閩東秋茄(Kandelia candel)人工林與天然林異齡葉性狀比較
2017-05-30 10:48:04潘云龍朱清波陳欣凡李彬歐陽玉瑩吳承禎洪滔
熱帶作物學報 2017年4期
潘云龍 朱清波 陳欣凡 李彬 歐陽玉瑩 吳承禎 洪滔


摘 要 比較閩東不同生境秋茄[Kandelia candel (Linn.) Druce]人工林與天然林異齡葉性狀間的相互關系,結果顯示:(1)2種林分的秋茄林葉MLA、SLA和LDMC,當年生均大于往年生,分別為MLA(38.4 cm2>36.0 cm2)、SLA(78.9 m2/kg>67.5 m2/kg)、LDMC(258.7 mg/g>193.8 mg/g);(2)當年生和往年生葉MLA人工林均大于天然林,分別為當年生(40.7 cm2>36.1 cm2)、往年生(37.5 cm2>34.4 cm2)。當年生葉SLA人工林小于天然林,為(78.4 m2/kg<79.4 m2/kg),往年生葉SLA人工林大于天然林,為(71.0 m2/kg>64.0 m2/kg)。當年生與往年生葉LDMC人工林均大于天然林,分別為當年生(344.4 mg/g>173.0 mg/g)、往年生(225.6 mg/g>162.0 mg/g)。研究表明,異齡葉對光資源獲取和儲存物質以構建葉器官這2種投資策略的權衡,是植物在各自生境中適應性進化的結果。秋茄能對生境因子脅迫調整生活策略應對,具有較強的環境適應性,適宜作為海岸防護林樹種人工栽植。
關鍵詞 秋茄;異齡葉;葉性狀
中圖分類號 S718.3 文獻標識碼 A
Abstract In the paper, the experiments were conducted to compare the foliar traits of uneven-aged leaves of Kandelia candel plantations and natural forests in the eastern regions of Fujian Province. The results showed that: (1)The leaf MLA, SLA and LDMC of the two kinds of K. candel forests were all higher in current-year than those in former-year leaves, respectively(38.4 cm2>36.0 cm2), (78.9 m2/kg>67.5 m2/kg), (258.7 mg/g>193.8 mg/g).(2)The MLA in both current-and former-year leaves of the plantation were significantly higher than those of the natural forest, respectively(40.7 cm2>36.1 cm2), (37.5 cm2>34.4 cm2). The leaf SLA of the plantation was higher in current-year leaves of the natural forest(78.4 m2/kg<79.4 m2/kg), while former-year leaves showed the reverse trend (71.0 m2/kg>64.0 m2/kg). The leaf LDMC was greater in the plantation than the natural forest in current-year and former-year leaves, respectively(344.4 mg/g>173.0 mg/g), (225.6 mg/g>162.0 mg/g). The study showed it was the result of adaptive evolution for uneven-aged leaves to hold investment trade-off between light resources acquisition and material storage to build leaf organs in various habitats. The K. candel could adjust live strategies to cope with stressed factors and develop strong environment adaptability, and therefore K. candel was suitable to be planted as the coastal shelter forest trees.
