外源ABA對木薯葉片β—胡蘿卜素合成通路相關(guān)基因表達(dá)的影響
2017-05-30 10:48:04鄧昌哲秦于玲李開綿陳松筆
熱帶作物學(xué)報 2017年4期
鄧昌哲 秦于玲 李開綿 陳松筆

摘 要 探究不同濃度外源ABA對木薯葉片β-胡蘿卜素合成途徑相關(guān)基因的影響。用HPLC測定β-胡蘿卜素含量的變化,用qRT-PCR的方法研究木薯胡蘿卜素合成途徑相關(guān)基因的表達(dá)水平。噴施20 mg/L外源ABA,木薯葉片β-胡蘿卜素含量升高,葉片中涉及類胡蘿卜素合成的基因LYCB上調(diào)表達(dá),而類胡蘿卜素裂解的CCD1基因下調(diào)表達(dá)。同時,核心酶PSY1與PSY2基因的下調(diào)表達(dá)與類胡蘿卜素含量的變化無關(guān)聯(lián)。而分別采用10 mg/L與80 mg/L濃度噴施后β-胡蘿卜素含量均降低,涉及β-胡蘿卜素合成通路類胡蘿卜素合成相關(guān)基因表達(dá)水平下降,而經(jīng)10 mg/L的外源ABA處理催化類胡蘿卜素裂解的CCD1基因上調(diào)表達(dá)。研究表明太低和太高濃度的外源ABA處理木薯葉片導(dǎo)致β-胡蘿卜素積累降低,但外源ABA濃度為20 mg/L時能顯著提高葉片β-胡蘿卜素含量。
關(guān)鍵詞 木薯葉片;脫落酸;β-胡蘿卜素含量;β-胡蘿卜素合成途徑; 基因表達(dá)
中圖分類號 Q78;S533 文獻(xiàn)標(biāo)識碼 A
Abstract The effects of different concentrations of exogenous ABA on the genes regarding β-carotene biosynthesis pathways on cassava leaves were investigated. HPLC was used to detect the change of β-carotene content, and RT-PCR method was applied to analyze the change of the gene expression regarding β-carotene biosynthetic pathway. Under the treatment with 20 mg/L of exogenous ABA, β-carotene content increased in cassava leaves, in which LYCB gene, associated with carotenoid synthesis, was up-regulated, and CCD1 gene related with carotenoid cleavage was down-regulated. Meanwhile, the down-regulation of core-enzyme genes PSY1 and PSY2 expressions was no correlation with the change of carotenoid content. The treatments with 10 mg/L and 80 mg/L of exogenous ABA, respectively, resulted in the decrease of β-carotene content in cassava leaves. The genes involved in the carotenoid biosynthetic were down-regulated, and CCD1 gene regarding carotenoid cleavage was up-regulated using 10 mg/L of exogenous ABA. The results showed that the treatments with too low and too high concentrations of exogenous ABA led to the decrease of β-carotene accumulation, however, using 20 mg/L of exogenous ABA could significantly increase the β-carotene content.
