MicroRNA-155 在肝細胞癌對索拉非尼抗藥中的作用研究
2017-05-17 03:45:44艾麥提牙森李德衛(wèi)
中國藥理學(xué)通報 2017年5期
呂 峰,王 偉,艾麥提·牙森,金 鑫,李德衛(wèi)
(重慶醫(yī)科大學(xué)附屬第一醫(yī)院肝膽外科,重慶 400016)
MicroRNA-155 在肝細胞癌對索拉非尼抗藥中的作用研究
呂 峰,王 偉,艾麥提·牙森,金 鑫,李德衛(wèi)
(重慶醫(yī)科大學(xué)附屬第一醫(yī)院肝膽外科,重慶 400016)
目的 探討微小RNA155(microRNA-155,miR-155)在肝細胞癌(hepatocellular carcinoma,HCC)對索拉非尼(sorafenib)抗藥中的作用。方法 將miR-155抑制慢病毒(miR-155 inhibitor)轉(zhuǎn)染miR-155表達相對較高的SMMC-7721細胞,而miR-155過表達慢病毒(miR-155)轉(zhuǎn)染miR-155表達相對較低的HepG2細胞;用熒光定量PCR(qPCR) 檢測經(jīng)慢病毒轉(zhuǎn)染的SMMC-7721細胞及HepG2細胞miR-155表達量,以驗證慢病毒轉(zhuǎn)染效果;通過CCK-8法及流式細胞術(shù)檢測各組細胞經(jīng)索拉非尼作用后的存活率及凋亡情況;用Western blot檢測凋亡相關(guān)蛋白活性caspase-3表達量,從而進一步探究各組細胞凋亡情況。結(jié)果 與對照組相比,轉(zhuǎn)染miR-155抑制慢病毒的SMMC-7721細胞表達miR-155明顯下調(diào)(P<0.01),經(jīng)索拉非尼處理后其存活率明顯降低(P<0.05),而索拉非尼誘導(dǎo)其凋亡明顯增加(P<0.01),其活性caspase-3表達量明顯上調(diào)(P<0.01);miR-155過表達慢病毒轉(zhuǎn)染HepG2細胞后,則相反。結(jié)論 miR-155 參與肝細胞癌對索拉非尼的抗藥,有希望成為一個肝癌治療的新靶標(biāo)。
微小RNA155; 肝細胞癌; 索拉非尼; 抗藥; 慢病毒; 活性 caspase-3
肝癌發(fā)病率及死亡率極高。目前,多靶點抗腫瘤新藥索拉非尼成為中晚期肝癌的重要治療措施[1]。然而,肝癌患者對索拉非尼的藥物抵抗問題也日趨暴露[2],研究表明多種 microRNAs 參與肝癌患者對化療藥物索拉非尼的抵抗。
MicroRNA是一類由內(nèi)源基因編碼的長度約為22個核苷酸的非編碼單鏈RNA分子,它們在機體參與靶基因抑制性轉(zhuǎn)錄后表達調(diào)控[3],許多microRNAs在腫瘤的發(fā)生、發(fā)展過程中有著重要作用[4]。miR-155是一個非常重要的microRNA,在多種腫瘤的發(fā)生、發(fā)展過程中均可見其蹤影,可促進腫瘤細胞的增殖、遷移、侵襲等,如乳腺癌[5-7]、膠質(zhì)瘤[8]、結(jié)直腸癌[9]、肝癌[10-12]等。目前研究證實miR-222[13]、miR-494[14]、miR-21[15]等參與肝癌患者對化療藥物索拉非尼的抵抗,但是miR-155是否參與肝癌患者對索拉非尼的抵抗仍然不清楚。本研究旨在通過慢病毒轉(zhuǎn)染調(diào)高或調(diào)低miR-155在肝癌細胞中的表達,來探究其是否參與索拉非尼抵抗。
