利用SSR標(biāo)記分析西瓜自交系的遺傳關(guān)系
2017-06-19 17:03:22許彥賓王艷玲溫新惠胡建斌孫守如馬長生
中國瓜菜 2017年4期
許彥賓,王艷玲,溫新惠,胡建斌,李 瓊,孫守如,馬長生
(1.河南農(nóng)業(yè)大學(xué)園藝學(xué)院 鄭州 450002; 2.平羅縣種子管理站 寧夏平羅 753400)
利用SSR標(biāo)記分析西瓜自交系的遺傳關(guān)系
許彥賓1,王艷玲1,溫新惠2,胡建斌1,李 瓊1,孫守如1,馬長生1
(1.河南農(nóng)業(yè)大學(xué)園藝學(xué)院 鄭州 450002; 2.平羅縣種子管理站 寧夏平羅 753400)
為了明確西瓜自交系間的遺傳關(guān)系,采用SSR和EST-SSR標(biāo)記分析34份高代自交系的基因型,計算各材料間的相似系數(shù),并進行聚類分析。結(jié)果表明,9個基因組SSR和8個EST-SSR標(biāo)記共檢測到83個等位基因,平均4.88個,PIC值變幅為0.112~0.686,平均0.425,基因組SSR的多態(tài)性明顯高于EST-SSR。所有材料聚為A、B、C 3類,B、C 2類包含多態(tài)性較高的4份自交系(X36、X48、X84和X89),均為來自西北地區(qū)的小果型材料,可以用來豐富栽培西瓜的多樣性;A類包含剩余的30份自交系,主要是國內(nèi)外的單果質(zhì)量大于或等于1.5 kg的材料(除X38、X80外),其遺傳多樣性較低,遺傳背景需要進一步拓展。A類可分為3個亞類,每個亞類材料的植物學(xué)性狀和來源地大不相同。研究結(jié)果將為西瓜雜交親本選配和種質(zhì)資源創(chuàng)新提供理論依據(jù)。
西瓜;自交系;SSR;遺傳關(guān)系
雜交育種是目前大多數(shù)園藝作物培育新品種的主要方法,其首要條件是具備一定遺傳差異的親本材料[1]。西瓜是我國主要園藝作物,種植面積和產(chǎn)量位居世界前列[2]。但我國不是西瓜的發(fā)源地,種質(zhì)資源相對匱乏。20世紀(jì)60年代,我國從美國、日本、前蘇聯(lián)等國家引入了許多優(yōu)良西瓜品種,促進了西瓜主要農(nóng)藝性狀的遺傳改良,也極大地推動了我國西瓜品種的更新?lián)Q代[3]。有研究表明,栽培西瓜的遺傳基礎(chǔ)較為狹窄,特別是品種之間DNA水平的差異較小[4-5],限制了雜交親本的選擇。近年來,我國西瓜栽培面積穩(wěn)中有增,消費市場多元化發(fā)展趨勢愈加明顯,品種更新速度也愈來愈快。但由于大多數(shù)西瓜育種者只重視雜交組合的配制而忽視育種材料遺傳關(guān)系評價,頻繁地利用少數(shù)親緣關(guān)系較近的骨干親本配制雜交組合,以致新育成品種的一些重要農(nóng)藝性狀未得到明顯改良,因而難以育成突破性的西瓜新品種。隨著西瓜全基因組測序的完成,利用分子標(biāo)記技術(shù)分析西瓜育種材料的遺傳關(guān)系,有助于育種者在DNA水平上選擇具有差異的育種材料,從而實現(xiàn)對西瓜重要農(nóng)藝性狀的有效改良[6]。
