模擬氣候變暖對東北興安落葉松徑向生長的影響
2017-06-26 11:28:08劉玉佳顧卓欣王曉春
生態學報 2017年8期
關鍵詞:生長
劉玉佳,顧卓欣,王曉春
東北林業大學生態研究中心, 哈爾濱 150040
?
模擬氣候變暖對東北興安落葉松徑向生長的影響
劉玉佳,顧卓欣,王曉春*
東北林業大學生態研究中心, 哈爾濱 150040
2004年在帶嶺、孫吳、松嶺和塔河構成的緯度梯度上分別選取幼齡興安落葉松(Larixgmelinii),將其統一移栽到緯度較低的帽兒山地區,以模擬溫度升高對興安落葉松徑向生長的影響。在2013年生長季(6—9月)分別在4個緯度移栽原地和移栽地(帽兒山)的樹木主干和小枝上取樣,主干用微生長錐鉆取樣芯,小枝直接截取一段,主干和小枝的樣品用石蠟切片法得到擴大細胞和成熟細胞的大小及數量的動態變化。結果表明:溫度升高導致興安落葉松主干擴大細胞大小和數量均增加,但是主干徑向生長與溫度升高不呈正比。生長季前期(6月),帶嶺小枝成熟細胞數量明顯高于其他兩組,但在8月晚材細胞數量偏少,表明其還在繼續生長。生長季后期(8月),松嶺興安落葉松小枝擴大細胞平均大小為5 μm,分化數為4個,而松嶺移栽到帽兒山后擴大細胞平均大小為9.9 μm,分化數為7個,增長了近1倍。松嶺成熟細胞大小較其他兩地偏大,可能與其水分狀況有關。溫度升高不能直接改變植物生長周期一類由基因控制的生理特性,但是對樹木不同構件的影響力是不同的,小枝受到的影響要大于主干。當全球變暖尤其是伴隨降水量減少時,將減少興安落葉松徑向生長,包括管胞直徑、數量和生長周期。通過緯度變化模擬氣候變暖,深入分析不同構件樹木生長對氣候變化的響應,為預測氣候變化對興安落葉松生長及分布的影響提供依據。
興安落葉松;年輪;暖化;形成層;切片
近幾十年以來,人類活動引起溫室氣體排量增加,致使地表溫度升高,導致全球持續變暖[1],模型預測未來升溫可能更為明顯[2]。預計到2100年,全球地表溫度將相對1980—1999年的水平升高1.1—6.4℃[3],而我國溫度將升高3.9—6.0℃[4]。北部高緯度預計將有更大幅度的變暖[5-6],全球升溫對高緯度地區影響顯著[1,7],這可能導致高緯度寒溫帶森林面積大幅度下降[7-8]。興安落葉松(Larixgmelinii)林是我國北方森林的優勢植被類型[9],在大興安嶺地區占到70%以上[10],其碳儲量約占全國森林總碳儲量的8.5%[11]。據張新時[12]、程肖俠等[13]預測,溫度、降水量增加到一定程度時,興安落葉松林可能向北大幅移動。因此,探索興安落葉松對全球變暖的響應具有重要意義。
在樹木徑向生長與氣候變化的響應關系上,溫度一直是一個主要研究因子。有些樹木年輪寬度與生長季前后的溫度呈負相關,即溫度越高徑向生長量越少[14],還有一些樹木剛好與之相反[15]。這樣當溫度變化時,不同樹木的響應就會出現差異[16-17]。對不同時間段的溫度變化,徑向生長也有不同響應,5—8月的溫度升高可以促進山楊(Populusdavidiana)的徑向生長[18]。在樹木徑向生長過程中,管胞的一些形態特征同年輪寬度一樣也傳遞著重要氣候信息[19]。管腔尺寸和氣候波動有緊密的聯系[20],2—3月的平均溫度升高將有利于管胞直徑的變大[21]。管胞直徑、數量、管胞壁厚等一些形態特征存在著較大的差異性[22-23],在氣候變化后,管胞的數量、大小、直徑、壁厚等特征不可能同時增加[24]。在每年早材形成的5—7月,溫度對其影響較高。由于早材細胞形成于春季,春季溫度的適度提升有利于形成層活動,可形成較大、較多的管胞。入秋后,溫度下降,形成層活動趨于停止,形成的管胞較小、較少[25-26]。在對北美云杉(Piceasitchensis)的研究中發現,管胞數量和直徑與日平均溫度有著明顯的相關關系[27]。在徑向生長的形成層活動中,形成層活動對于溫度的響應存在一定的差異,在植物休眠階段通過局部加熱發現,形成層活化區僅為加熱區域[28],形成層開始活動后逐漸分化出早材細胞,如果降低溫度,對形成層影響較小,主要是晚材比例下降[29]。
本研究通過將不同緯度梯度的興安落葉松幼苗,移植到其分布區南緣的帽兒山森林生態系統國家野外觀測研究站,模擬升溫對興安落葉松的徑向生長(包括年輪寬度、莖和枝的木質部管胞特征及形成層分化情況)的影響,以探討:(1)不同程度的溫度升高對興安落葉松徑向生長有什么樣的影響,影響是否有差別?(2)不同程度的升溫給興安落葉松徑向生長帶來的影響在不同部位是否有差別?(3)興安落葉松移栽前后的同一時期,徑向生長是否存在差別?