1 材料與方法
1.1 材料 SMMC-7721及HepG2細胞均由重慶醫(yī)科大學(xué)附屬第一醫(yī)院實驗研究中心提供;索拉非尼(sorafenib)購自Selleckchem公司(20 mg/支),相對分子質(zhì)量為464.82,批次:S739702,純度:99.87%;RPMI 1640培養(yǎng)基購自Corning公司;胎牛血清購自PAN-BiotechGmbH公司;青霉素/鏈霉素雙抗、RIPA裂解液(強)、PMSF(100 nmol·L-1)、SDS-PAGE凝膠配制試劑盒、0.25%胰酶均購自碧云天公司;熒光定量PCR試劑及逆轉(zhuǎn)錄試劑盒均購自WesternBiotechnology公司;慢病毒購自吉凱公司;Cell Counting Kit-8(CCK-8)試劑盒購自日本株式會社同仁化學(xué)研究所;兔抗人caspase-3購自Abcam公司;辣根過氧化物酶標(biāo)記的羊抗兔IgG購自武漢三鷹生物技術(shù)有限公司。
1.2 細胞培養(yǎng)及穩(wěn)定細胞株的建立 SMMC-7721及HepG2細胞用含10%的胎牛血清、青霉素(1×105IU·L-1)和鏈霉素(100 g·L-1)的RPMI 1640培養(yǎng)基,于5% CO2、37 ℃ 孵箱培養(yǎng)。
將對數(shù)生長期的人肝癌細胞SMMC-7721、HepG2接種于6孔板中,每孔約1×105個細胞,待d 2細胞匯合度達50%~70%后,用miR-155 inhibitor及control慢病毒轉(zhuǎn)染SMMC-7721細胞;用miR-155及control慢病毒轉(zhuǎn)染HepG2細胞;MOI=10,感染24 h后換新鮮培養(yǎng)基,繼續(xù)培養(yǎng)24 h后用嘌呤霉素(2 mg·L-1)進行篩選,以建立穩(wěn)定細胞株。篩選完成后,用熒光顯微鏡觀察細胞GFP綠色熒光表達及其形態(tài)學(xué)的改變。
1.3 qPCR檢測各組細胞miR-155的表達 細胞總RNA的提取、反轉(zhuǎn)錄為cDNA及擴增步驟均按照熒光定量PCR試劑盒說明書操作。反轉(zhuǎn)錄條件為:25 ℃ 10 min,42 ℃ 60 min,85 ℃ 5 min。擴增條件為:94 ℃預(yù)變性4 min;94 ℃變性20 s,60 ℃退火30 s,72 ℃延伸30 s,循環(huán)35次;72 ℃檢測信號。以U6作為內(nèi)參計算miR-155相對表達量。
1.4 CCK-8法檢測細胞存活率 分別取對數(shù)生長期的SMMC-7721 miR-155 inhibitor、SMMC-7721 miR-Control、HepG2 miR-155、HepG2 miR-Control細胞,以1×103/孔的密度接種于96孔板,每孔培養(yǎng)基為100 μL,培養(yǎng)24 h后,等量更換為含有索拉非尼的新鮮培養(yǎng)基,參閱文獻[13-14]其終濃度為10 μmol·L-1。各組并設(shè)只具有細胞、CCK-8液、不含索拉非尼的常規(guī)培養(yǎng)基的不加藥組及只具有培養(yǎng)基及CCK-8液而沒有細胞的空白組,每組設(shè)6個復(fù)孔。