目前,RAPD、AFLP、SSR等多種分子標(biāo)記已用于西瓜的遺傳多樣性分析,其中SSR分子標(biāo)記因其穩(wěn)定性好、多態(tài)性高、共顯性、多等位性等優(yōu)點而應(yīng)用更為廣泛[7-9]。筆者采用SSR標(biāo)記對34份西瓜自交系的遺傳關(guān)系進行分析,以期選擇在DNA水平上具有差異的育種材料,為西瓜雜交親本的選配提供參考依據(jù)。
1 材料與方法
1.1 材料
供試材料為34份西瓜自交系,由河南豫藝種業(yè)科技發(fā)展有限公司提供(表1)。所有材料均是從國內(nèi)外多個西瓜商業(yè)品種中自交分離而得,自交7代以上,主要農(nóng)藝性狀已經(jīng)穩(wěn)定,但自交系的含糖量與雜交種相比普遍偏低。2010—2015年在鄭州滎陽田間連續(xù)種植,性狀調(diào)查見表1(5 a平均值),各材料在果實性狀方面存在較大的差異,可溶性固形物含量取果實中心至邊緣1/2處的果肉測定。

表1 2010—2015年34份西瓜材料及果實性狀調(diào)查
1.2 SSR-PCR擴增
采用CTAB小量法[10]從西瓜幼嫩葉片中提取基因組DNA,利用紫外分光光度計檢測DNA濃度與純度,將其稀釋到終濃度20 ng·μL-1。試驗采用基因組SSR和EST-SSR進行基因型鑒定,基因組SSR引物選自Joobeur等[11]的報道,EST-SSR引物選自Guerra-Sanz[12]和Verma等[13]的報道,篩選多態(tài)性好的SSR引物。PCR反應(yīng)體系15 μL,其中包括:1×Buffer,1.5 mmol·L-1MgCl2,200 μmol·L-1dNTPs,0.4 μmol·L-1引物,1 UTaq酶,50 ng DNA模板。PCR反應(yīng)在PTC-200型PCR擴增儀(MJ Reserch,Inc.)上進行。反應(yīng)程序為:94℃預(yù)變性5 min;94℃變性1 min,55℃復(fù)性45 s,72℃延伸1 min,35個循環(huán);72℃延伸6 min。擴增產(chǎn)物用6%非變性聚丙烯酰胺凝膠電泳分離,進行銀染、顯帶。
1.3 數(shù)據(jù)分析
以PCR條帶在凝膠中相對遷移位置的有無賦值為“1”或“0”,生成分子數(shù)據(jù)矩陣。采用Jaccard[14]的方法計算各材料間的遺傳距離和相似系數(shù),利用NTSYS-pc 2.10軟件中的不加權(quán)成對算術(shù)平均法(UPGMA)對34份自交系進行聚類,繪制樹狀聚類圖。根據(jù)Botstein等[15]報道的方法計算SSR標(biāo)記的多態(tài)性信息含量(PIC,polymorphism information con?tent)值,即PIC=1-∑Xi2,其中Xi表示第i種基因型出現(xiàn)的頻率。