Key words Cassava leaves; abscisic acid; β-carotene content; β-carotene synthesis pathway; genes expression
doi 10.3969/j.issn.1000-2561.2017.04.013
木薯(Manihot esculenta Crantz)為大戟科木薯屬植物,與馬鈴薯和紅薯并列世界三大薯類作物;具有高光效和高淀粉積累特性,耐干旱和貧瘠,對環(huán)境適應(yīng)能力強,是世界熱區(qū)近8億人的主糧[1-3]。木薯葉片中富含胡蘿卜素,而塊根缺乏。因此,在非洲等地,長期食用缺乏類胡蘿卜素木薯塊根容易導(dǎo)致夜盲癥的發(fā)生,而適當(dāng)?shù)囊阅臼砣~片為食能夠補充缺少類胡蘿卜素帶來的傷害。前人研究表明,類胡蘿卜素合成的直接前體是牻牛兒基牻牛兒基焦磷酸(GGPP)。類胡蘿卜素合成起始于兩分子GGPP的縮合反應(yīng),由八氫番茄紅素合成酶(PSY)催化兩分子GGPP形成沒有顏色的八氫番茄紅素。該反應(yīng)是類胡蘿卜素合成的關(guān)鍵限速位點[4]。在植物中PSY通常由多基因編碼,不同PSY基因表達(dá)具有明顯的組織特異性及對不同環(huán)境響應(yīng)的特性。木薯中存在3個PSY基因,其中PSY1和PSY2在植物葉片與塊根中表達(dá),而PSY3在木薯中幾乎不表達(dá)[5]。八氫番茄紅素經(jīng)過脫氫酶(PDS)和異構(gòu)化酶催化(ZDS)后形成有顏色的番茄紅素,在水稻胚乳中脫氫反應(yīng)也是重要的限速環(huán)節(jié)[6]。植物中存在兩條支流(LYCB和LYCE)控制番茄紅素向下轉(zhuǎn)化,使得類胡蘿卜素合成途徑中出現(xiàn)β和ε兩條分支。在β分支中,番茄紅素分子的兩端被LYCB環(huán)化形成β-胡蘿卜素;在ε路徑中,番茄紅素經(jīng)LYCB和LYCE環(huán)化后形成α-胡蘿卜素[7]。通過氨基酸序列比對,LYCB和LYCE可能有共同的起源[8]。番茄成熟過程中LCYB基因的下調(diào)是導(dǎo)致果實番茄紅素積累的原因[9]。在非血紅素鐵加氧酶類羥化酶(HYD)的作用下,環(huán)化后的類胡蘿卜素形成β,β-類胡蘿卜素[10]。在植物中類胡蘿卜素的降解主要是由類胡蘿卜素雙加氧酶(CDs)完成的,其分為9-順式-環(huán)氧類胡蘿卜素雙加氧酶(NECDs)和類胡蘿卜素裂解雙加氧酶(CCDs)兩大類。CCDs是一種類胡蘿卜素氧化酶,在特定位點裂解氧化類胡蘿卜素。CCD1能夠在類胡蘿卜素的C9-C10位上剪切番茄紅素、胡蘿卜素、玉米黃質(zhì)或脫輔基類胡蘿卜素等,從而形成芳香類物質(zhì)[11]。
脫落酸(Abscisic acid,ABA)是高等植物重要的激素之一,在逆境下如高溫、干旱等能夠顯著降低對葉綠體超微結(jié)構(gòu)的破壞,并誘導(dǎo)某些酶重新合成,增加植物的抗逆性[12-13]。同時,ABA與類胡蘿卜素合成途徑有著密切的聯(lián)系。類胡蘿卜素是合成ABA的前體物質(zhì),ABA的合成不僅受到內(nèi)源NCED基因表達(dá)水平的影響[14],同時上游類胡蘿卜素途徑PSY的超量表達(dá)也能夠增加ABA的含量[15]。在擬南芥中外源ABA處理6 h后許多與ABA合成和分解的基因也受誘導(dǎo)調(diào)節(jié)[16]。此外,也有研究表明外源噴施ABA能夠增加番茄果實與種子的ABA含量[17]。本研究采用不同濃度的外源ABA處理水培生長的木薯植株,探究類胡蘿卜素合成途徑的相關(guān)基因及β-胡蘿卜素含量在處理后的變化,揭示外源ABA在木薯葉片類胡蘿卜素代謝中的作用,為今后研究木薯類胡蘿卜素積累的調(diào)控機制提供科學(xué)依據(jù)。