更換培養(yǎng)基后,SMMC-7721 miR-155 inhibitor、SMMC-7721 miR-Control 兩組細胞培養(yǎng)48 h,HepG2 miR-155、HepG2 miR-Control兩組細胞培養(yǎng)24 h后,每孔加入CCK-8液10 μL,孵育4 h,用酶標(biāo)儀(波長450 nm)測定OD值,計算細胞存活率:存活率/%=(加藥組OD值-空白組OD值)/(不加藥組OD值-空白組OD值)×100%。
1.5 流式細胞儀檢測細胞凋亡 用預(yù)熱的不含EDTA胰酶消化收集經(jīng)索拉非尼處理后的各組細胞(索拉非尼終濃度及處理時間同上),用預(yù)冷PBS液洗滌細胞2次,1 000 r·min-1離心5 min,棄上清,用250 μL 1×Binding buffer重懸細胞,取100 μL細胞懸液于5 mL流式管中,再加入5 μL Annexin V-PE和10 μL 7-AAD,輕輕混勻,避光、室溫反應(yīng)15 min,加入400 μL 1×Binding buffer,混勻后盡快檢測。流式細胞儀檢測、分析細胞凋亡步驟參考說明書。
1.6 Western blot 檢測細胞中活性caspase-3的表達 細胞經(jīng)索拉非尼處理完成后(索拉非尼終濃度及處理時間同上)提取總蛋白;SDS-PAGE電泳,以60 V電壓分離上層膠,在溴酚藍進入下層膠時電壓變?yōu)?20 V繼續(xù)電泳, 待溴酚藍到達下層膠底端附近停止電泳;以250 mA恒定電流轉(zhuǎn)膜30 min;轉(zhuǎn)膜完畢后,漂洗1~2 min再加入脫脂奶粉,在搖床上緩慢搖動,室溫封閉2 h;以1 ∶1 000比例稀釋兔抗人caspase-3,4 ℃ 孵育過夜;孵育完成后回收一抗并加入TBST,在搖床上緩慢搖動漂洗3次,每次10 min;二抗為辣根過氧化物酶(HRP)標(biāo)記的羊抗兔IgG(1 ∶2 000),于37 ℃孵育2 h;TBST洗膜3次, 每次10 min,顯影并分析活性caspase-3的表達情況。
2 結(jié)果
2.1 穩(wěn)定細胞株的建立與細胞形態(tài)學(xué)改變 慢病毒轉(zhuǎn)染細胞并篩選完成后, 熒光顯微鏡下觀測到GFP綠色熒光明顯表達,每個細胞均高表達綠色熒光(Fig 1)。與SMMC-7721 miR-Control組相比,SMMC-7721 miR-155 inhibitor組細胞形態(tài)呈規(guī)則形或近圓形;與HepG2 miR-Control組相比,HepG2 miR-155組細胞形態(tài)呈長梭形、不規(guī)則。提示各種穩(wěn)定細胞株成功建立,miR-155在肝癌細胞中高表達能誘導(dǎo)細胞表型的改變,從而影響細胞的生物學(xué)活性。