1 材料與方法
1.1 研究區域概況
本研究將實驗地分為兩個部分,一部分為移栽原地,另一部分為移栽地。在興安落葉松自然分布區內由南向北選擇4個實驗點:帶嶺、孫吳、松嶺和塔河,構建1條主要由溫度驅動的緯度梯度樣帶(表1)。于2004年秋季,根據其起源、樹齡和經營方式等選取上述4個地點的興安落葉松幼苗、地被物和土壤A、B層,移植到其分布區南緣的帽兒山森林生態系統國家野外觀測研究站實驗地,構成帶嶺、孫吳、松嶺和塔河4個實驗處理[29]。
實驗地采用隨機設計,每個處理設置4塊重復樣地,每塊樣地面積為10 m×10 m,移栽木行距為1.5 m[29]。
1.2 樣品采集
2013年生長季(6—9月),在移栽原地(帶嶺、孫吳、松嶺和塔河)和移栽地(帽兒山)分別隨機選取3棵樣樹,用微生長錐在每棵樣樹上取3個樣芯并放入福爾馬林-乙酸-乙醇(FAA)固定液中固定。每棵樣樹上再分別取3個5 cm長小枝同樣放入FAA固定液中固定。固定好的樣品后帶回實驗室后將樣芯放入70%乙醇溶液中,隨后再以5%乙醇濃度遞增直至100%的乙醇溶液脫水浸蠟包埋,隨后對包埋好樹芯進行石蠟切片,樣芯切片后對木質化和非木質化部分主要用染色劑來區分,番紅可以將木質化部分染成紅色,固綠將非木質化部分染成藍綠色。用光學顯微鏡觀察切片封好的片子,通過測量細胞形成層擴大細胞的尺寸,結合樣地處理后的氣候數據進行分析。在時間尺度上通過觀察分化細胞的變化,了解木質部形成的季節動態變化。在生長季結束時,用生長錐在塔河、松嶺、孫吳和帶嶺的移栽地(帽兒山)取樣,將樹芯帶回室內實驗室后打磨,在光學顯微鏡下觀察成熟管胞的徑向長度(以下簡稱管胞直徑)及數量。
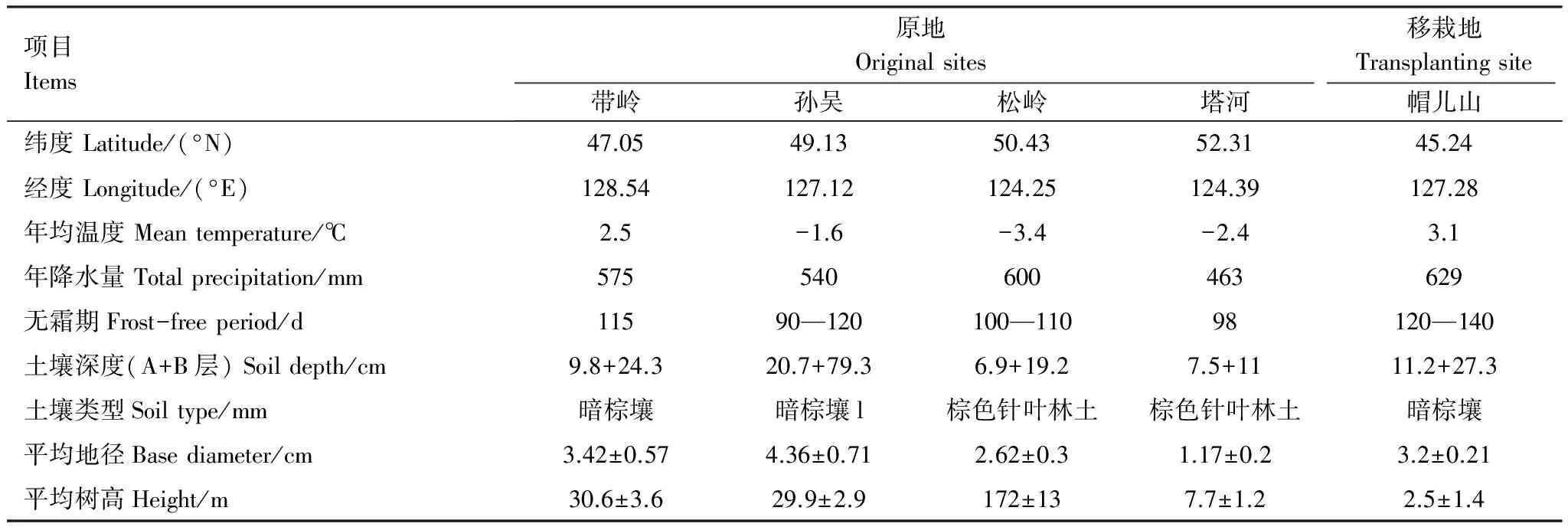
表1 興安落葉松移栽原地及移栽地的生境基本特征[29]
1.3 統計分析
采用單因素方差分析統計了2013年移栽原地和移栽地樹干和小枝之間擴大細胞和成熟細胞大小、數量以及管胞直徑、數量,用多重比較來分析每個移栽原地之間的差別和移栽后的差別。
2 結果與分析
2.1 移栽原地自然溫度梯度下興安落葉松徑向生長的差異
溫度升高對興安落葉松徑向生長有一定促進作用(圖1)。4個移栽原地構成的緯度梯度中,緯度較低的帶嶺和孫吳的平均年輪寬度顯著高于緯度較高的松嶺和塔河。從時間序列上看,緯度最高的塔河每年徑向生長量波動不大,每年徑向生長量基本上在4個移栽原地中都是最低的。緯度最低的帶嶺每年的徑向生長量都相對較高,而孫吳在個別年份徑向生長量突出。