1 材料與方法
1.1 材料
供試材料木薯栽培種華南9號(SC9)采自中國熱帶農(nóng)業(yè)科學(xué)院熱帶作物品種資源研究所國家木薯種質(zhì)資源圃。選取粗細(xì)均一,長度20 cm的SC9種莖放入干凈的塑料杯(杯高15 cm,直徑9 cm)中,加入150 mL的自來水,置于常溫,進(jìn)行培養(yǎng)。每間隔3 d更換1次營養(yǎng)液。當(dāng)水培至40 d后,出現(xiàn)18~25片展開葉,選取葉片長勢均一的植株,整株施用外源ABA。實驗設(shè)計3個ABA濃度,分別為低濃度10 mg/L、中濃度20 mg/L和高濃度80 mg/L,每個濃度處理6株,以噴施清水作為對照。處理6 h后,取出植株,用清水洗凈葉片,迅速置于液氮速凍,放置于-80 ℃冰箱中保存?zhèn)溆谩?/p>
1.2 方法
1.2.1 葉片類胡蘿卜素的提取 葉片類胡蘿卜素的提取參照Lucia等[18],稱取0.5 g SC9功能葉片液氮碾磨至粉末狀,轉(zhuǎn)入10 mL離心管,加入2 mL冷丙酮渦混,再加入2 mL冷凍石油醚渦混。4 ℃,3 000 r/min,離心5 min后取上清液,轉(zhuǎn)入10 mL離心管,剩余殘渣加入1 mL石油醚再次渦混、離心,重復(fù)抽提3次,直到殘渣變成無色,收集的樣品液用氮氣吹干,加入500 μL丙酮溶解后轉(zhuǎn)入棕色樣品瓶中。用YCM Carotenoid S-3柱、30 ℃柱溫、甲醇/叔丁基甲醚(70/30, V/V)流動相、1 mL/min流速、20 μL進(jìn)樣量和450 nm測定波長進(jìn)行HPLC分析。
1.2.2 RNA樣品提取 參照TIANGEN RNAprep Pure Plant Kit多糖多酚試劑盒說明書進(jìn)行木薯SC9功能葉片總RNA提取。
1.2.3 RNA反轉(zhuǎn)錄及產(chǎn)物檢測 參照TaKaRa公司的PrimeScriptTM RT regant Kit with gDNA Eraser (Perfect Real Time)進(jìn)行去除基因組及反轉(zhuǎn)錄。
1.2.4 目的基因熒光定量分析 依照TaKaRa公司的SYBR Prime Ex Taq試劑的要求進(jìn)行反應(yīng)液混合:SYBR Prime Ex TaqⅡ5 μL,上游引物0.5 μL,下游引物0.5 μL,cDNA模板1 μL,dH2O 3 μL。PCR反應(yīng)循環(huán)條件為:94 ℃ 1.5 min,94 ℃ 30 s,59 ℃ 30 s,72 ℃ 30 s,一共40個循環(huán);繪制溶解曲線。以噴施清水的表達(dá)量為1,采用2-ΔΔCt 分析不同處理基因的表達(dá)量。每個處理3次生物學(xué)重復(fù)。木薯持家基因(Actin)[19];ζ-胡蘿卜素脫氫酶基因(ZDS)、八氫番茄紅素脫氫酶基因(PDS)、八氫番茄紅素合成酶基因(PSY1, PSY2)、β-胡蘿卜素羥化酶基因(HYD)、胡蘿卜素裂解雙加氧酶基因 (CCD1)、番茄紅素ε-環(huán)化酶基因(LYCE)和番茄紅素β-環(huán)化酶基因(LYCB)的引物見表1。
