊混交林10月以后的模型時,出現(xiàn)擬合不顯著的情況。但David[34]在森林深處、森林小路沿線和野餐區(qū)的鳥類調(diào)查研究后發(fā)現(xiàn),人類的休閑活動可能會降低巢性鳥類的適宜度,但對大多數(shù)鳥類的棲息地沒有不利影響。對于人為活動是否會對鳥類群落多度格局造成影響,還有待研究。 [1] 林金安. 植物科學(xué)綜論. 哈爾濱: 東北林業(yè)大學(xué)出版社,1993:222- 223. [2] 中國科學(xué)院生物多樣性委員會. 生物多樣性研究的原理與方法. 北京: 中國科學(xué)技術(shù)出版社,1994:141- 165. [3] 馬克明. 物種多度格局研究進(jìn)展. 植物生態(tài)學(xué)報, 2003,27 (3):412- 526. [4] Pielou E.C. Ecological Diversity. New York: Wiley, 1975 [5] 張金屯. 群落中物種多度格局的研究綜述. 農(nóng)村生態(tài)環(huán)境, 1997, 13(4): 48- 54. [6] 彭少麟, 殷祚云, 任海, 郭勤峰. 多物種集合的種-多度關(guān)系模型研究進(jìn)展. 生態(tài)學(xué)報, 2003, 23(8): 1590- 1605. [7] Tokeshi M. Species abundance patterns and community structure. Advances in Ecological Research, 1993, 24: 111- 186. [8] Tokeshi M. Niche apportionment or random assortment: species abundance patterns revisited. Journal of Animal Ecology, 1990, 59(3): 1129- 1146. [9] Hoagstrom C W. The unified neutral theory of biodiversity and biogeography: Stephen P. Hubbell. Annals of the Entomological Society of America, 2005, 98(2): 241- 242. [10] 周淑榮, 張大勇. 群落生態(tài)學(xué)的中性理論. 植物生態(tài)學(xué)報, 2006, 30(5): 868- 877. [11] 廖成章, 徐永興, 柳江, 閆淑君, 畢曉麗. 福建將石自然保護(hù)區(qū)鳥類物種相對多度模型的擬合研究. 中國生態(tài)農(nóng)業(yè)學(xué)報, 2004, 12(2): 36- 39. [12] 董大穎, 范宗驥, 李扎西姐, 陳卓琳, 王敏增, 王奇峰, 李凱, 關(guān)文彬. 北京西山地區(qū)大山雀與其它鳥類種群種間聯(lián)結(jié)分析. 生態(tài)學(xué)報, 2013, 33(20): 6614- 6633. [13] 范宗驥, 董大穎, 鄭然, 王敏增, 王奇峰, 關(guān)文彬. 北京靜福寺側(cè)柏古樹林鳥類群落多樣性研究. 北京林業(yè)大學(xué)學(xué)報, 2013, 35(5): 46- 55. [14] Whittaker RH. Community and Ecosystem. 2nd ed. New York: Macmillan. 1975: 135- 168. [15] Whittaker R H. Evolution of species diversity in land communities // Hecht M K, Steere B W N C, eds. Evolutionary Biology. New York: Plenum Press, 1977, 10: 1- 67. [16] Molles Jr M C. Ecology: Concepts and Applications. Dubuque, IA: WCB/McGraw-Hill, 1999. [17] 吳承禎, 洪偉, 鄭群瑞. 福建萬木林保護(hù)區(qū)觀光木群落物種相對多度模型的擬合研究. 