在2013年生長季末,將當年的年輪樹芯打磨光滑后在顯微鏡下測得管胞直徑并且按著由早材向晚材方向排列(圖2)。在生長季初期,4個移栽原地中塔河的管胞直徑明顯低于其它3個移栽原地,帶嶺、孫吳和松嶺3個移栽原地之間沒有明顯差別。在生長季后期的晚材管胞直徑上,可以看出帶嶺管胞的直徑要高于其它移栽原地的管胞直徑,塔河移栽原地依然是各原地中最低的。由此,管胞的直徑與緯度相反,緯度越高,興安落葉松管胞直徑越小。從管胞數量看,帶嶺在生長季階段數量最多,然后隨緯度的升高而遞減,松嶺的管胞數量比孫吳略低,緯度最高的塔河管胞數量是4個移栽原地中最少的,生長期也是最短的。溫度差異導致帶嶺興安落葉松管胞直徑和數量均最高,塔河的均最低。
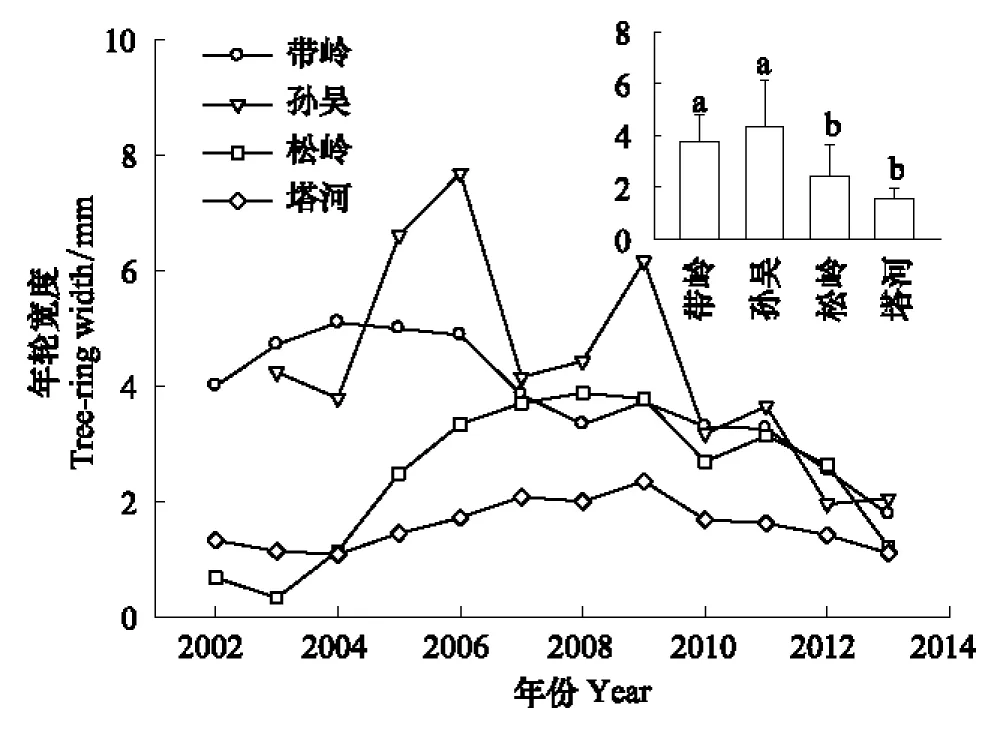
圖1 帶嶺、孫吳、松嶺和塔河移栽原地年輪寬度比較 Fig.1 Comparison of tree-ring width of Larix gmelinii in Dailing, Sunwu, Songling and Tahe
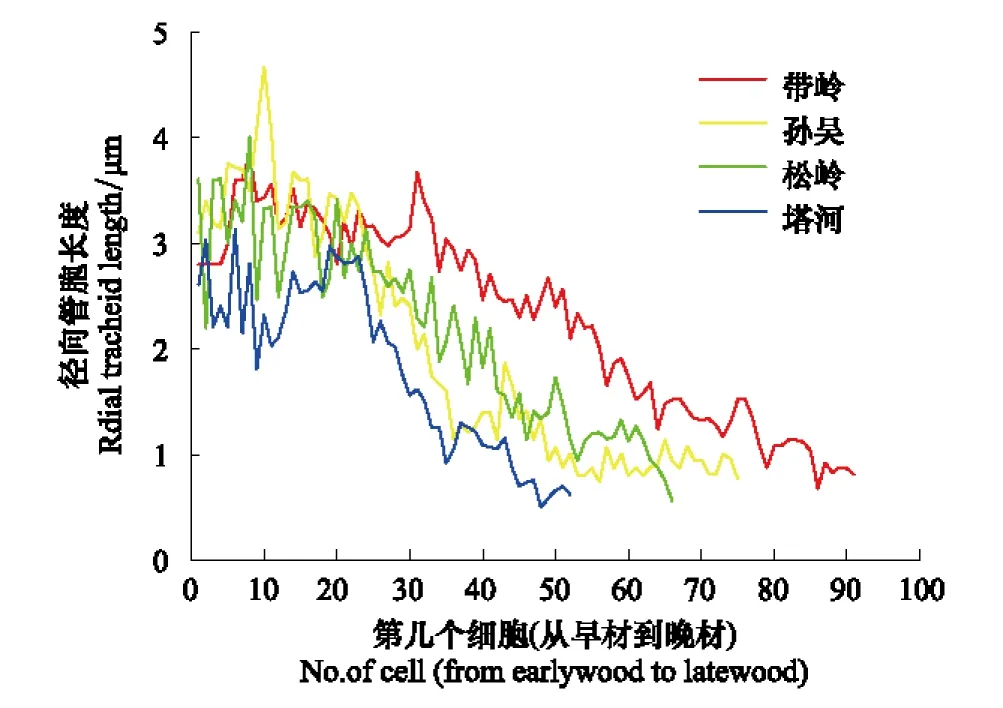
圖2 帶嶺、孫吳、松嶺和塔河移栽原地徑向管胞直徑從早材到晚材變化比較Fig.2 Comparison of radial tracheid length from the first earlywood cell to the last latewood cell at Dailing, Sunwu, Songling and Tahe
2.2 模擬升溫對興安落葉松主干形成層擴大細胞的影響
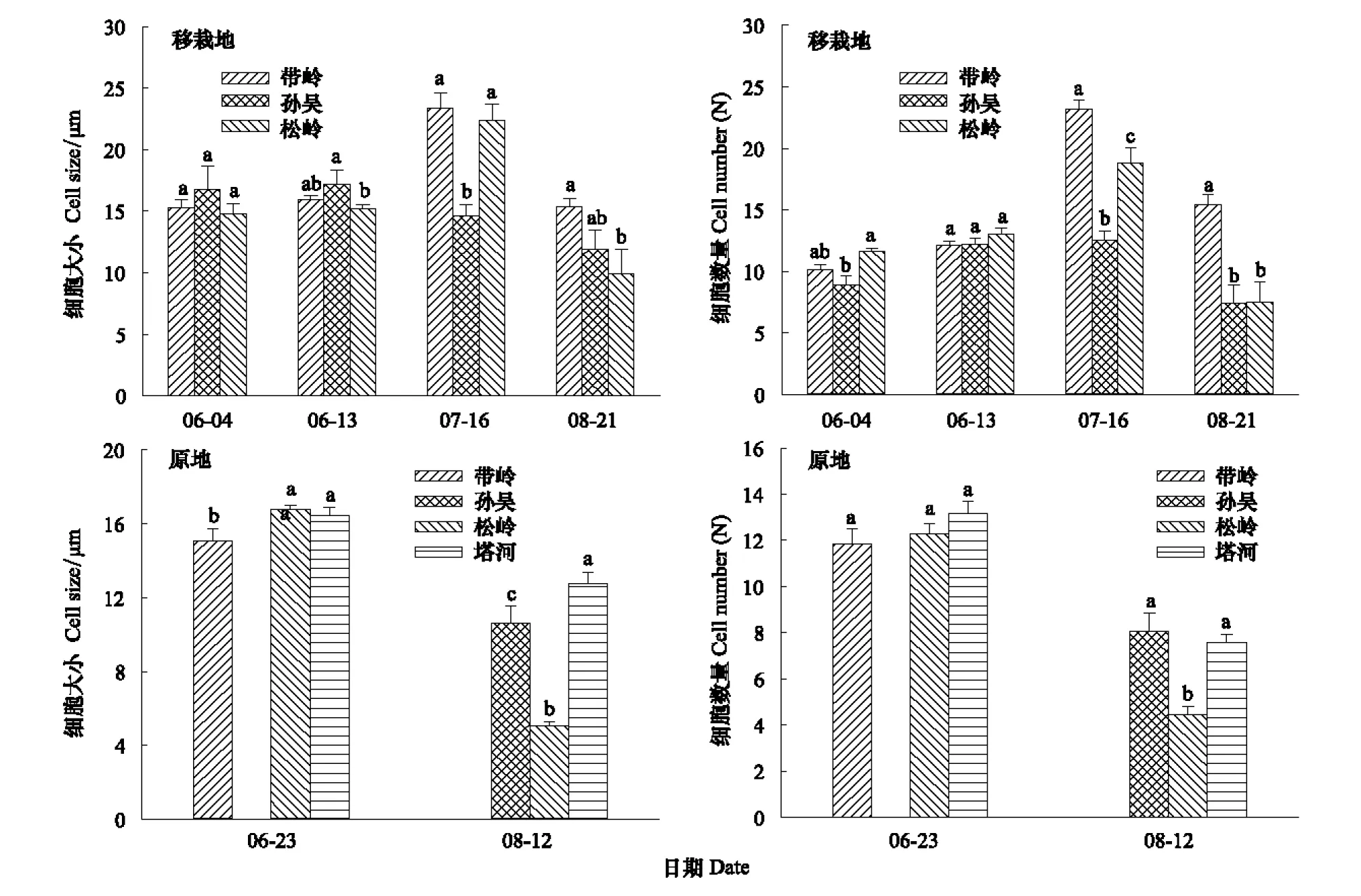
圖3 在生長季不同階段移栽地和原地微生長芯擴大細胞數量及大小比較Fig.3 Comparison of the expanded cell number and size at different time of growing season for original and transplanting sites
