干旱脅迫對黃秋葵幼苗生長以及葉片光合特性的影響
2017-06-12 12:21:04王繼玥陳俊峰卯丹劉燕謝文鋼杜斌
熱帶農業科學 2017年5期
關鍵詞:生長
王繼玥+陳俊峰+卯丹+劉燕+謝文鋼+杜斌+楊丹+宋常美


摘 要 通過PEG模擬干旱脅迫,初步研究干旱脅迫對黃秋葵幼苗生長及葉片光合特性的影響。結果顯示:2個品種的葉柄長、株高、莖粗、根長、苗鮮重、苗干重、最大葉長、最大葉寬、功能葉面積以及抗旱系數均隨著PEG濃度的增加而減少,表明干旱脅迫抑制了黃秋葵幼苗的生長,隨著干旱脅迫程度的增加,其抗性逐漸減弱;2個品種葉片的凈光合速率、氣孔導度均隨PEG濃度的增加而減少,說明干旱脅迫抑制了黃秋葵的光合能力。相關性分析表明,黃秋葵葉柄長、株高、莖粗、根長、苗鮮重、苗干重、最大葉寬、最大葉長、凈光合速率、氣孔導度均與抗旱系數極顯著相關,說明相關性狀及生理生化指標可作為干旱脅迫下黃秋葵幼苗的抗旱性鑒定指標。
關鍵詞 黃秋葵 ;干旱脅迫 ;生長 ;光合特性
中圖分類號 S643.9 文獻標識碼 A Doi:10.12008/j.issn.1009-2196.2017.05.002
Effect of Drought Stress on Seedling Growth and
Leaf Photosynthetic Traits of Okra
WANG Jiyue1) CHEN Junfen1) MU Dan1) LIU Yan1)
XIE Wengang1) DU Bin2) YANG Dan1) SONG Changmei1)
(1 School of Biological and Environmental Engineering, Guiyang University, Guiyang 550005;
2 School of the Food and Pharmaceutical Engineering, Guiyang University, Guiyang 550005)
Abstract Okra seedlings were treated under drought stress simulated with different concentrations of PEG-6000 solutions to observe their growth and leaf photosynthesis. The results showed that the petiole length, plant height, root diameter, root length, seedling fresh weight, dry weight, the largest leaf width, the largest leaf length, functional leaf area per plant and drought-resistance index of okra were significantly decreased with an increase in concentration of PEG-6000 solution. This implied that the growth of okra seedlings was inhibited by drought stress. The resistance of the seedlings to drought stress decreased gradually with the increase of the drought stress. The photosynthetic rate and stomatal conductance also significantly decreased with the increase of the concentration of PEG-6000 solution. This suggested that photosynthesis of the seedlings was suppressed by drought stress. Correlation analysis showed that the petiole length, plant height, root diameter, root length, fresh weight, dry weight, the largest leaf width, the largest leaf length, the photosynthetic rate and stomatal conductance were highly significantly correlated to the drought resistance coefficient. These traits and physiological and biochemical indexes can be used as the indexes to identify drought resistance of okra seedlings under drought stress.