熱帶亞熱帶植物學(xué)報, 2001, 9(3): 235- 242. [18] González-Estrada E, Fawcett R H, Herrero M. Integrating models of relative abundance of species with the dry-weight-rank method for the botanical analysis of forest understorey vegetation. Grass and Forage Science, 2002, 57(2): 171- 183. [19] May R M. Theoretical Ecology. Oxford: Blackwell Scientific Pub., 1976. [20] Preston F W. The commonness, and rarity, of species. Ecology, 1948, 29(3): 254- 283. [21] Fisher R A, Corbet A S, Williams C B. The relation between the number of species and the number of individuals in a random sample of an animal population. Journal of Animal Ecology, 1943, 12(1): 42- 58. [22] Whittaker R H. Dominance and diversity in land plant communities. Science, 1965, 147(3655): 250- 260. [23] Whittaker R H. Evolution and measurement of species diversity. Taxon, 1972, 21(2/3): 213- 251. [24] MacArthur R H. On the relative abundance of bird species. Proceedings of the National Academy of Sciences of the United States of America 1957, 43(3): 293- 295. [25] Legendre P, Legendre L. Numerical Ecology. 2nd ed. Amsterdam: Elsevier Science, 1998: 194- 198. [26] 杜樂山, 楊洪曉, 郭曉蕾, 董大穎, 關(guān)文彬. 黃連木群落種間聯(lián)結(jié)指數(shù)—等級格局模型研究. 北京林業(yè)大學(xué)學(xué)報, 2013, 35(5): 37- 55. [27] 郭逍宇, 張金屯, 宮輝力, 張桂蓮, 董志. 安太堡露天礦區(qū)人工植被的物種多度分布分析. 林業(yè)科學(xué), 2007, 43(3): 118- 121. [28] 馬克平, 劉燦然, 于順利, 王巍. 北京東靈山地區(qū)植物群落多樣性的研究. Ⅲ. 幾種類型森林群落的種-多度關(guān)系研究. 生態(tài)學(xué)報, 1997, 17(6): 573- 583. [29] 張金屯. 美國紐約州闊葉林物種多度格局的研究(英文). 植物生態(tài)學(xué)報, 1999, 23(6): 481- 589. [30] 趙洪峰, 雷富民. 岷江上游典型退化生態(tài)系統(tǒng)鳥類物種多樣性的初步研究. 應(yīng)用與環(huán)境生物學(xué)報, 2003, 9(1): 42- 58. [31] 付裕. 人為活動對泰山森林植物群落物種多樣性的影響研究[D]. 泰安: 山東農(nóng)業(yè)大學(xué), 2009. [32] 陸祎瑋. 城市化對鳥類群落的影響及其鳥類適應(yīng)性的研究[D]. 上海: 華東師范大學(xué), 2007. [33] 李曉文, 胡遠(yuǎn)滿, 肖篤寧. 景觀生態(tài)學(xué)與生物多樣性保護(hù). 生態(tài)學(xué)報, 1999, 19(3): 399- 507. [34] Palomino D, Carrascal L M. Impact of recreation on forest bird communities: non-detrimental effects of trails and picnic areas. Acta Zoologica Sinica, 2007, 53(1): 54- 63. Bird abundance distribution patterns of Jing Fu Temple in Beijing Xishan CHEN Zhuolin,JIA Lili,GUAN Wenbin* BeijingForestryUniversity,Beijing100083,China