Key words Kandelia candel; uneven-aged leaves; leaf traits
doi 10.3969/j.issn.1000-2561.2017.04.007
植物葉片是維持陸地生態系統機能的最基本要素,是大氣-植物系統能量交換的基本單元[1],對環境變化具有重要的指示意義[2]。植物葉片適應環境變化所形成的生存對策[3-6]稱為植物葉性狀(leaf traits),葉性狀作為植物性狀的重要定量指標,能夠反映植物因環境的變化而形成的內在生理及外在形態方面的適應對策及植物資源的利用能力,被認為是環境狀況的敏感指示者[7],具有重要的生態學及植物生理學研究價值[8-10]。
隨著對自然環境變化研究的深入,葉性狀成為研究植物對環境的適應機理的熱點[11]。前人研究認為,植物往年生葉片因生活史較長,結構成熟固化,葉片中很大一部分物質用于增加葉肉細胞密度或構建保衛結構[12-14],因此,面對環境因子變化時,可能響應不敏感[15];此外,隨葉齡增加,葉片衰老過程中結構改變、葉片內部CO2導度下降、擴散受阻,葉內主要養分元素含量及葉片光合能力衰退,由此引起的資源再分配導致異齡葉對環境響應表現出較大的差異[16-18]。而當年生葉處于葉片生長初期,需投入更多資源于強化光合器官、獲得最大的光資源競爭優勢[19-20],以增大光截獲能力去補償自身建成消耗來應對環境變化[21],也有學者認為,展葉過程中葉片的光合作用強度與葉壽命成反比[22],葉齡低的個體,一般要比葉齡高的光合能力強,對于水分脅迫的調節能力較往年生強[23],因此,當環境因子發生擾動時,當年生葉對環境的響應可能較生長后期的葉片更為敏感[24]。由于當年生葉與往年生葉在獲取資源能力及維持生態系統穩定性方面具有不同的生態功能,由此推斷,海岸濕地植物異齡葉間的葉性狀因生長策略、生活史不同可能產生差異。平均葉面積(mean leaf area,MLA)、比葉面積(specific leaf area,SLA)和葉干物質含量(leaf dry matter content,LDMC)是植物葉片的三個關鍵葉性狀,綜合反應植物利用資源的能力以及適應環境的生存對策。其中,MLA反映植物獲取光資源的能力;SLA反映了葉片捕獲光照資源的能力和形成干物質的結果;LDMC為葉片的干重與鮮重之比,表征了葉片在自然狀態下的含水率狀況。前人對MLA、SLA和LDMC等主要葉片特性之間的關系進行大量研究[25],項目組也曾研究葉片在不同環境下的適應策略[26]及海岸山地優勢樹種葉性狀海拔資源位特征[27-28]、薛立等[29]在逆境情況下預測葉性狀變化特征、王希群等[30]對葉面積指數進行研究、黃海俠等[31]分析常綠植物新老葉特征等,但研究海岸濕地樹種異齡葉片性狀差異的研究仍未見報道[32]。
研究地位處福建省羅源灣濱海濕地,地方為發展經濟,近年在灣區興建鋼鐵廠等重工業,這些外部因素促使環境因子明顯改變,秋茄葉可能對這種環境變化產生響應,因此從不同林分類型秋茄異齡葉片受環境影響產生差異角度探尋海岸濕地植物葉片對環境變化的響應規律,對推動我國東南沿海紅樹林恢復、保護提供科學依據。
1 材料與方法
1.1 材料
研究區域位于福建省福州市羅源灣濱海區(26°23′~26°39′N,119°07′~119°54′E),全區東西長約25 km,南北寬約15 km,總面積約230 km2。羅源灣口窄腹大,海灣曲折,呈倒葫蘆狀,在東北角缺口通東海,屬中亞熱帶海洋性季風氣候,年平均氣溫19 ℃,年平均降水量1 652 mm,年平均日照時數1 691.1 h。研究區曾大量分布秋茄紅樹林,后被互花米草入侵替代,當地林業部門治理互花米草后,在清理跡地多次人工栽植多種紅樹林樹種,唯密植(株距0.5 m)的秋茄幼苗成功存活。現復植秋茄林成林面積33 hm2,林相整齊,平均樹高2 m,林齡3 a。天然秋茄林均高2.5 m,林齡5 a,株距0.9 m,分布均勻,長勢旺盛。