2.2 各組細胞miR-155的表達 首先用qPCR法分別檢測SMMC-7721細胞系及HepG2細胞系miR-155的表達,發(fā)現(xiàn)SMMC-7721細胞miR-155表達量較HepG2細胞高(P<0.01)(Fig 2A)。因此,選用miR-155抑制慢病毒轉(zhuǎn)染SMMC-7721細胞,miR-155過表達慢病毒轉(zhuǎn)染HepG2細胞。轉(zhuǎn)染完成后,再用qPCR法進行驗證,檢測結(jié)果顯示(Fig 2B):SMMC-7721 miR-155 inhibitor組細胞中miR-155表達量較SMMC-7721 miR-Control組明顯減少(P<0.01);HepG2 miR-155組細胞中miR-155表達量較HepG2 miR-Control組明顯增多(P<0.01)。證實各組細胞成功轉(zhuǎn)染慢病毒,且各組細胞miR-155表達量均得到相應(yīng)表達,其差異有統(tǒng)計學(xué)意義,提示可進行后續(xù)實驗。
2.3 各組細胞存活率 用CCK-8法檢測經(jīng)索拉非尼作用后的各組細胞存活率。檢測結(jié)果顯示(Fig 3),SMMC-7721 miR-155 inhibitor組細胞存活率(56.54±4.98)%較SMMC-7721 miR-Control組(77.70±10.34)%明顯降低(P<0.05);HepG2 miR-155組細胞存活率(63.46±7.85)%較HepG2 miR-Control組(48.21±4.87)%明顯升高(P<0.05)。可見調(diào)低肝癌細胞miR-155的表達可降低其經(jīng)索拉非尼作用后的存活率,相反,調(diào)高肝癌細胞miR-155的表達可增加其經(jīng)索拉非尼作用后的存活率。以上結(jié)果提示,miR-155可能在肝細胞癌對索拉非尼抵抗中起作用。
2.4 各組細胞凋亡率 為進一步驗證以上結(jié)論,采用流式細胞術(shù)檢測細胞凋亡率。結(jié)果顯示(Fig 4),經(jīng)索拉非尼處理各組細胞后,SMMC-7721 miR-155 inhibitor組細胞凋亡率(35.00±2.65)%較SMMC-7721 miR-Control組(12.33±1.53)%明顯升高(P<0.01);HepG2 miR-155組細胞凋亡率(9.67±1.53)%較HepG2 miR-Control組(19.00±1.73)%明顯降低(P<0.01)。可見肝癌細胞miR-155的表達量下調(diào)后,其經(jīng)索拉非尼作用后的凋亡率明顯上升,而肝癌細胞miR-155的表達量上調(diào)后,其經(jīng)索拉非尼作用后的凋亡率則明顯下降。此結(jié)果進一步提示miR-155可能參與肝細胞癌對索拉非尼抗藥作用。
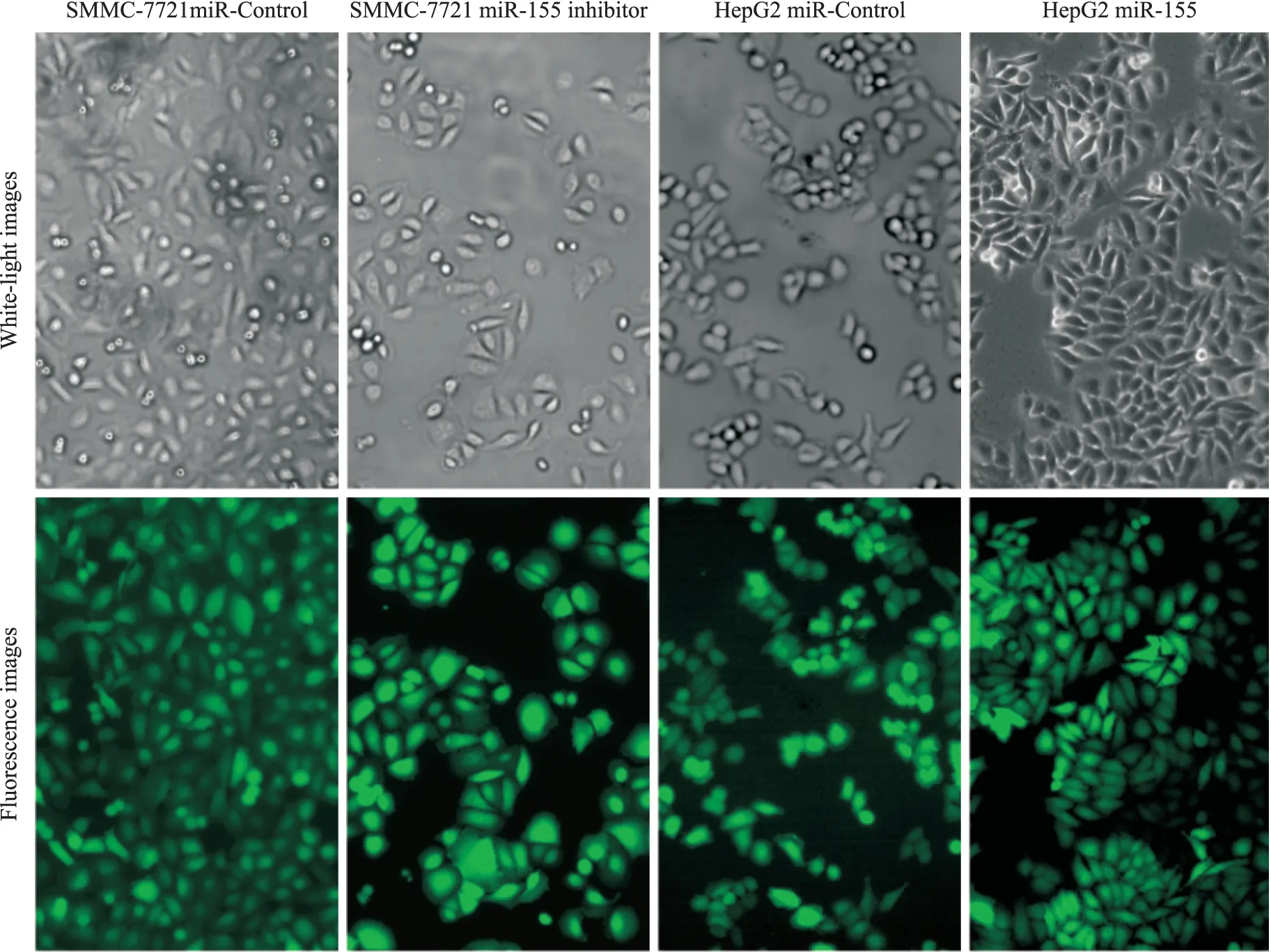
Fig 1 Fluorescent microscopy of infection efficiency of lentivirus in SMMC-7721 and HepG2 cells and cell morphologies(×100)
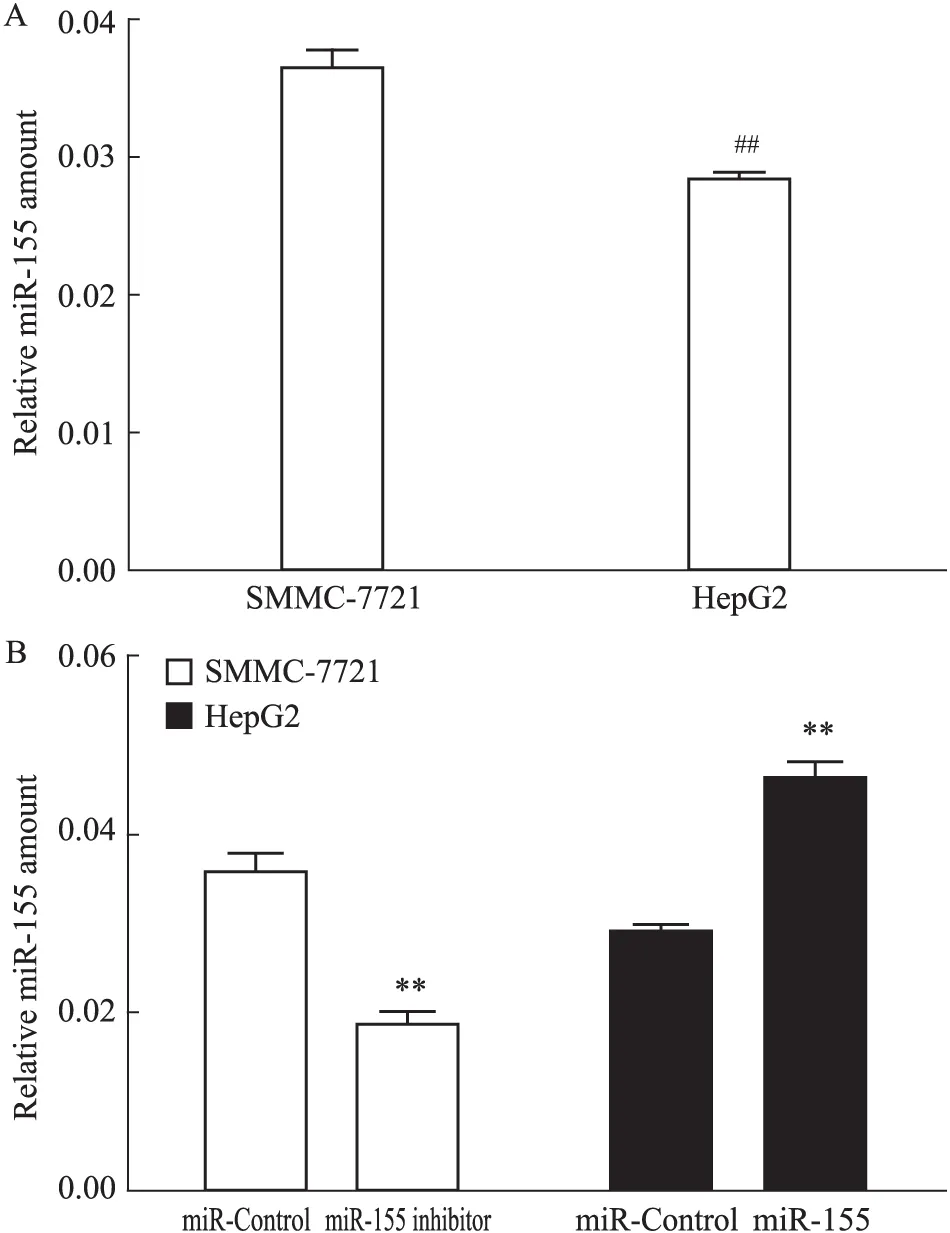
Fig 2 Expression of miR-155 in hepatocellular carcinoma cells
##P<0.01vsSMMC-7721;**P<0.01vsmiR-Control
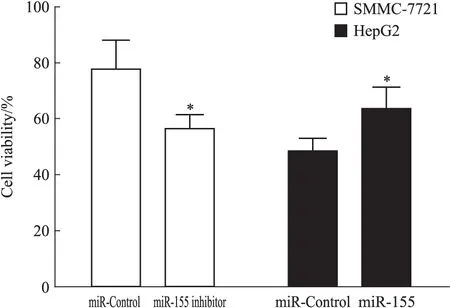
Fig 3 Viability of hepatocellular carcinoma cells
*P<0.05vsmiR-Control