帽兒山地區移栽木從分化細胞的數量上看(圖3),6月初,孫吳擴大細胞數量相對較少,擴大細胞大小略大。整體看,3個地點移栽木在擴大細胞大小上沒有顯著差別,而擴大細胞數量則是松嶺移栽木顯著高于孫吳移栽木。6月中旬,3組移栽木的擴大細胞數量沒有顯著差別,擴大細胞大小則是孫吳顯著高于松嶺,并且達到擴大細胞大小的峰值。7月中旬,3組移栽木的擴大細胞數量均達到峰值,帶嶺顯著高于其它兩組,孫吳擴大細胞數量最少。帶嶺和松嶺移栽木的擴大細胞大小也在7月達到峰值,均顯著高于孫吳擴大細胞。8月下旬,移栽木擴大細胞數量與大小的趨勢相同,帶嶺移栽木擴大細胞數量和大小均為最高,顯著高于其它地區的移栽木(圖3)。8月份的這一趨勢也與移栽木之前原地的緯度相一致,即原地緯度越高停止生長越早。綜合移栽木擴大細胞大小和數量分析,帶嶺移栽木在7月出現峰值,且均高于其它兩地的移栽木;孫吳移栽木變化曲線幅度不大;松嶺移栽木則是在生長季階段細胞分化最早的移栽木,但是在8月和孫吳移栽木開始一起逐漸停止生長,時間上要早于帶嶺的移栽木(圖3)。
由移栽原地與移栽地興安落葉松形成層擴大細胞活動可知,松嶺地區興安落葉松與移栽地一樣在6月初期也開始分化細胞開始生長,而在8月份的時候逐漸停止分化;雖然這一習性整體上并沒有因為溫度的升高而改變,但是在細節上二者存在差異較大。松嶺原地在8月份時,分化的擴大細胞平均大小為5 μm,分化數量為4。而移栽地在8月份時擴大細胞平均大小為9.9 μm,分化數量為7,增長了近一倍;說明溫度升高雖然不能改變松嶺地區興安落葉松的生活習性,但是可以影響各個發育階段的生長幅度。8月份原地取樣,擴大細胞均值最小的為松嶺,主要原因是切片中松嶺幾乎全部都是接近停止生長的狀態,而其次的孫吳出現了部分接近停止分化的樣本。擴大細胞在數量上松嶺也是移栽原地樣樹中最少的(表2)。

表2 原地與移栽地樹干擴大細胞數量和大小比較
*表示移栽地和原地差異顯著(P< 0.05);M:移栽地Transplanting site,Y:原地Original sites
2.3 模擬升溫對興安落葉松小枝形成層擴大與成熟細胞的影響
從不同緯度移栽至帽兒山的興安落葉松小枝(2—3年生)細胞大小可以看出(圖4),6月初,松嶺移栽木的擴大細胞大小略大于其它兩地的移栽木,但不顯著。6月中旬,松嶺移栽木的擴大細胞大小顯著高于其它兩地的移栽木。7月份,3組移栽木的擴大細胞大小差異不顯著,孫吳移栽木擴大細胞的大小達到峰值。8月份,孫吳移栽木的擴大細胞大小顯著低于其它兩地的移栽木。帶嶺和松嶺達到擴大細胞大小的峰值。9月份3地的移栽木差異不顯著,但是孫吳還是略低的。孫吳移栽木的生長時期相對較短,帶嶺的增長速率則是最高的。