2 結(jié)果與分析
2.1 SSR標(biāo)記的多態(tài)性
根據(jù)文獻報道共合成了35對SSR引物,從中篩選出帶譜清晰、多態(tài)性好的17對。其中,基因組SSR引物9對,EST-SSR引物8對。17對引物對34份西瓜自交系的擴增結(jié)果見表2。所有引物共檢測到83個等位基因,平均每對引物檢測到4.88個,MCPI05檢測到的等位基因數(shù)最多(10個),EST00667檢測到的等位基因數(shù)最少(2個);PIC值變幅 0.112(EST00667)~0.686(MCPI11),平均0.425,圖1為MCPI11電泳圖。基因組SSR的平均等位基因數(shù)和 PIC值分別為 5.89和 0.514,EST-SSR的平均等位基因數(shù)和PIC值分別為3.75和0.326,基因組SSR的多態(tài)性明顯高于EST-SSR。
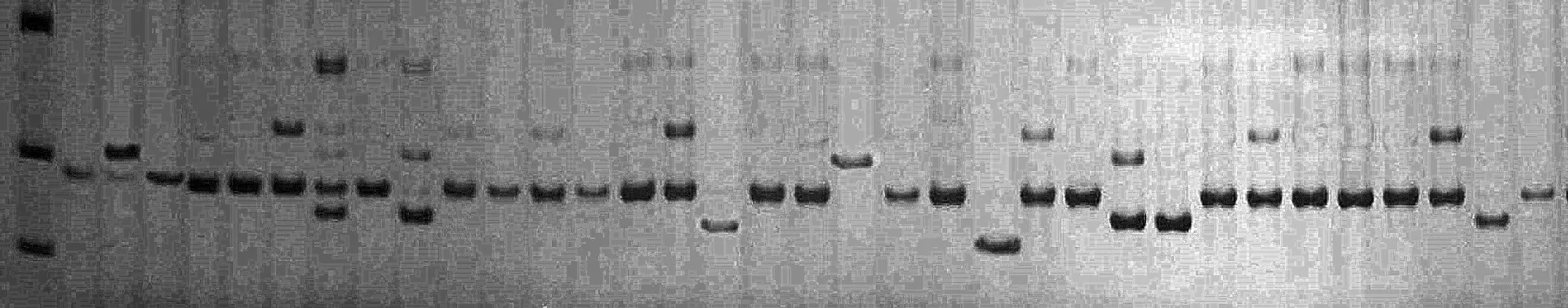
圖1 MCPI11非變性聚丙烯酰胺凝膠電泳圖
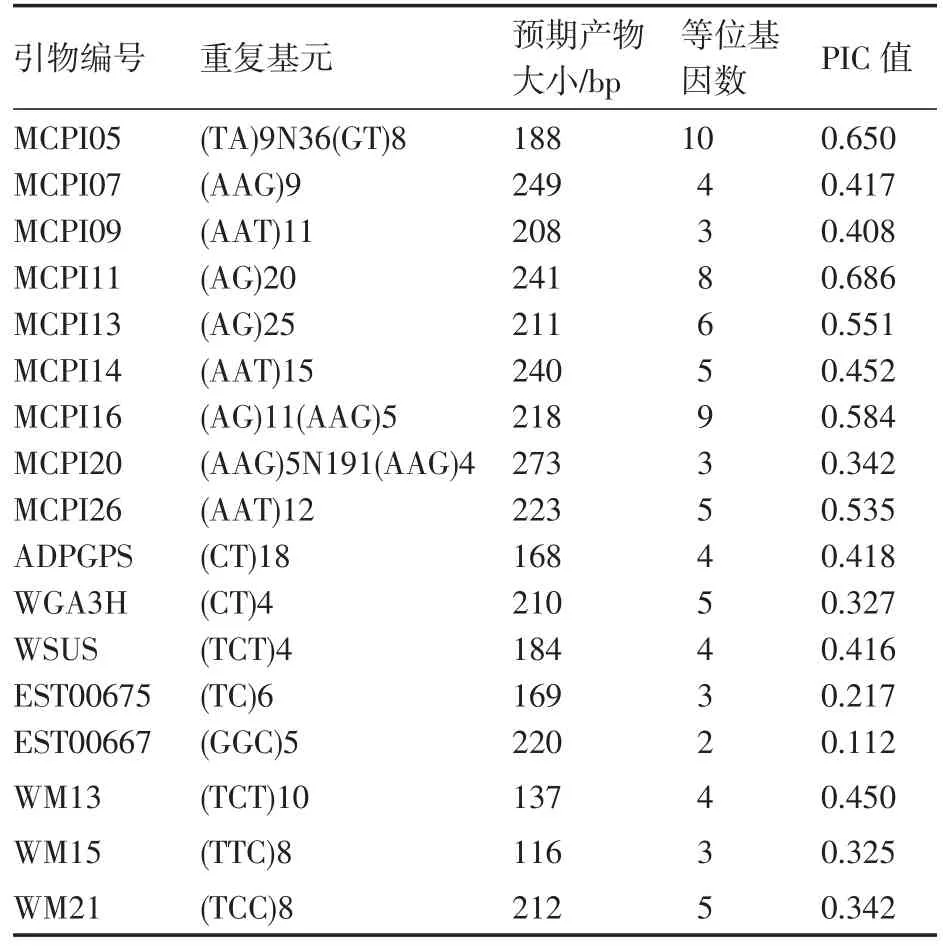
表2 17對SSR引物的特征及其多態(tài)性
2.2 西瓜自交系的遺傳多樣性
為了明確西瓜各自交系之間的遺傳關(guān)系,采用SSR基因型數(shù)據(jù)計算各材料之間的相似系數(shù),獲得34份自交系的相似系數(shù)矩陣。結(jié)果表明,所有材料的相似系數(shù)變幅為 0.24~0.93,平均相似系數(shù)為0.56,X66與X61的相似系數(shù)最大,X82或X85與X36之間的相似系數(shù)最小。如果排除自交系X36,所有材料的平均相似系數(shù)為0.65。上述結(jié)果說明,西瓜自交系之間的遺傳分化較小,遺傳背景較為狹窄。
2.3 西瓜自交系的聚類分析
根據(jù)各材料間的相似系數(shù)對所有自交系進行聚類作圖,以明確其遺傳關(guān)系。圖2表明,34份材料在相似系數(shù)為0.48的水平上可聚為A、B、C共3類。其中,X36單獨成一類(C類),說明其遺傳背景較為獨特。B類也僅包括3份自交系,即X48、X84和X89。這2類所包含的4份材料均為小果型自交系(單果質(zhì)量小于1.2 kg),是從中國西北地區(qū)的西瓜品種中分離所得,具有較高的遺傳多樣性。
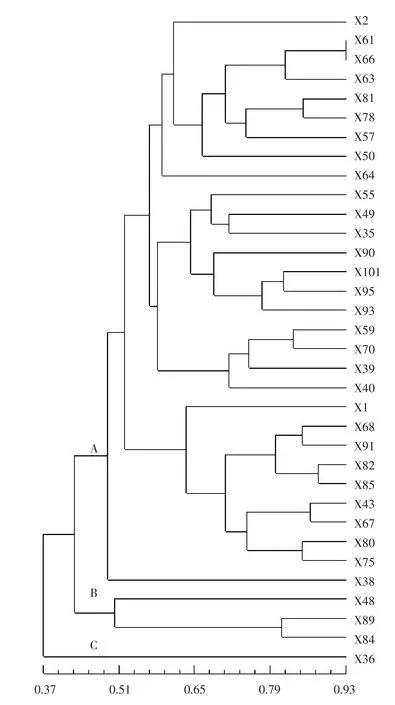
圖2 34份西瓜自交系的聚類結(jié)果
除B、C類之外的30份自交系全部聚集在A類,其平均相似系數(shù)高達0.78,說明該類材料遺傳多樣性不足,遺傳背景十分狹窄。A類中,X38可單獨作為一個亞類,其他29份材料分別可劃分為3個亞類,第1個亞類有9份材料(X2、X61、X66、X63、X81、X78、X57、X50、X64),第2個亞類有11份材料(X55、X49、X35、X90、X101、X95、X93、X59、X70、X39、X40),第3個亞類有9份材料(X1、X68、X91、X82、X85、X43、X67、X80、X75),每個亞類材料的來源地并不一致,其果實性狀也有一定的差異。A類中,除X38、X80外,其他單果質(zhì)量都大于或等于1.50 kg