Keywords okra ; drought Stress ; growth ; photosynthetic traits
黃秋葵(Abelmoschus esculentus)是錦葵科秋葵屬,一年生草本植物。,有研究指出其源于非州,經土耳其傳入中國[1]。黃秋葵食藥兼具[2-3],目前中國各地均有種植,以臺灣地區最多。黃秋葵富含各種維生素和礦物質,其根、莖、葉、花皆可入藥[4],應用前景廣泛。
中國干旱區、半干旱和半濕潤易旱區面積占總國土面積50%以上,其中約有耕地5千萬公頃。因此,干旱脅迫是目前農業生產中面臨的最嚴峻問題之一。作物一旦缺水,尤其是處在生長發育的關鍵期,必然抑制作物生長、降低產量,嚴重時甚至絕收[5]。研究表明,缺水影響作物的生長取決于作物生長的時期以及水分脅迫的程度[6]。韋吉[7]等研究表明,干旱脅迫下,結莢期黃秋葵葉片的相對含水量、葉綠素含量下降,離體失水率、相對電導率、游離脯氨酸和可溶性蛋白質含量升高。研究干旱脅迫下植物形態結構、生理和生化過程的改變將有助于揭示植物對干旱脅迫的響應機制,對培育抗旱新品種,利用分子育種手段提高植物的抗旱性具有重要意義。目前對水稻[8]、玉米[9]、大豆[10]等作物抗干旱脅迫機制的研究較多,也培育出一些抗旱作物,但有關蔬菜耐旱機制以及抗性育種的研究相對較少。黃秋葵具有耐旱、耐高溫、適應性廣等特點[11],但對其遺傳與生理機制的研究尚不深入。本文通過PEG-6000模擬干旱脅迫,研究干旱脅迫對黃秋葵幼苗生長以及光合特性的影響,為揭示黃秋葵抗干旱脅迫的生理機制提供理論參考。
1 材料與方法
1.1 材料
2份黃秋葵供試材料由貴陽學院生物與環境工程學院分別從遵義和江蘇收集而來,編號分別為51號(卡里巴)和44號(楊貴妃)。前期通過PEG模擬干旱脅迫對30多份種質資源進行抗旱性鑒定,發現這2個品種具有較強的抗性。51號為綠果,44號為白果,51號鮮果產量高于44號,44號生育期較51號短10~20 d。
1.2 方法
1.2.1 干旱脅迫處理
2份黃秋葵品種中隨機選擇200粒種子,用5%次氯酸溶液消毒后,再用蒸餾水沖洗3次,直播于高25 cm、直徑18 cm營養缽中(內裝80%營養土)。待出苗后,每缽定植長勢一致的5株苗。設3個干旱脅迫處理,分別為5%PEG-6000、10%PEG-6000、15%PEG-6000,每個處理5次重復,每隔2 d澆PEG溶液1次,以水處理為空白對照。隨機擺放在溫室大棚內,大棚內溫度在20~30℃之間,于處理25 d后測定農藝形狀以及光合特性。
1.2.2 測定指標
1.2.2.1 農藝性狀
每處理取生長一致的植株10株,分別測定最大葉長、最大葉寬、總葉片數、展開葉片數、株高、莖粗、葉柄長、根長、苗鮮重和苗干重。采用離體葉面積儀測定葉面積,75℃烘干至恒重,計算比葉重,比葉重=葉片干重/葉片面積。采用厲廣輝[8]的方法計算抗旱系數(DC),DC=Yd/Yp(Yd:干旱脅迫下生物量,Yp:非脅迫下生物量),展葉比=展開葉片數/總葉片數。
1.2.2.2 光合特性
采用Li-6400 便攜式光合測定儀,在上午 9:00~12:00測定光合速率(Pn)、氣孔導度(Gs)以及CO2濃度(Ci),重復測定5次。采用 UV-2450 紫外分光光度計比色,測定主莖中部葉片葉綠素含量,每處理5次重復。
1.2.3 數據處理
利用SPASS 12.0軟件進行數據處理和統計分析。
2 結果與分析
2.1 干旱脅迫對黃秋葵農藝性狀的影響
不同干旱脅迫下黃秋葵的農藝性狀表現見表1。由表1可看出,2個品種的葉柄長、株高、莖粗、根長、苗鮮重、苗干重以及抗旱系數均隨PEG濃度的增加而減少,只是有的差異顯著,有的差異不顯著。2個品種的根長、苗鮮重、葉柄長與對照均差異顯著,而各處理間莖粗均差異不顯著。不同PEG模擬干旱脅迫下,51號品種的抗旱系數均明顯高于44號,表明51號的抗旱性強于44號。
2.2 干旱脅迫對黃秋葵葉片性狀的影響