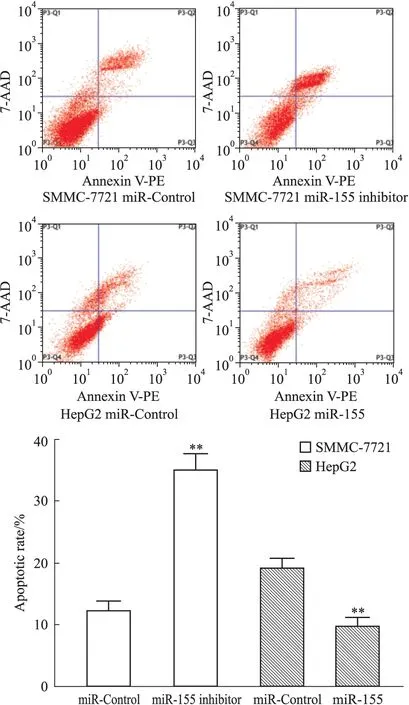
Fig 4 Influence of miR-155 on apoptosis of hepatocellular carcinoma cells induced by sorafenib
**P<0.01vsmiR-Control
2.5 各組細胞中活性caspase-3的表達量 Caspase-3是細胞凋亡過程中最主要的終末剪切酶,caspase-3在細胞凋亡中起著不可替代的作用,能很好反映細胞的凋亡情況。在凋亡過程中caspase-3可以被多種因素活化,從而誘導(dǎo)細胞的凋亡。Western blot檢測結(jié)果顯示(Fig 5),與對照組相比,SMMC-7721 miR-155 inhibitor組細胞活性caspase-3表達量明顯較高(P<0.01),HepG2 miR-155組細胞活性caspase-3表達量明顯較低(P<0.01)。結(jié)果表明miR-155低表達能促進索拉非尼誘導(dǎo)的肝癌細胞凋亡,而高表達能抑制索拉非尼誘導(dǎo)的肝癌細胞凋亡,更進一步說明miR-155可能參與肝細胞癌對索拉非尼抗藥作用。

Fig 5 Relative protein expression of caspase-3 in each group
**P<0.01vsmiR-Control
3 討論
肝細胞肝癌對靶向藥物索拉非尼的抵抗問題日趨暴露,多種microRNAs參與肝癌患者對化療藥物索拉非尼的抵抗,但是miR-155是否參與肝癌患者對索拉非尼的抵抗仍然不清楚。研究發(fā)現(xiàn),肝癌組織中miR-155的表達與肝癌細胞的低分化、高轉(zhuǎn)移、TNM分期分級、一年無復(fù)發(fā)生存率明顯相關(guān)[16]。一些研究證實miR-155可靶向抑制SOX6,從而間接下調(diào)p21waf1/cip1的表達,促進肝癌細胞的增殖[17]。miR-155異常表達能促進肝移植后肝癌患者的癌細胞侵襲,導(dǎo)致患者預(yù)后不良[11]。miR-155可靶向抑制TP53INP1誘導(dǎo)肝癌干細胞形成及增殖[18]。上述研究表明, miR-155 的異常表達可促進肝癌細胞惡性表型的轉(zhuǎn)變,是一個致癌microRNA,我們推測其可能參與肝細胞肝癌對索拉非尼的藥物抵抗。 本研究以SMMC-7721細胞系及HepG2細胞系為肝細胞肝癌的細胞模型,發(fā)現(xiàn)SMMC-7721細胞系相對高表達miR-155,而HepG2細胞系則相對低表達miR-155,所以選擇抑制性慢病毒轉(zhuǎn)染SMMC-7721細胞系,而過表達慢病毒轉(zhuǎn)染HepG2細胞系。發(fā)現(xiàn)miR-155抑制慢病毒轉(zhuǎn)染的SMMC-7721細胞系表達miR-155與對照組相比明顯下調(diào),經(jīng)索拉非尼處理后其存活率明顯低于對照組,而其凋亡率明顯高于對照組;而轉(zhuǎn)染過表達miR-155組的HepG2細胞表達miR-155較對照組明顯上調(diào),經(jīng)索拉非尼處理后其存活率明顯高于對照組,而其凋亡率明顯低于對照組。這表明miR-155增強了肝細胞癌對索拉非尼的抵抗作用,各組間活性caspase-3的表達差異也進一步證實了這一點。因此,miR-155有望成為一個肝癌診斷及治療的新靶標(biāo)。