3 討 論
本研究中自交系的可溶性固形物含量普遍比雜交種偏低,原因可能有幾點:1)自交系果實含糖量一般會低于雜交種;2)自交系果實中心至邊緣可溶性固形物含量梯度差異較大,其中間部位含量偏低;3)2010—2015年自交系材料均是露地種植,管理相對粗放,采收期為6月下旬至7月上旬,此時正是多雨季節(jié),土壤水分過高則降低了果實的可溶性固形物含量。
SSR標(biāo)記可分為基因組SSR和EST-SSR,前者來源于基因組的重復(fù)序列,一般通過構(gòu)建文庫、篩選、測序等步驟開發(fā),后者是基于cDNA文庫克隆(EST序列)的重復(fù)序列開發(fā)所得[16]。由于EST-SSR來源于轉(zhuǎn)錄組序列,其多態(tài)性很有可能與該物種的性狀有關(guān),因而被認(rèn)為是“絕對”的遺傳標(biāo)記,但其多態(tài)性一般低于基因組SSR。本研究結(jié)果表明,EST-SSR所檢測到的等位基因數(shù)和PIC值均低于SSR,與Fernandez-Silva等[17]的報道一致。Mondini等[18]認(rèn)為利用2種或2種以上的分子標(biāo)記進行基因型分析,可提高基因組覆蓋率,檢測更多的基因組變異。
基于2種分子標(biāo)記的研究結(jié)果表明,34份西瓜自交系的平均相似系數(shù)為0.56,大體可分為3類(A、B和C)。其中,B、C 2類包含4份自交系(X36、X48、X84和X89),其果實較小,均從中國新疆、甘肅等地的品種中自交分離所得,具有較高的遺傳多樣性。在主要植物學(xué)性狀上,這些小果型材料與當(dāng)?shù)氐牡胤狡贩N極為相似,可以作為育種材料增加栽培西瓜的遺傳多樣性,但其農(nóng)藝性狀(適應(yīng)性、抗病性等)有待進一步考察。
A類包含30份自交系,除X38外均為大果型材料,平均相似系數(shù)高達0.78,其遺傳背景十分狹窄。盡管A類存在3個明顯的亞類,但并不能依據(jù)材料的性狀和來源歸類,說明不同地區(qū)的材料之間可能存在基因交換。仔細(xì)分析發(fā)現(xiàn),來自美國的5份自交系(X43、X63、X75、X81和X91)和日本的2份自交系(X49和X95)均歸在A類,與國內(nèi)西瓜品種的自交系聚在一起,說明國內(nèi)外西瓜材料間可能有一定的血緣關(guān)系。中國曾在20世紀(jì)60年代大規(guī)模引進美國和日本的商業(yè)西瓜品種,如美國的‘Charleston Gray’‘Jubilee’‘Crimson Sweet’‘Sugar?lee’‘Sugar Baby’等,日本的主要是新大和、旭大和系列[3],這些品種及其衍生系被反復(fù)用作雜交親本,在中國西瓜的育種歷史上發(fā)揮了重要作用,可以說中國的西瓜品種具有很深的美國、日本品種的烙印。