從擴大細胞數量上看(圖4),6月份,孫吳移栽木顯著小于其它地點的移栽木。7月份,3個地點的移栽木細胞數量沒有顯著差異,均達到峰值。8月份孫吳移栽木的細胞數量顯著低于其它兩地的移栽木。9月份,3個地點的移栽木在小枝擴大細胞數量上沒有顯著差異。綜合不同地點移栽木小枝擴大細胞大小和數量比較,可以得出孫吳小枝的生長時間最短,松嶺略低于帶嶺。
從移栽木小枝成熟細胞大小上看(圖4),松嶺移栽木的成熟細胞明顯大于其它兩地,6月份的成熟細胞中全部都是早材細胞,早材多與水分相關,水分充足時管胞直徑較大,因為松嶺相比其它兩地濕度差最大,導致早材形成的管胞較大。8月份,松嶺移栽木成熟細胞大小明顯低于其它兩地。8月份已經開始出現晚材細胞,這樣成熟細胞大小的均值被拉低,晚材細胞數量要高于其它兩地,接近停止生長,導致細胞大小均值上顯著低于其它地區。
從移栽木小枝成熟細胞數量上看(圖4),帶嶺地區要高于其它兩組,說明在小枝活動中帶嶺移栽木優先開始生長活動。在隨后月份中,數量都是逐漸增加的,但是增加速度最快的是松嶺移栽木,它在6月份活動相對較晚,但是從7月份開始就在數量上領先,并在8月份顯著高于其它組。結合成熟細胞大小,可以發現松嶺移栽木在8月份時數量明顯超過帶嶺,但是在平均大小上卻略小。所以在8月份時孫吳和松嶺移栽木都出現大量晚材細胞,相比較而言,帶嶺移栽木晚材細胞最少,生長時期更長。
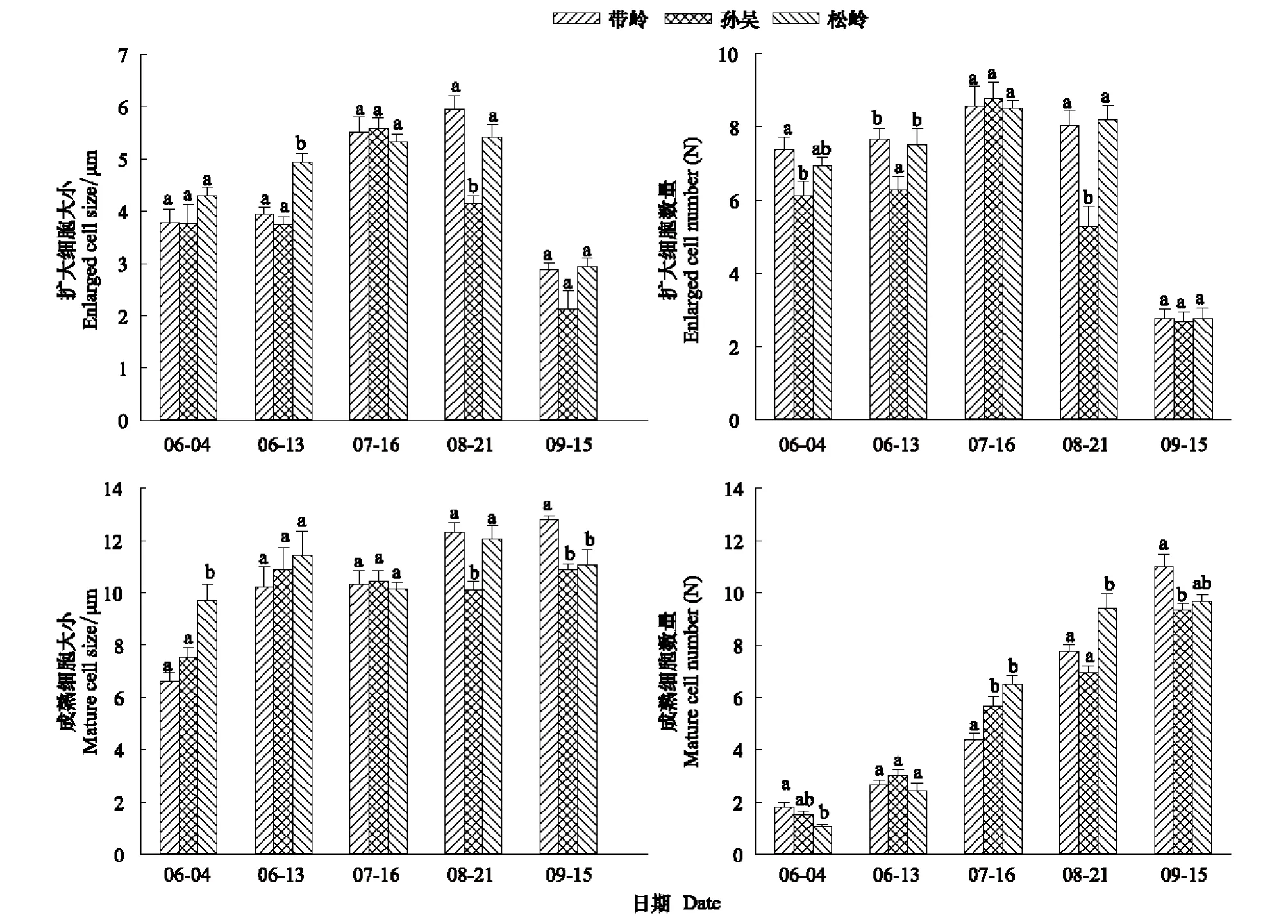
圖4 移栽地興安落葉松小枝擴大、成熟細胞大小及數量比較Fig.4 Comparisons of the enlarged and mature cell size and number at braches of Larix gmelinii from different transplanting sites
6月份的小枝在移栽原地中,帶嶺的擴大細胞大小明顯高于其它移栽原地(圖5),數量上帶嶺和松嶺明顯高于塔河(圖5)。而6月份的各個移栽原地中成熟細胞的大小無顯著差異(圖5),數量上看,帶嶺最高,塔河最低(圖5)。在生長季初期,興安落葉松的小枝徑向生長量與緯度分布相吻合,即溫度在生長季初期小枝的徑向生長中是關鍵因子。在不同緯度的移栽原地中,小枝成熟細胞的大小都沒有顯著差異,緯度較低的帶嶺地區,成熟細胞的數量在生長季初期、中期和晚期都顯著高于其它移栽原地。此外,帶嶺地區的擴大細胞大小和數量在生長季中期明顯高于其它移栽原地。