1.3 數(shù)據(jù)分析
使用Excel 2016和SPSS 22.0統(tǒng)計軟件對數(shù)據(jù)進(jìn)行分析,差異顯著分析采用新復(fù)極差法(Duncan)和最小顯著差異法(LSD)。
2 結(jié)果與分析
2.1 木薯SC9功能葉片在外源ABA處理后β-胡蘿卜素含量的變化
不同濃度外源ABA處理對木薯SC9功能葉片中類胡蘿卜素含量的變化見圖1所示。在不同濃度外源ABA處理下,木薯SC9功能葉片β-胡蘿卜素含量發(fā)生顯著性變化。與對照噴施清水相比,噴施10 mg/L ABA,葉片β-胡蘿卜素含量顯著降低;當(dāng)外源ABA濃度升高至20 mg/L時,葉片β-胡蘿卜素含量顯著高于對照;然而隨著外源ABA濃度上升至80 mg/L,葉片β-胡蘿卜素含量顯著下降,結(jié)果與噴施ABA 10 mg/L濃度相近。研究表明采用太低和太高濃度的外源ABA處理木薯葉片導(dǎo)致β-胡蘿卜素積累降低,但外源ABA濃度為20 mg/L時能顯著提高葉片β-胡蘿卜素含量。
2.2 類胡蘿卜素合成途徑相關(guān)基因的表達(dá)水平分析
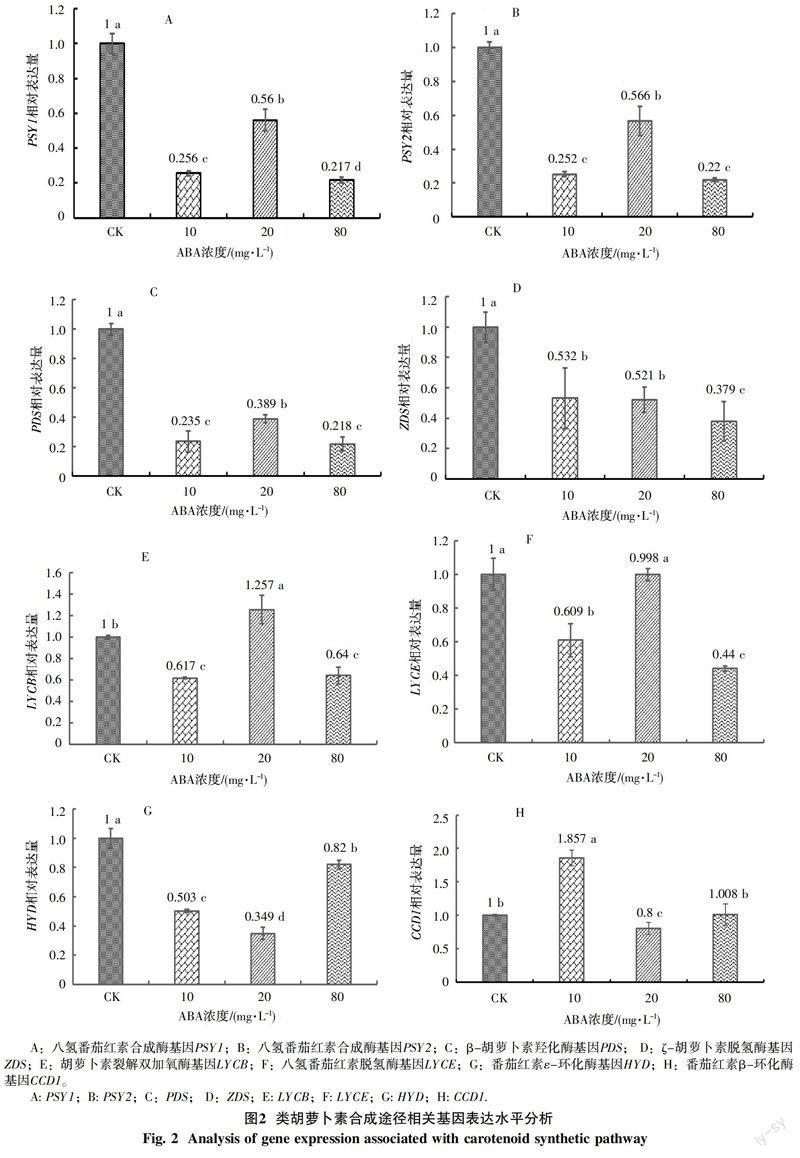
采用不同濃度ABA噴施木薯品種SC9葉片,6 h后分析葉片中類胡蘿卜素合成途徑相關(guān)基因表達(dá)水平的變化,結(jié)果如圖2所示,噴施20 mg/L ABA時,PSY1和PSY2基因在葉片的表達(dá)水平顯著高于10 mg/L與80 mg/L ABA處理的表達(dá)水平,但低于清水對照(圖2-A、B)。PDS催化類胡蘿卜素的脫氫,在外源ABA處理下,其表達(dá)水平均受到顯著抑制,但噴施20 mg/L ABA的表達(dá)水平要高于10 mg/L和80 mg/L處理(圖2-C)。ZDS在較低濃度的外源ABA處理(10 mg/L和20 mg/L)時,其表達(dá)量顯著高于采用高濃度的ABA處理(80 mg/L),但均低于清水對照(圖2-D)。而LYCB表達(dá)水平在20 mg/L外源ABA處理時達(dá)到最高,顯著高于清水對照;而經(jīng)低濃度(10 mg/L)和高濃度(80 mg/L)外源ABA處理后,其表達(dá)水平顯著性受到抑制(圖2-E)。LYCE的表達(dá)水平在低濃度(10 mg/L)和高濃度(80 mg/L)外源ABA處理后,均受到顯著性抑制;而采用20 mg/L外源ABA處理的表達(dá)水平與清水對照沒有顯著性差異(圖2-F)。HYD在不同濃度的外源ABA處理下,其表達(dá)水平均受到顯著抑制,其中表達(dá)水平受到抑制最強的是20 mg/L外源ABA處理,10 mg/L處理次之(圖2-G)。CCD1編碼的酶能夠催化類胡蘿卜素的降解,在10 mg/L低濃度外源ABA處理下,其表達(dá)水平最高;噴施80 mg/L高濃度ABA,其表達(dá)水平與清水對照沒有顯著差異;但處理濃度為20 mg/L時,CCD1的表達(dá)水平最低,說明此時類胡蘿卜素降解受到抑制(圖2-H)。
3 討論