The analysis of species abundance patterns is fundamental to understand community structure. In the present paper, the broken stick model, used to describe the relationship between species and abundance, was used to study the quantitative relationships of the bird community of the JingFu temple in Beijing Xishan. The model was tested using a correlation coefficient with a rank correlation analysis. The results showed that: (1) The bird community structure in this area was significantly correlated with the broken stick model. The model could be used to fit the pattern of bird community structure well in the region. (2) The pattern of bird abundance varied temporally and spatially. (3) The results showed that the abundance pattern in January was significantly different to the other months, and there were no significant differences in forest communities monthly. (4) The abundance pattern in winter was significantly different from those in spring and autumn, and between between deciduous broad-leaved and coniferous-broadleaved forest, and neither deciduous broad-leaved nor coniferous-broadleaved forest were significantly different from the coniferous forest seasonally. (5) There was no annual significant difference. bird community; abundance pattern; broke stick model 國家林業(yè)公益性行業(yè)科研專項經(jīng)費項目(200904019) 2015- 11- 04; 日期:2016- 08- 02 10.5846/stxb201511042234 *通訊作者Corresponding author.E-mail: swlab@bjfu.edu.cn 陳卓琳,賈麗麗,關(guān)文彬.北京西山靜福寺地區(qū)鳥類多度分布格局.生態(tài)學(xué)報,2017,37(6):1805- 1815. Chen Z L,Jia L L,Guan W B.Bird abundance distribution patterns of Jing Fu Temple in Beijing Xishan.Acta Ecologica Sinica,2017,37(6):1805- 1815.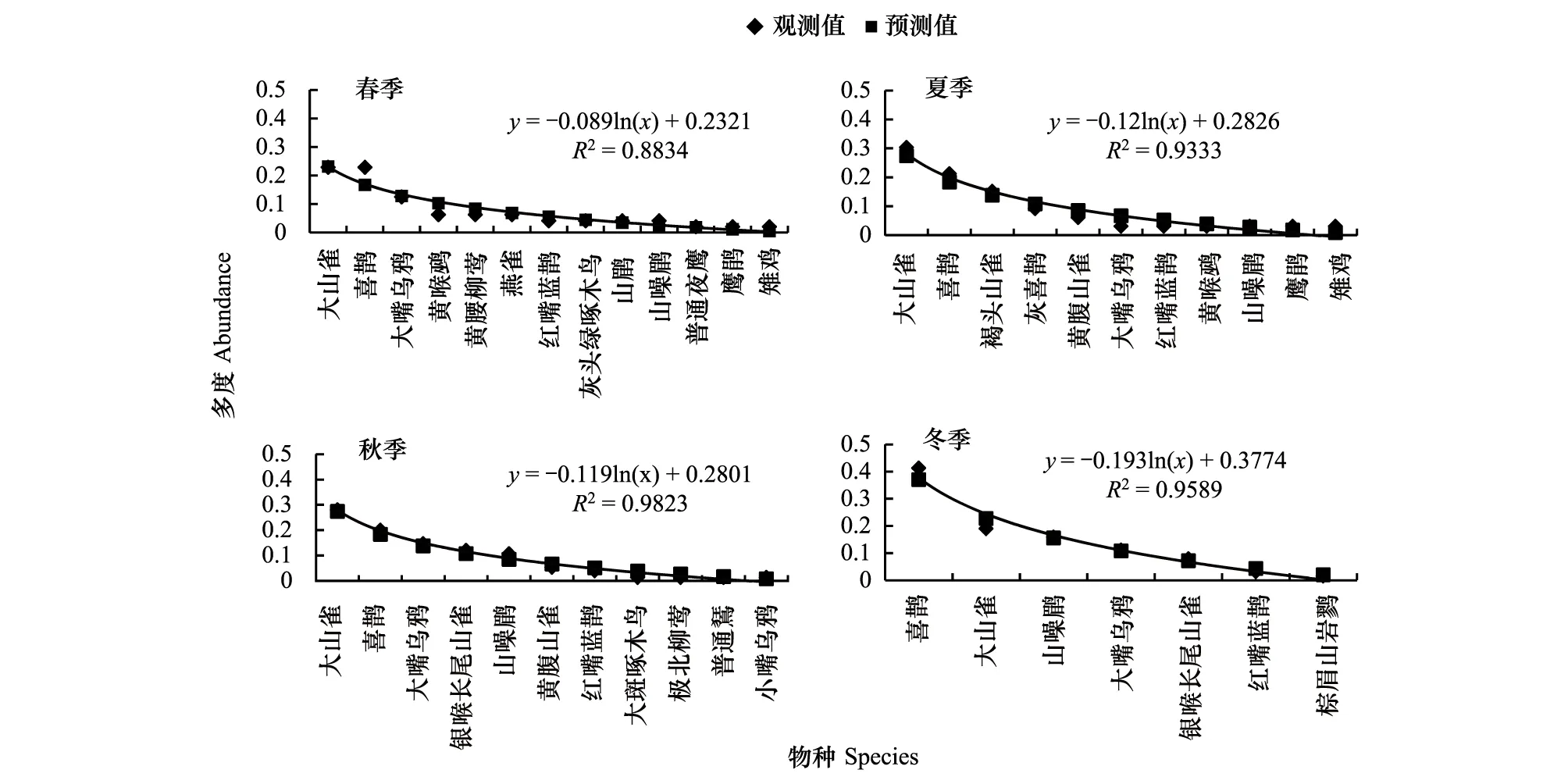

3 結(jié)論與討論
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
音樂探索(2022年2期)2022-05-30 21:01:37
保健醫(yī)苑(2021年7期)2021-08-13 08:48:02
學(xué)生天地(2020年36期)2020-06-09 03:12:30
小學(xué)科學(xué)(學(xué)生版)(2020年5期)2020-05-25 07:11:32
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
少兒科學(xué)周刊·少年版(2015年1期)2015-07-07 17:15:12