從移栽地和原地小枝細胞數量和大小比較可以看出(表3),溫度升高有利于興安落葉松小枝的徑向生長。但是移栽地中帶嶺移栽木的小枝并沒有因為溫度升高而增加細胞數量和大小,相反在生長季初期,帶嶺成熟細胞大小上甚至明顯低于原地和其它地區的移栽木。在生長季末,帶嶺移栽木小枝成熟細胞數量比原地減少了近1/3。在松嶺和帶嶺地區在生長季初的6月,其擴大細胞的數量和大小以及成熟細胞的數量和大小沒有差異,表明與移栽后的小枝在帽兒山有共同的生長軌跡,以表示對移栽地的適應。
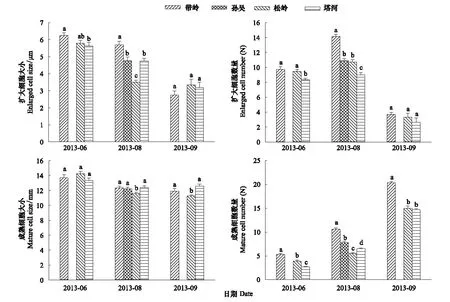
圖5 原地興安落葉松小枝擴大、成熟細胞大小及數量比較Fig.5 Comparisons of the enlarged and mature cell size and number at braches of Larix gmelinii from original sites
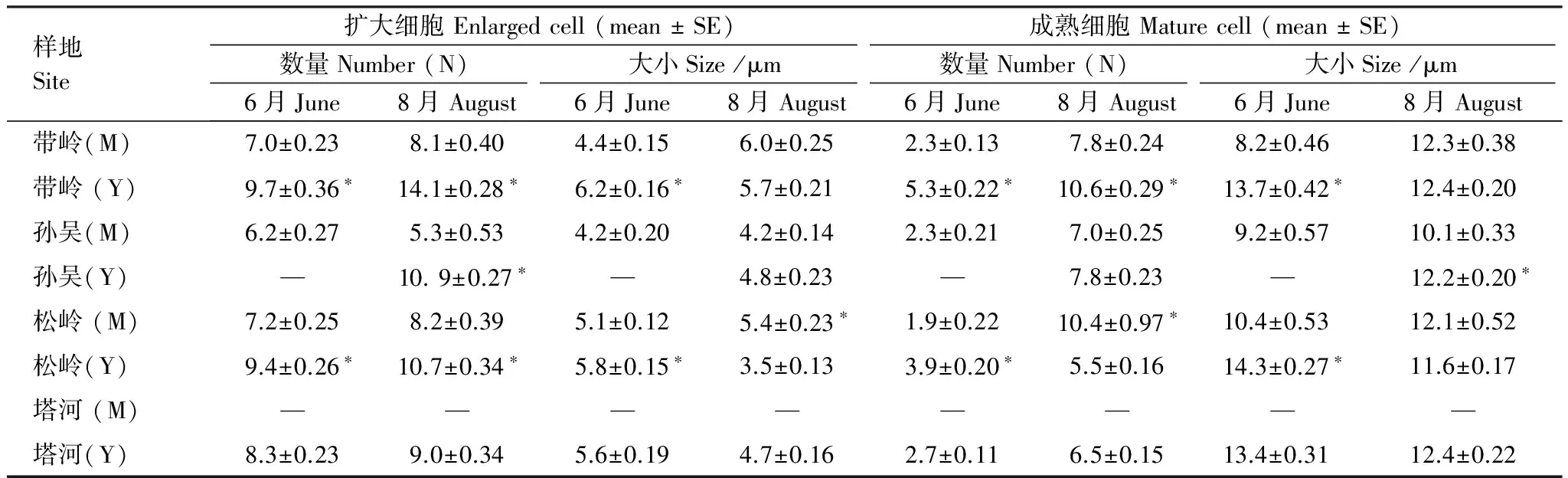
表3 原地與移栽地小枝擴大、成熟細胞數量和大小比較
*表示移栽地和原地差異顯著(P< 0.05);M:移栽地Transplanting site,Y:原地Original sites
3 討論
樹木年輪是研究樹木生長與氣候關系的重要途徑,具有定年準確、連續性強、分辨率高、易于獲取和復本量大等優點,已被廣泛用于全球變化以及古氣候研究[15- 26,30- 33]。樹輪解剖特征是反映樹木生長的一種更高分辨率代用指標,也是樹輪在細胞尺度上的分量。已被廣泛用于全球變化研究[19- 24,34-35]。近年來,東北地區溫度顯著升高[30,31]。在寒冷環境中生長的樹木,木質部是由溫度主導代謝活動[32],因此溫度變化影響和改變樹木的年輪特征[33]。隨緯度梯度變化的主要氣候因子是溫度[35],不同緯度移栽模擬了溫度升高對興安落葉松徑向生長的影響。移栽后,孫吳的年輪寬度超過低緯度的帶嶺,表明增溫幅度不同對興安落葉松徑向生長的影響也存在差異。樹輪對環境變化的記錄上除了寬度外,樹輪解剖結構也記錄不少的生態環境信息[34, 36]。樹輪解剖結構相對于樹輪寬度長期對氣候的響應,更能展現短期內氣候變化對樹木徑向生長的影響。2013年興安落葉松的管胞大小及數量在不同緯度地區有明顯的區分,帶嶺管胞直徑相對較大,管胞數量明顯多于其它地區,這導致帶嶺的徑向生長量較大。塔河年均溫度比帶嶺低約6℃,年均降水量也是4個移栽原地中最少的,其管胞大小和樹輪明顯低于其它地區,導致塔河地區的徑向增量最小。