植物組織中, PSY是八氫番茄紅素合成的關(guān)鍵酶,也是決定類胡蘿卜素含量的關(guān)鍵限速酶;本研究結(jié)果顯示不同濃度的外源ABA處理均使木薯葉片PSY1和PSY2基因的表達(dá)水平受到抑制,但20 mg/L外源ABA處理對PSY1和PSY2基因表達(dá)的抑制最小。前人研究結(jié)果表明干旱脅迫雖然也造成木薯葉片PSY1與PSY2基因的表達(dá)水平下降,但類胡蘿卜素含量并未發(fā)生顯著變化[5],推測木薯PSY基因的表達(dá)水平并非直接影響葉片類胡蘿卜素的含量。不同木薯β-胡蘿卜素含量的差異可能是由于PSY基因的單核苷酸多態(tài)性造成PSY酶活性變化,進(jìn)而影響木薯類胡蘿卜素含量[20]。PDS、ZDS、LCYB、LCYE和CCD1綜合作用影響八氫番茄紅素向α/β-胡蘿卜素的轉(zhuǎn)化進(jìn)程,大量研究表明,LCYE和LCYB的轉(zhuǎn)錄調(diào)控及其編碼酶的活性是決定番茄紅素分支流量的重要因素[21]。本研究采用20 mg/L外源ABA處理導(dǎo)致PDS和ZDS基因的表達(dá)水平低于清水對照,但LCYE基因的表達(dá)水平與清水沒有顯著差異,而LCYB基因的表達(dá)水平顯著高于清水對照,表明向β-胡蘿卜素通路加強;分析催化α/β-胡蘿卜素降解的CCD1基因的表達(dá)水平,發(fā)現(xiàn)采用20 mg/L外源ABA處理,CCD1基因的表達(dá)水平與清水對照相比,顯著受到抑制;因此通過提高合成途徑和降低分解途徑相關(guān)基因表達(dá)水平的綜合作用,使得20 mg/L外源ABA處理導(dǎo)致β-胡蘿卜素的積累顯著高于清水對照(圖1)。低濃度(10 mg/L)和高濃度(80 mg/L)外源ABA處理,導(dǎo)致PDS、ZDS、LCYB和LCYE基因的表達(dá)水平均低于清水對照;此外,與清水對照相比,低濃度(10 mg/L)外源ABA處理還誘導(dǎo)CCD1基因的表達(dá)水平升高,導(dǎo)致α/β-胡蘿卜素降解加大,促使α/β-胡蘿卜素積累降低。
HYD和NCED是促使α/β-胡蘿卜素合成ABA的重要蛋白酶。王淑娟等[22]通過對木薯葉片噴施ABA發(fā)現(xiàn)外源ABA對內(nèi)源ABA含量影響顯著,能抑制ABA合成關(guān)鍵酶NCED基因的表達(dá)。已有研究表明HYD的表達(dá)水平與ABA的含量成正相關(guān)[23],本研究表明噴施外源ABA導(dǎo)致HYD基因表達(dá)降低,促使β-胡蘿卜素含量的高積累(圖1和圖2-G)。前人研究表明適中濃度的外源ABA能夠提高植物中抗氧化酶的活力并減弱葉片氧化脅迫從而提高類胡蘿卜素的含量[24-25],因此ABA對類胡蘿卜素合成有潛在的調(diào)控作用[26]。而類胡蘿卜素是ABA合成的前體物質(zhì),表明外源ABA能夠抑制類胡蘿卜素向下游的轉(zhuǎn)化,本研究結(jié)果也證實了施用外源ABA能夠調(diào)控木薯葉片中β-胡蘿卜素的含量。
類胡蘿卜素的合成途徑較為復(fù)雜,其合成途徑相關(guān)基因的表達(dá)和調(diào)控不僅涉及到轉(zhuǎn)錄,而且還影響到光調(diào)控、轉(zhuǎn)錄后的修飾、蛋白質(zhì)的翻譯和編輯等。至于外源ABA是如何進(jìn)行調(diào)控相關(guān)基因間的作用機制,需要進(jìn)一步的研究。
參考文獻(xiàn)
[1] 張振文, 李開綿, 葉劍秋, 等. 木薯光合作用特性研究[J]. 云南大學(xué)學(xué)報(自然科學(xué)版), 2007, 29(6): 628-632.
[2] 陳冠喜, 李開綿, 葉劍秋, 等. 6 個木薯品種生長發(fā)育及產(chǎn)量性狀的初步研究[J]. 熱帶農(nóng)業(yè)科學(xué), 2009, 29(6): 26-29.