1.2 方法
按多點混合法采取植物樣品,在人工林與天然林各設5塊10 m×10 m樣地,每樣地選擇3株標準木進行葉片采集。采集葉片時,依據枝條上芽鱗痕形成的節環來區分不同年齡枝段,當年生枝段上已完全展開的葉片定義為當年生葉片,其他較老枝條上的葉片統稱為往年生葉片。使用枝剪從每株個體的4個不同方向各采剪10片完好無損的葉片。采集葉片后打濕處理,放入自封袋密封并放置水桶內帶回。在12 h內,從每個自封袋中取40片完好無損的葉片清洗擦干,稱其鮮重。用葉面積儀CI-203(美國產)測定樣品葉面積(S),每葉重復3次。將葉片裝入信封置于干燥箱中以80℃烘干至恒重,再稱量其干重。
通過以下公式計算出每株葉片的MLA(cm2)、SLA(m2/kg)和LDMC(mg/g):
為避免從單一植物類型討論植物葉片性狀變異,從天然林與人工林兩個角度探討闊葉樹種異齡葉片的變異性。在相同條件下,對同一被測量進行連續多次測量,所得結果的一致性配對數據t檢驗來判斷葉片性狀在異齡葉片之間的差異。計算異齡葉性狀變異系數,以體現葉性狀變異程度。
1.3 數據與分析
采用Excel 2016軟件對試驗所得數據進行均值、標準差、變異系數的計算,運用SPSS 19.0中的單因素方差分析法(One-way ANOVA)和多重比較(least-significant difference,LSD)法對比同一林分不同葉齡間及不同林分相同葉齡間各性狀的差異。
2 結果與分析
2.1 異齡葉性狀間的差異性
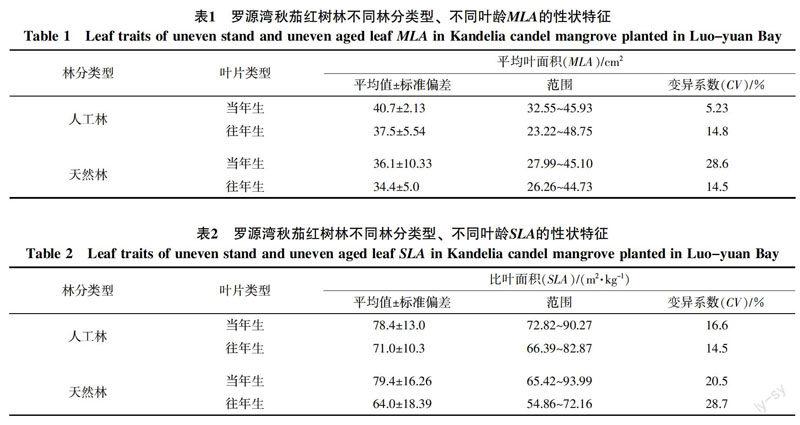
從異齡葉性狀比較分析,羅源灣秋茄紅樹植物異齡葉MLA、SLA和LDMC表現出一定規律性(表1~3)。總體上,人工林與天然林葉面積MLA、SLA與LDMC當年生均大于往年生,分別為(38.4 cm2>36.0 cm2)、(78.9 m2/kg>67.5 m2/kg)、(258.7 mg/g>193.8 mg/g)。①對同種林分不同葉齡的秋茄林而言,人工林與天然林當年生MLA大于往年生,分別為(40.7 cm2>37.5 cm2)、(36.1 cm2>34.4 cm2)。SLA當年生也大于往年生,分別為(78.4 m2/kg>71.0 m2/kg)、(79.4 m2/kg>64.0 m2/kg);LDMC同樣大于往年生,分別為(344.4 mg/g>225.6 mg/g)、(173.0 mg/g>162.0 mg/g)。②從不同林分相同葉齡的秋茄林而言,人工林當年生與往年生葉MLA均大于天然林當年生與往年生葉片,分別為(40.7 cm2>36.1 cm2)、(37.5 cm2>34.4 cm2)。人工林當年生葉SLA小于天然林當年生葉,為(78.4 m2/kg<79.4 m2/kg),往年生相反,為(71.0 m2/kg>64.0 m2/kg)。人工林當年生與往年生LDMC均大于天然林,分別為(344.4 mg/g>173.0 mg/g)、(225.6 mg/g>162.0 mg/g)。
2.2 異齡葉性狀的變異性