不同干旱脅迫下黃秋葵葉片性狀表現分析結果(表2)表明,2個品種的最大葉長、最大葉寬,功能葉面積均隨著PEG濃度的增加而減少,比葉重隨著PEG濃度的增加而增加。51號的最大葉長、最大葉寬、功能葉面積隨著PEG濃度增加而減少的幅度高于44號,總葉片數各處理間無差異;44號各處理間展開葉片數無差異,但顯著低于對照;51號不同干旱脅迫下的展開葉片數葉顯著低于對照,但10% PEG處理的展開葉片數顯著高于其余處理。
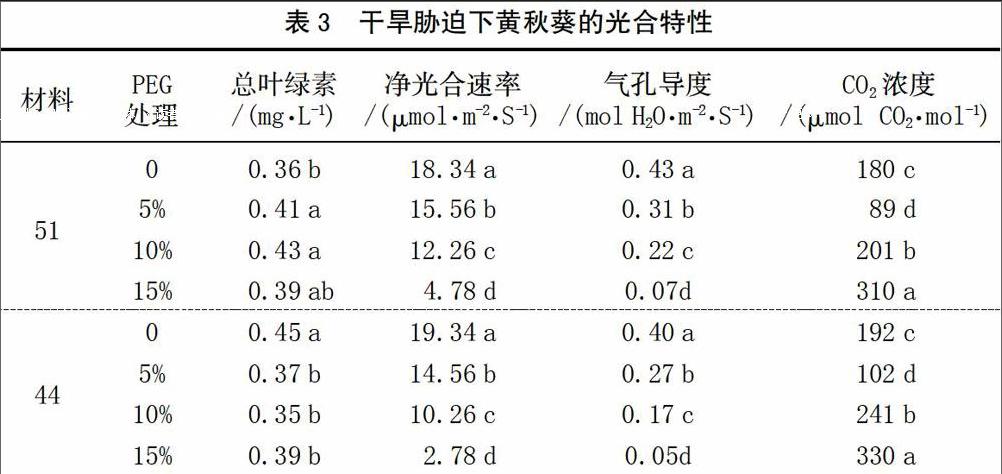
2.3 干旱脅迫對黃秋葵光合特性的影響
干旱脅迫下黃秋葵的光合特性見表3。2個品種葉片的凈光合速率、氣孔導度均隨著PEG濃度的增加而減少,且各處理間均差異顯著。其中51號葉片的凈光合速率、氣孔導度隨著PEG濃度增加而減少的幅度明顯高于44號。2個品種CO2濃度均隨著PEG濃度的增加呈先降低后增加的趨勢,即5%PEG處理顯著低于對照,但10% PEG和15% PEG處理顯著高于對照和5%PEG處理。2個品種的葉片葉綠素含量沒有表現出一致的規律,51號葉片葉綠素含量有隨著PEG濃度的增加而增加的趨勢,但各PEG處理間差異不顯著,且15%PEG處理與對照差異也不顯著。44號葉片葉綠素含量不同濃度PEG下均顯著低于對照,但PEG處理間又差異不顯著。
2.4 相關性分析
相關性分析結果顯示,2個材料的葉柄長、株高、莖粗、根長、苗鮮重、苗干重、最大葉寬、最大葉長、凈光合速率、氣孔導度均與抗旱系數極顯著相關(P<0.01)。
3 討論
本研究基于PEG模擬干旱脅迫實驗,初步研究了干旱脅迫對苗期黃秋葵生長以及葉片光合特性的影響。結果顯示,2個品種葉柄長、株高、莖粗、根長、苗鮮重、苗干重最大葉長、最大葉寬、功能葉面積等農藝性狀及抗旱系數均隨PEG濃度的增加而減少,表明干旱脅迫會抑制黃秋葵幼苗的生長,并且其危害程度也隨干旱脅迫程度的增加而增加。同時,黃秋葵幼苗對干旱脅迫的抗性逐漸降低,這與劉志媛[12]等的研究結果相似。本研究發現,相對于對照處理,不同濃度PEG處理條件下黃秋葵的生長均受抑制,但不同性狀指標的受抑制程度有所不同。2個品種的葉柄長、莖粗、總葉片數、最大葉寬在不同濃度PEG處理下差異均不顯著,說明不同組織和器官對干旱脅迫的響應可能不同。相關性分析表明,葉柄長、株高、莖粗、根長、苗鮮重、苗干重、最大葉寬、最大葉長均與抗旱系數極顯著相關,說明相關性狀可作為黃秋葵幼苗抗干旱脅迫的鑒定指標,這與花生[13]、玉米[14]等作物中的研究結果類似。
2個品種葉片的凈光合速率、氣孔導度均也隨著PEG濃度的增加而減少,說明干旱脅迫抑制了黃秋葵的光合作用,從而導致其胞間的二氧化碳濃度增加。但5%PEG 處理下二氧化碳濃度低于對照,說明黃秋葵幼苗對輕度干旱脅迫具有耐受性,輕度干旱脅迫下仍能維持較強的光合作用以確保植物正常的生長,這與厲廣輝[13]等人的研究結果類似。2個材料不同干旱脅迫下葉片葉綠素的含量并未表現出一致的規律,有的降低,有的增加,表明不同材料葉片葉綠素含量對干旱脅迫的抗性有所不同。李國防[15]則發現,重度干旱脅迫會提高蘋果葉片葉綠素的含量,干旱脅迫提高了葉片中活性氧的含量,而植物體內的活性氧清除劑可降低葉片中的活性氧含量,并提高葉綠素含量。因此,葉片葉綠素含量的提高一定程度上可反映出植物對干旱脅迫的抗性。
光合作用為植物提供能量以維持生長,缺水導致氣孔關閉,從而影響光合速率和蒸騰速率,相關研究[16]表明,植物葉片氣孔相關的生理生化性狀與品種抗干旱脅迫的能力密切相關。因此,王士強等[17]認為,植物的抗旱性可選用氣孔導度、光合速率和蒸騰速率等生理生化指標來鑒定,但必須在干旱脅迫下,因為正常供水條件下并不表現一致的規律。
參考文獻
[1] Ahmadi A, Emam Y, Pessarakli M. Response ofvarious cultivars of wheat and maize to salinity stress[J]. Food Agric Environ, 2009, 7(1): 123-128.