miR-155通過何種機制增強肝細胞癌對索拉非尼的抵抗作用仍然不清楚。miR-155在乳腺癌抵抗化療藥物中也起著至關(guān)重要的作用,其機制包括對抑癌基因FOXO3a的抑制作用、絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)及上皮間質(zhì)轉(zhuǎn)化(EMT)信號通路的促進作用、對RhoA靶蛋白的抑制作用等[19]。許多其他microRNAs在肝細胞癌對索拉非尼的抵抗作用中也有參與,如miR-222通過激活PI3K/Akt信號通路,促進肝癌細胞的增殖、遷移、侵襲及對索拉非尼抵抗作用[13];miR-494通過靶向抑制PTEN,從而激活PI3K/Akt信號通路,促進肝癌細胞增殖、遷移、侵襲及對索拉非尼抵抗作用[14];miR-21通過PTEN/Akt通路抑制肝癌細胞的自噬作用,從而加強肝癌細胞對索拉非尼的抵抗作用[15]。可見microRNAs可通過作用多靶點、多種機制調(diào)節(jié)腫瘤對化療藥物的抵抗作用。而miR-155是否也通過激活PTEN/PI3K/Akt信號通路及其他信號通路,從而抑制、阻斷索拉非尼誘導(dǎo)的肝癌細胞凋亡,尚有待進一步研究。
綜上所述,miR-155的高表達可增強肝細胞癌對索拉非尼抗藥性,然而其機制需進一步研究。因而,miR-155作為一種潛在的靶向治療分子在肝細胞肝癌的生物治療中具有重要臨床意義。
(致謝:本實驗完成于重慶醫(yī)科大學(xué)附屬第一醫(yī)院實驗研究中心。本研究中許多實驗技術(shù)由實驗研究中心的湯為學(xué)老師、鄧曉娟老師指導(dǎo),在此深表感謝!)
[1] 王 芳,符立梧. 多靶點抗腫瘤新藥索拉非尼的研究進展[J].中國藥理學(xué)通報,2008,24(8):1117-20.
[1] Wang F, Fu L W. Research advances in sorafenib:amultiple targeted anti-tumor agent[J].ChinPharmacolBull,2008, 24(8):1117-20.
[2] 朱 云,程 旸,李愛民. 原發(fā)性肝細胞癌對索拉非尼耐藥機制的研究進展[J]. 中國藥理學(xué)通報,2013,29(6):752-5.
[2] Zhu Y, Cheng Y,Li A M. Mechanisms of drug resistance to sorafenib in hepatocellular carcinoma[J].ChinPharmacolBull,2013, 29(6): 752-5.
[3] Bartel D P. MicroRNAs: genomics, biogenesis, mechanism, and function[J].Cell,2004, 116(2): 281-97.
[4] Jansson M D, Lund A H. MicroRNA and cancer[J].MolOncol,2012, 6(6): 590-610.
[5] Iorio M V, Ferracin M, Liu C G, et al. MicroRNA gene expression deregulation in human breast cancer[J].CancerRes,2005, 65(16):7065-70.