[3] Li K, Zhu W, Zeng K, et al. Proteome characterization of cassava(Manihot esculenta Crantz) somatic embryos, plantlets and tuberous roots[J]. Proteome Science, 2010(8): 10.
[4] Terol J, Naranjo M A, Ollitrault P, et al. Development of genomic resources for Citrus clementina: Characterization of three deep-coverage BAC libraries and analysis of 46 000 BAC end sequences[J]. BMC Genomics, 2008, 9: 423.
[5] Jacobo A, Florian W, Peter B, et al. Characterization of phytoene synthase from cassava and their involvement in abiotic stress-mediated responses[J]. Planta, 2010, 232: 1 251-1 262.
[6] Burkhardt P K, Beyer P, Wunn J, et al. Transgenic rice (Oryza sativa) endosperm expressing daffodil (Narcissus pseudonarcissus) phytoene synthase accumulates phytoene, a key intermediate of provitamin a biosynthesis[J]. Plant Journal, 1997, 11: 1 071-1 078.
[7] Josse E M, Simkin A J, Gaffe J, et al. A plastid terminal oxidase associated with carotenoid desaturation during chromoplast differentiation[J]. Plant Physiology, 2000, 123: 1 427-1 436.
[8] Yu Q, Schaub P, Ghisla S, et al. The lycopene cyclase crty from pantoea ananatis (ex Erwinia uredovora) catalyzes an FADrea-dependent non-redox reaction[J]. Journal of Biological Chemistry, 2010, 285: 12109.
[9] Ronen q Carmel-Goren L, Zamir D, Hirschberg J. An alternative pathway to beta-carotene formation in plant chromoplasts discovered by map-based cloning of Beta and old-gold color mutations in tomato[J]. Proceedings of the National Academy of Sciences of the United States of America, 2000, 97: 11102.