變異系數是衡量植物變異程度的一個統計量,變異系數大說明性狀可塑性較強。對秋茄林葉性狀變異系數研究發現:①在同種林分不同葉齡的秋茄林變異系數上,人工林當年生MLA小于往年生,為(5.23%<14.77%),天然林MLA卻相反,為(28.6%>14.5%);SLA變異系數表現出與MLA相反的規律,為人工林當年生變異系數大于往年生,天然林當年生變異系數小于往年生;其值為(16.6%>14.5%)、(20.5%<28.7%);人工林LDMC變異系數小于往年生,分別為(13.7%>9.7%)、天然林變異系數相等(18.7%<18.1%)。②在不同林分相同葉齡的秋茄林變異系數上,人工林當年生MLA變異系數明顯小于天然林當年生,為(5.23%<28.6%)、人工林往年生變異系數大于天然林往年生,為(14.8%>14.5%);人工林當年生與往年生SLA變異系數明顯小于天然林當年生與往年生,為(16.6%<20.5%)、(14.5%<28.7%)。人工林當年生與往年生LDMC變異系數小于天然林當年生與往年生,為(13.7%<18.7%)、(9.7%<18.1%)。
3 討論
常綠植物葉片生活史較長,面臨復雜多變的環境狀況,為適應環境條件,致使葉面積在全球范圍內變化達到6個數量級[33],因此,葉片性狀(MLA、SLA和LDMC)反映了植物適應性進化過程及生活策略差異[34]。本研究中,秋茄MLA分布于23.22~48.75 cm2,人工林的MLA均值當年生與往年生分別為40.7、37.5 cm2,天然林的MLA均值當年生與往年生分別為36.1、34.4 cm2,均表現出當年生MLA大于往年生MLA的規律。前人認為,植物葉片面積受養分[35]、溫度[36]等環境因素影響;但本研究結論表明,葉齡也明顯影響葉面積大小。本研究中,秋茄葉MLA當年生大于往年生的規律與黃海俠等[31]研究天童山常綠闊葉植物MLA結論相反,這可能因秋茄為海岸濕地樹種,其生境較陸生植物惡劣,林分密度遠高于陸生植物(本研究區秋茄人工林株距為50 cm2、天然林株距為90 cm2),以抵御潮汐水流的沖擊。這導致植物個體之間空間競爭激烈,植物必須盡可能擴大占據較優生態位的新葉葉面積,獲取更多光照等資源以滿足自身需求導致。光照條件是影響葉面積大小的重要因素,更大的葉面積被認為是應對低光環境的生態學策略[37]。在灣區內新建的鋼鐵廠等重工業排放,造成空氣中反射太陽光的顆粒物大量增加,可能導致新葉在一定程度上面臨光強較往年降低,為彌補自身需求,獲取最大的光資源和碳生產,當年生葉片在生長初期就將葉片面積展開至最大[38-39]。秋茄SLA人工林與天然林當年生大于往年生,分別為(78.4 m2/kg>71.0 m2/kg)、(79.4 m2/kg>64.0 m2/kg),說明SLA大的葉片,其葉片干物質含量較小,同時較大的葉片面積有利于捕獲更多的光能[40]。SLA還體現植物構建葉面積時所投資的單位生物量, 與葉片建成消耗負相關[41],秋茄往年葉較當年葉承受更長時間的環境脅迫,在防御保護構造上的額外投資較多[34],導致往年葉的建成消耗大于當年葉。
秋茄人工林與天然林當年葉與往年葉的各性狀指標具有明顯差別。天然林林齡較長,從其樹高(2.5 m)、株距(0.9 m)可以看出,葉片空間結構及生長策略相對穩定。而人工林林齡只有2 a,為適應環境脅迫,林分密度較大(株距0.5 m),造成林分內個體競爭激烈,葉性狀指標較天然林更高。也說明秋茄容易受生境因子脅迫,調整生活策略應對,有較強環境適應性,適宜作為海岸防護林樹種人工栽植。
總之,人工林和天然林異齡葉性狀指標均具有明顯差別,這反映異齡葉對光資源獲取能力、儲存物質構建葉器官這兩種投資策略的權衡,是植物在各自生境中適應性進化的結果。
參考文獻
[1] 毛 偉, 李玉霖, 張銅會, 等. 不同尺度生態學中植物葉性狀研究概述[J]. 中國沙漠, 2012, 32(1): 33-41.