[6] Kong W, Yang H, He L, et al. MicroRNA-155 is regulated by the transforming growth factor beta/Smad pathway and contributes to epithelial cell plasticity by targeting RhoA[J].MolCellBiol,2008, 28(22):6773-84.
[7] Kong W, He L, Coppola M, et al. microRNA-155 regulates cell survival, growth, and chemosensitivity by targeting FOXO3a in breast cancer[J].JBiolChem,2010, 285(23):17869-79.
[8] Ling N, Gu J, Lei Z, et al. MicroRNA-155 regulates cell proliferation and invasion by targeting FOXO3a in glioma[J].OncolRep,2013, 30(5):2111-8.
[9] Zhang G J, Xiao H X, Tian H P, et al. Upregulation of microRNA-155 promotes the migration and invasion of colorectal cancer cells through the regulation of claudin-1 expression[J].IntJMolMed,2013, 31(6):1375-80.
[10]Yan X L, Jia Y L, Chen L, et al. Hepatocellular carcinoma-associated mesenchymal stem cells promote hepatocarcinoma progression: role of the S100A4-miR155-SOCS1-MMP9 axis[J].Hepatology,2013, 57(6):2274-86.
[11]Han Z B, Chen H Y, Fan J W, et al. Up-regulation of microRNA-155 promotes cancer cell invasion and predicts poor survival of hepatocellular carcinoma following liver transplantation[J].JCancerResClinOncol,2012, 138(1):153-61.
[12]Liu F, Kong X, Lv L, et al. TGF-β1 acts through miR-155 to down-regulate TP53INP1 in promoting epithelial-mesenchymal transition and cancer stem cell phenotypes[J].CancerLett,2015,359(2):288-98.