[10] Vallabhaneni R, Wurtzel E T. Timing and biosynthetic potential for carotenoid accumulation in genetically diverse germplasm of maize[J]. Plant Physiology, 2009, 150: 562-572.
[11] Simkin A J, Schwartz S H, Auldridge M, et al. The tomato carotenoid cleavage dioxygenase 1 genes contribute to the formation of the flavor volatiles β-ionone, pseudoionone, and geranylacetone[J]. Plant Journal, 2004, 40: 882-892.
[12] Vallabhaneni R, Bradbury L M T, Wurtzel E T. The carotenoid dioxygenase gene family in maize, sorghum, and rice[J]. Archives of Biochemistry and Biophysics, 2010, 504: 104-111.
[13] Zeevaart J A D, C'reelman R A. Metabolism and physiology of abscisicacid[J]. Annu Rev Plant Physiol Plant Mol Biol, 1988, 39: 439-473.
[14] Marta de Torres-Zabala, Truman W, Bennett M H, et al. Pseudomonas syringae pv tomato hijacks the Araluednpses abscisic acid signalling pathway to cause disease[J]. EMBO J, 2007, 26(5): 1 434-1 443.
[15] Thompson A J S, Burbidge A, Jackson A C, et al. Ectopic expression of a dioxygenase gene causes over-production of abscisic 363-374 tomato 9-cis-epoxycarotenoid acid[J]. Plant J, 2000b, 23(3): 363-374.
[16] Estevez J M, Cantero A, Reindl A, et al. 1-Deoxy-D-xylulose-5-phosphate isoprenoid biosynthesis in plants. synthase, a limiting enzyme for plastidic[J]. J Biol Chem, 2001, 276(25): 22 901-22 909.
[17] Zhang M, Yuan B, Leng P. The role of ABA in triggering ethylene biosynthesis and ripening of tomato fruit[J]. J Exp Bot, 2009, 60: 1 579-1 588.
[18] Lucia M, Alcides R, Ronoel L, et al. Retention of total carotenoid and β-carotene in yellow sweet cassava (Manihot esculenta Crantz) after domestic cooking[J]. Food and Nutrition Research, 2012, 56: 15788-DOI: 10.3042.
[19] 許 娟, 羅興錄. 木薯Actin基因片段的克隆及序列分析[J]. 生物技術(shù)學(xué)報, 2011(6): 65-70.
[20] Ralf Welsch, Jacobo Arango, et al. Provitamin a accumulation in cassava (Manihot esculenta) roots driven by a single nucleotide polymorphism in a phytoene synthase gene[J]. The Plant Cell, 2010, 22: 3 348-3 356.
[21] Cazzonelli C, Pogson B. Source to sink: regulation of carotenoid biosynthesis in plants[J]. Trends in Plant Science, 2010, 15: 266-274.
[22] 王淑娟, 方開星, 王文泉, 等. 外源ABA對木薯葉片內(nèi)源激素及淀粉合成相關(guān)基因影響[J]. 中國農(nóng)業(yè)大學(xué)學(xué)報, 2015, 20(3): 100-107.
[23] Qin G J, Gu H Y, Ma L C, et al. Disruption of phytoene desaturase gene results in albino and dwarf phenotypes in Arabidopsis by impairing chlorophyll, carotenoid, and gibberellin biosynthesis[J]. Cell Res, 2007, 17(5): 471-482.
[24] Zhou X, Mcquinn R, Fei Z, et al. Regulatory control of high levels of carotenoid accumulation in potato tubers[J]. Plant, Cell & Environment, 2011, 34: 1 020-1 030.
[25] 范 敏, 金黎平, 屈冬玉. 等. 干旱脅迫對馬鈴薯類黃酮和類胡蘿卜素合成關(guān)鍵酶基因表達(dá)的影響[J]. 園藝學(xué)報, 2008, 35(4): 535-542.
[26] Agarwal S, Sairam R K, Srivastava G C, et al. Changes in antioxidant enzymes activity and oxidative stress by abscisic acid and salicylic acid in wheat genotypes[J]. Biol Plantarum, 2005, 49(4): 541-550.