[2] 張 林, 羅天祥, 鄧坤枚, 等. 云南松比葉面積和葉干物質含量隨冠層高度的垂直變化規律[J]. 北京林業大學學報, 2008, 30(1): 40-44.
[3] Ackerly D. Self-Shading, Carbon gain and leaf dynamics: a test of alternative optimality models[J]. Oecologia, 1999, 119(3): 300-310.
[4] Coley P D. Effects of plant-growth rate and leaf lifetime on the amount and type of anti-herbivore defense[J]. oecologia, 1988, 74(4): 531-536.
[5] Vendramini F, Díaz S, Gurvich D E, et al. Leaf traits as indicators of resource-use strategy in floras with succulent species[J]. New Phytologist, 2002, 154(1): 147-157.
[6] Westoby M. A leaf-height-seed(LHS0plant ecology strategy scheme[J]. Plant and soil, 1998, 199(20): 213-227.
[7] 許松葵, 薛 立. 6種闊葉樹種幼林的葉性狀特征[J].西北林學院學報, 2012, 27(6): 20-25.
[8] Chapin F S, Bloom A J, Field C B, et al, Plant-responses to multiple environmental-factors[J]. Bioscience, 1987, 37(1): 49-57.
[9] Wright I J, Groom P K, Lamont B B, et al. Leaf trait relationships in Australian plant species[J]. Functional Plant Biology, 2004, 31(5): 551-558.
[10] 張 林, 羅天祥. 植物葉壽命及其相關葉性狀的生態學研究進展[J]. 植物生態學報, 2004, 28(6): 844-852.
[11] 李永華, 羅天祥, 盧 琦. 青海省沙珠玉治沙站17種主要植物葉性因子的比較[J]. 生態學報, 2005, 25(5): 994-999.
[12] Zhang L, Luo T. Advances in ecological studies on leaf lifespan and associated leaf traits[J]. Acta Phytoecologica Sinica, 2004, 28(6): 844-852.
[13] Westoby M. The worldwide leaf economics spectrum[J]. Nature, 2004, 428(6 985): 821-827.
[14] Brian Anacker, Nishanta Rajakaruna, David Ackerly, et al. Ecological strategies in California chaparral: interacting effects of soils, climate, and fire on specific leaf area[J]. Plant Ecology & Diversity, 2011, 4(2-3): 179-188.
[15] Oguchi R, Hikosaka K, Hirose T. Does the photosynthetic light-acclimation need change in leaf anatomy?[J]. Plant Cell & Environment, 2003, 26(4): 505-512.
[16] Field C. Allocating leaf nitrogen for the maximization of carbon gain: Leaf age as a control on the allocation program[J]. Oecologia, 1983, 56(2): 341-347.
[17] Bleecker A B. The evolutionary basis of leaf senescence: Method to the madness?[J]. Current Opinion in Plant Biology, 1998, 1(1): 73-78.
[18] Ono K, Nishi Y, Watanabe A, et al. Possible Mechanisms of Adaptive Leaf Senescence[J]. Plant Biology, 2001, 3(3): 234-243.
[19] Field C, Mooney H A. Leaf age and seasonal effects on light, water, and nitrogen use efficiency in a California shrub[J]. Oecologia, 1983, 56(2): 348-355.
[20] 夏瑩瑩, 毛子軍, 馬立祥, 等. 水分條件對紅松和西伯利亞紅松針葉脯氨酸與葉綠素含量的影響[J]. 植物研究, 2008, 28(3): 330-335.