樹木生長發育過程受到外部因素和內部因素的控制,外部因素主要有溫度、降水和日照等。物候觀測、遙感數據等證實了溫度升高提早樹木展葉期、延遲落葉期,進而延長了植物的生長期[37-38],而生長期的延長會顯著影響樹木的徑向生長[39]。高緯度地區樹木形成層的活動主要依賴夏季溫度,在低溫時期提升溫度,可以促進形成層開始活動[28]。移栽后,松嶺興安落葉松樹干生長時期延長,導致生長期內形成層分化的細胞數量和大小增加,進而使徑向生長量增加,Vaganov等[40]也發現北方樹線附近的落葉松管胞直徑會因長期的夏季升溫而顯著增加。移栽原地中,帶嶺緯度最低,在6月份,移栽后樹干擴大細胞大小并未顯著增加,細胞數量也有減少趨勢。這表明溫度在一定閾值內增加有利于樹木的徑向生長,超過這個閾值繼續增加可能減少樹木徑向[41]。不同緯度樣地落葉松南移(升溫)后,徑向生長上表現一定程度上增加趨勢,但模擬升溫的強度與相應的生長量或形成層活動并不成正比,這可能是移栽(升溫)后控制落葉松徑向生長的限制因子發生轉變的結果[19,42];不同緯度興安落葉松南移后,溫度對落葉松的限制作用相對減弱,其他的環境因子如升溫所引起的暖干旱可能會使得水分取代溫度成為新的主控因子。已有研究表明,東北升溫在一定程度上促進了樹木徑向生長,但是近幾十年徑向增長的速度有所減緩,這可能與溫度的閾值效應以及升溫帶來的干旱脅迫有關[30,43]。高緯度地區,樹木生長(或新細胞產生)約于8月中旬停止,生長季長度主要由形成層活動的開始日期決定[44];相反,本研究則發現移栽地(升溫)與原地興安落葉松莖干形成層擴大細胞的數量及大小,在生長季初期差異不顯著,而生長季末期差異顯著,表明升溫可能會以推遲生長季結束時間的方式延長生長季。
研究不同功能型植物的小枝生長特征,可以更好地理解植物功能性狀隨環境變化發生的一些改變[45]。在生理上許多復雜的器官有單獨的代謝機能,小枝可以看作是樹木生長的獨立器官,具有一定的獨立性[46],是樹木最有活力的部分之一[47]。小枝的次級組織較少,在內部能很快的進行營養物質和碳水化合物流動和轉移,相比整株樹木對環境的響應更顯著或者具有與主干不同的響應特征[48]。移栽前,在生長季初期,原地小枝擴大細胞大小與溫度呈正比,即隨緯度升高,溫度降低,擴大細胞大小降低;在生長季末期,小枝成熟細胞數量也與溫度成正比,帶嶺成熟細胞數量是最多的。移栽到低緯度地區后,松嶺小枝成熟細胞大小高于其它地點的移栽木,但在成熟細胞數量上的優勢不明顯。