雖然34份自交系的系譜無法全部考證,但這些引種事實可以部分解釋國內(nèi)外材料緊密聚類在一起的原因。Levi等[19]曾報道非洲野生西瓜的遺傳多樣性極為豐富,且具有較強的抗病性、適應(yīng)性,可以用來拓寬栽培西瓜的遺傳背景。但這些野生材料的抗病或抗逆基因一般與品質(zhì)不利基因(如苦味、澀味等)連鎖,因此在應(yīng)用過程中應(yīng)注意利用輻射、化學(xué)誘變、多代回交等方法打破遺傳連鎖,加快種質(zhì)資源的創(chuàng)新與性狀改良進程。
[1]曹家樹,申書興.園藝植物育種學(xué)[M].北京:中國農(nóng)業(yè)大學(xué)出版社,2001.
[2]馬躍.透過國際分析,看中國西瓜甜瓜的現(xiàn)狀與未來[J].中國瓜菜,2011,24(2):64-67.
[3]劉文革.我國西瓜品種選育研究進展[J].中國瓜菜,2016,29(1):1-7.
[4]LEVI A,THOMAS C E,WEHNER T C,et al.Low genetic diversity indicates the need to broaden the genetic base of cultivated watermelon[J].HortScience,2001,36(6):1096-1101.
[5]LEVI A,THOMAS C E,KEINATH A P,et al.Genetic diversity among watermelon(Citrullus lanatusandCitrullus colocynthis)accessions[J].Genetic Resources Crop Evoution,2001,48(6):559-566.
[6]GUO S,ZHANG J,SUN H,et al.The draft genome of watermelon(Citrullus lanatus)and resequencing of 20 diverse accessions[J].Nature Genetics,2013,45(1):51-60.
[7]劉傳奇,高鵬,欒非時.西瓜遺傳陶譜構(gòu)建及果實相關(guān)性狀QTL分析[J].中國農(nóng)業(yè)科學(xué),2014,47(14):2814-2829.
[8]高磊,趙勝杰,路緒強,等.西瓜回交世代酸味株系基因型分析[J].中國瓜菜,2016,29(4):5-9.
[9]REDDY U K,NIMMAKAYALA P,LEVI A,et al.High-resolu?tion genetic map for understanding the effect of genome-wide recombination rate on nucleotide dilversity in watermelon[J].G3:Genes|Genomes|Genetics,2014,4(11):2219-2230.