[21] Schaffer W M, Fox G A. Plant strategies and the dynamics and structure of plant communities[J]. The Quarterly Review of Biology, 1989, 64(2): 409-411.
[22] Warren C R. Why does photosynthesis decrease with needle age in Pinus pinaster?[J]. Trees, 2006, 20(2): 157-164.
[23] Sultan S E. Phenotypic plasticity for plant development, function and life history[J]. Trends in Plant Science, 2000, 5(12): 537-542.
[24] Westoby M. The worldwide leaf economics spectrum[J]. Nature, 2004, 428(6985): 821-827.
[25] ülo Niinemets. Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs[J]. Ecology, 2001, 82(2): 453-469.
[26] 王英姿, 洪 偉, 吳承禎, 等. 靈石山米櫧林優勢種群不同葉齡葉屬性的研究[J]. 福建林學院學報, 2009, 29(3): 203-209.
[27] 洪 滔, 吳承禎, 陳 燦, 等. 福建海岸山地5種優勢樹種葉形態特征對海拔資源位的響應[J]. 福建林學院學報, 2013, 33(1): 7-11.
[28] 洪 滔, 吳承禎, 陳 燦, 等.閩東南海岸山地幾種樹種海拔資源位的葉綠素特征[J]. 福建農林大學學報, 2013, 42(1): 35-39.
[29] 薛 立, 曹 鶴. 逆境下植物葉性狀變化的研究進展[J]. 生態環境學報, 2010, 19(8): 2 004-2 009.
[30] 王希群, 馬履一, 賈忠奎, 等. 葉面積指數的研究和應用進展[J]. 生態學雜志, 2005, 24(5): 537-541.
[31] 黃海俠, 楊曉東, 孫寶偉, 等. 浙江天童常綠植物當年生與往年生葉片性狀的變異與關聯[J]. 植物生態學報, 2013, 37(10): 912-921.
[32] 李善家, 蘇培璽, 張海娜, 等.荒漠植物葉片水分和功能性狀特征及其相互關系[J]. 植物生理學報, 2013, 49(2): 153-160.
[33] Milla R, Reich P B. The scaling of leaf area and mass: the cost of light interception increases with leaf size[J]. Proceedings Biological Sciences, 2011, 274(1 622): 2 109-2 115.
[34] Weiner J. Allocation, plasticity and allometry in plants[J]. Perspectives in Plant Ecology Evolution & Systematics, 2004, 6(4): 207-215.
[35] Roth-Nebelsick A, Kerp H. Evolution and function of leaf venation architecture: A review [Review][J]. Annals of Botany, 2001, 87(5): 553-566.
[36] Yang D M, Zhang J J, Zhou D, et al. Leaf and twig functional traits of woody plants and their relationships with environmental change: A review[J]. Chinese Journal of Ecology, 2012, 31(3): 702-713.
[37] Mcdonald P G, Fonseca C R, Westoby M. Leaf-Size Divergence along Rainfall and Soil-Nutrient Gradients: Is the Method of Size Reduction Common among Clades?[J]. Functional Ecology, 2003, 17(1): 50-57.
[38] Williams K, Percival F, Merino J, Mooney H A. Estimation of tissue construction cost from heat of combustion and organic nitrogen content. Plant[J]. Cell & Environment, 1987, 10, 725-734.
[39] Wright I J, Wearoby M.Cross-species relationships between seedling relative growth rate, nitrogen productivity and root vs.leaf function in 28 Australian woody species[J]. Funct Ecol, 2000, 14(1): 97-107.
[40] Feng Y L, Fu G L, Zheng Y L. Specific leaf area relates to the differences in leaf construction cost, nitrogen allocation and use efficiency between invasive and noninvasive congeners of two genera[J]. Planta, 2008, 228(3): 383-390.
[41] And B F C, Hicks D J. The Ecology of Leaf Life Spans[J]. Annual Review of Ecology & Systematics, 1982, 13(1): 229-259.