[2] Adelakun O E,Qyelade O J,Ade-omowaye B I,et al. Chemical composition and the antioxidative properties of NigerianOkra seed (Abelmoschus esculentus Moench) flour[J]. Food Chem Toxicol, 2009, 47(6): 1 123-1 126.
[3] Amoatey H, Ahiakpa J, Quartey E, et al. Agromorphological Characterisation of 29 Accessions of Okra (Abelmoschus spp L.)[J]. Journal of Biology, Agriculture and Healthcare, 2015, 5(4): 6-16.
[4] Jarret R, Wang M, Levy I. Seed oil and fatty acid content in okra (Abelmoschus esculentus) and related species[J]. J Agric Food Chem, 2011, 38(8): 419-424.
[5] Tas S. Some physiological responses of drought stress in wheat genotypes with different ploidity in Turkiye[J]. World Journal of Agricultural Sciences,2007, 3: 178-183.
[6] Clavel D, Drame N K, Roy-Macaule H, et al. Analysis of early responses to drought associated with field drought adaptation in four Sahelian groundnut (Arachishypogaea L.) cultivars[J]. Environ Exp Bot, 2005, 54: 219-230.
[7] 韋 吉,黎軍平,羅燕春,等. 不同黃秋葵種質對干旱的生理生化反應[J]. 熱帶農業科學,2008,24(6):452-455.
[8] Wang W, Chen Q, Hussain S, et at. Seed priming in dry direct-seeded rice: consequences for emergence, seedling growth and associated metabolic events under drought stress[J]. Plant Growth Regulation, 2016, 78(2): 167-178.
[9] Masuka B, Araus J L, Das B, et at. Phenotyping for abiotic Stress tolerance in maize: drought stress. A field manual[J]. J Integr Plant Biol,2012, 54(4): 238-249.
[10]James R, Carl R, Philip J. Drought-Stress effects on branch and mainstem seed yield and yield components of determinate soybean[J]. Crop Science,2015, 44(3): 759-763.
[11] 曾日秋,洪建基,姚運法,等. 黃秋葵資源的植物學特征特性與生態適應性評價[J]. 熱帶作物學報,2015,36(3):523-529.
[12] 劉志媛,黨選民,曹振木. 土壤水分對黃秋葵苗期生長和光合作用的影響[J]. 熱帶作物學報,2003,24(1):70-72.
[13] 厲廣輝,張 昆,劉風珍,等. 不同抗旱性花生品種的葉片形態及生理特性[J]. 中國農業科學,2014,47(4):644-654.
[14] 張興華,高 杰,杜偉莉,等. 干旱脅迫對玉米品種苗期葉片光合特性的影響[J]. 作物學報,2015,41(1):154-159.
[15] 李國防.干旱脅迫下蘋果葉片光合機構運轉機制的研究[D]. 楊陵:西北農林科技大學碩士學位論文,2014.
[16] Shardendu K S, Reddy K R. Regulation of photosynthesis, fluorescence, stomatal conductance and water-use efficiency of cowpea (Vigna unguiculata Walp.) under drought[J]. Journal of Photochemistry and Photobiology B: Biology, 2011, 105(1): 40-50.
[17] 王士強,胡銀崗,佘奎軍,等. 小麥抗旱相關農藝性狀和生理生化性狀的灰色關聯度分析[J]. 中國農業科學,2007,40(11):2 452-2 459.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