[10]MURRAY M G,THOMPSON W F.Rapid isolation of high molecular weight plant DNA[J].Nucleic Acids Research,1980,8(19):4321-4325.
[11]JOOBEUR T,GUSMINI G,ZHANG X,et al.Construction of a watermelon BAC library and identification of SSRs anchored to melon orArabidopsisgenomes[J].Theoretical and Applied Genetics,2006,112(8):1553-1562.
[12]GUERRA-SANZ J M.Citrullussimple sequence repeats markers from sequence databases[J].Molecular Ecology Notes,2002,2(3):223-225.
[13]VERMA M,ARYA L.Development of EST-SSRs in watermelon(Citrullus lanatusvar.lanatus)and their transferability toCucumisspp.[J].Journal of Horticultural Science&Biotechnology,2008,83(6):732-736.
[14]JACCARD P.Nouvelles recherches sur la distribution florale[J].Bulletin De La Societe Vaudoise Des Sciences Naturelles,1908,44(163):223-270.
[15]BOTSTEIN D,WHITE R L,SKOLNICK M,et al.Construction of genetic linkage map in man using restriction fragment length polymorphisms[J].American Journal of Human Genetics,1980,32(3):314-331.
[16]KALIA R K,RAI M K,KALIA S,et al.Microsatellite markers:an overview of the recent progress in plants[J].Euphytica,2011,177(3):309-334.
[17]FERNANDEZ-SILVA I,EDUARDO I,BLANCA J,et al.Bin mapping of genomic and EST-derived SSRs in melon(Cucumis meloL.)[J].Theoretical and Applied Genetics,2008,118(1):139-150.
[18]MONDINI L,NOORANI A,PAGNOTTA M A.Assessing plant genetic diversity by molecular tools[J].Diversity,2009,1(1):19-35.
[19]LEVI A,THOMAS C E,KEINATH A P,et al.Estimation of genetic diversity amongCitrullusaccessions using RAPD markers [J].Hortscience,2000,35(3):385-390.
The use of SSR markers for analyzing the genetic relationship among watermelon inbred lines
XU Yanbin1,WANG Yanling1,WEN Xinhui2,HU Jianbin1,LI Qiong1,SUN Shouru1,MA Changsheng1
(1.College of Horticulture,Henan Agricultural University,Zhengzhou 450002,Henan,China;2.Seed Control Station of Pingluo Country,Pingluo 753400,Ningxia,China)
The present study aims to identify the relationship among the watermelon inbred lines by using SSR markers.Two markers,SSR and EST-SSR,were used to analyze the genotypes of the 34 inbred lines,which was adopted to calculate the similarity coefficient among the accessions and then cluster the accessions.The results showed that 9 genomic SSRs and 8 EST-SSRs detected 83 alleles totally,with a mean of 4.88.The PIC value ranged from 0.112 to 0.686,averaging 0.425.The polymorphism level of genomic SSR was higher than EST-SSR.All the materials were clustered into 3 groups(A,B and C). of the groups,B and C contained 4 accessions(X36,X48,X84,and X89)with high polymorphism,which were small fruit type accessions collected from northwest region.The 4 accessions could be used to increase the polymorphism of the cultivated watermelon.Group A contained the rest 30 domestic and foreign inbred lines which fruit weight no less than 1.5 kg with an exception of X38 and X80.These accessions had a low polymorphism and the genetic background need to be ex?panded further.Group A could be subdivided into 3 subgroups,each of which contained accessions with different origins and botanical characters.The results obtained here will offer theoretical basis for parent selection of watermelon cross breeding and new germplasm creation.
Watermelon;Inbred lines;SSR;Genetic relationship
2017-02-10;
:2017-03-14
國家農(nóng)業(yè)科技成果轉(zhuǎn)化基金項目(2014D00000103)
許彥賓,男,在讀碩士研究生,研究方向為西甜瓜遺傳育種。E-mail:xuyanb0514@163.com
胡建斌,男,教授,研究方向為西甜瓜遺傳育種。E-mail:jianbinhu@henau.edu.cn