[13]Liu K, Liu S, Zhang W, et al. miR-222 regulates sorafenib resistance and enhance tumorigenicity in hepatocellular carcinoma[J].IntJOncol, 2014, 45(4): 1537-46.
[14]Liu K, Liu S, Zhang W, et al. miR-494 promotes cell proliferation, migration and invasion, and increased sorafenib resistance in hepatocellular carcinoma by targeting PTEN[J].OncolRep,2015,34(2):1003-10.
[15]He C,Dong X,Zhai B,et al.MiR-21 mediates sorafenib resistance of hepatocellular carcinoma cells by inhibiting autophagy via the PTEN/Akt pathway[J].Oncotarget,2015,6(30):28867-81.
[16]Guan C, Yang F, He X, et al. Clinical significance of microRNA-155 expression in hepatocellular carcinoma[J].OncolLett,2016,11(2):1574-80.
[17]Xie Q, Chen X, Lu F, et al. Aberrant expression of microRNA 155 may accelerate cell proliferation by targeting sex-determining region Y box 6 in hepatocellular carcinoma[J].Cancer,2012,118(9):2431-42.
[18]Liu F, Kong X, Lv L, et al. MiR-155 targets TP53INP1 to regulate liver cancer stem cell acquisition and self-renewal[J].FEBSLett,2015,589(4):500-6.
[19]Yu D D, Lv M M, Chen W X, et al. Role of miR-155 in drug resistance of breast cancer[J].TumourBiol,2015,36(3):1395-401.
Effect of microRNA-155 on sorafenib resistance in hepatocellular carcinoma
LYU Feng, WANG Wei, AI Mai Ti·Ya Sen, JIN Xin, LI De-wei
(DeptofHepatobiliarySurgery,theFirstAffiliatedHospitalofChongqingMedicalUniversity,Chongqing400016,China)
Aim To investigate the effect of microRNA-155(miR-155) on sorafenib resistance in hepatocellular carcinoma(HCC).Methods Lentivirus mediated miR-155 inhibition was transfected into SMMC-7721 cells, while lentivirus mediated miR-155 overexpression was transfected into HepG2 cells. The level of miR-155 was evaluated by qPCR. Cell viability and apoptosis were analyzed by cell counting kit-8(CCK-8) assay and flow cytometry, respectively.The protein expression of activated caspase-3 was measured by Western blot.Results Compared to control group, the expression of miR-155 was significantly downregulated in miR-155 inhibition lentivirus infected SMMC-7721 cells(P<0.01), sorafenib treatment markedly suppressed cell viability(P<0.05) and increased cell apoptosis(P<0.01), as well as enhanced the expression of activated caspase-3(P<0.01). However, HepG2 cells were infected by miR-155 overexpression lentivirus which deserved completely opposite results.Conclusion miR-155 may participate in sorafenib resistance in HCC and provide a promising molecular target for the treatment of HCC.
mircoRNA-155; hepatocellular carcinoma; sorafenib; drug resistance; lentivirus; activated caspase-3
2016-12-15,
2017-01-10
國家自然科學(xué)基金資助項目(No 81470898)
呂 峰(1990-),男,碩士生,研究方向:miR-155 在肝細胞癌中的生物學(xué)作用及其機制,E-mail:1036688945@qq.com; 李德衛(wèi)(1970-),男,博士,教授,研究方向:肝膽疾病,通訊作者,E-mail: lidewei406@sina.com
時間:2017-4-24 11:20
http://kns.cnki.net/kcms/detail/34.1086.R.20170424.1120.028.html
10.3969/j.issn.1001-1978.2017.05.014
A
1001-1978(2017)05-0657-06
R329.25;R342.2;R373.9;R394.2;R735.702.2;R735.705
猜你喜歡
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
天津醫(yī)科大學(xué)學(xué)報(2021年1期)2021-01-26 00:57:10
天津醫(yī)科大學(xué)學(xué)報(2019年3期)2019-08-13 06:53:08
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
中國繼續(xù)醫(yī)學(xué)教育(2015年3期)2016-01-06 01:36:44
分子影像學(xué)雜志(2015年3期)2015-12-04 03:28:58
腫瘤預(yù)防與治療(2015年1期)2015-09-26 07:26:20
中國當(dāng)代醫(yī)藥(2015年16期)2015-03-01 02:03:11