孫吳小枝成熟和擴大細胞都略低于來自松嶺的移栽木,可能是由于移栽后帽兒山水分高于原地所致。在生長季中,小枝擴大細胞大小和數量差異都不顯著,說明小枝的徑向生長與溫度的升高不成正比。但在生長季初期的成熟細胞數量可以看出移栽木形成層活動的起始時間還是略有不同。相對樹木的莖干而言,移栽后興安落葉松的小枝木質部解剖特征相比主莖變化更為顯著,小枝對氣候變化(升溫)響應更為敏感,樹木小枝的解剖特征相比主干更適合被用于研究樹木生長與氣候關系的變化。升溫后小枝擴大細胞數量明顯減少(表3),表明升溫后,興安落葉松的生長策略已由強調多細胞同時分化的數量速度取勝模式向強調分化持續時間的時間取勝模式轉變[42]。
年輪中記載了大量的信息,是眾多環境因子在不同時間段內綜合作用結果,很難確定某一特定環境因子在特定時期內對樹木生長的單獨影響[49]。一般認為,溫度是限制高緯度地區樹木生長的關鍵因子,也是影響寒帶或溫帶森林樹木形成層活動或解剖特征重要因素[42]。雖然移栽原地落葉松樹輪寬度并未隨溫度梯度而呈現出線性的上升或下降的趨勢,但整體表現出高緯度年輪較窄,低緯度年輪較寬的特征(圖1),這也從側面印證了緯度梯度上溫度的限制作用。孫吳地區落葉松輪寬整體上高于帶嶺地區;一方面,可能是由于該地區土層相對較深(A+B層厚為(20.7+79.3) cm),能為落葉松生長提供更多的養分;另一方面,異常充沛的降水也為落葉松生長提供必要的水分條件;如2004、2005年孫吳地區生長季初期(4—5月)的降水量分別為104.7、158 mm遠高于近幾十年來均值(69 mm),充沛的降水提早落葉松的形成層活動并加速細胞的分裂與分化,進而形成寬輪;2009年充沛的生長季(5—8月,530 mm近似年平均540 mm)降水也有可能是導致該年生長量較高的原因。此外,種間或種內關系等也可能是產生以上差異的部分原因。
4 結論
本研究通過將帶嶺、孫吳、松嶺和塔河4個不同緯度的興安落葉松幼苗南移至帽兒山地區,來模擬氣候變暖對興安落葉松年輪寬度以及莖、枝木質部解剖特征的影響。溫度升高在一定程度上有利于興安落葉松的徑向生長,但是溫度升高與生長量的增加不成正比;降水、種間或種內關系等溫度以外干擾因素的影響可能是產生上述現象的原因。不同緯度興安落葉松南移(升溫)后,溫度對落葉松的限制作用相對減弱,水分對樹木生長的限制作用增強。南移(升溫)后,興安落葉松的生長策略已由強調多細胞同時分化的數量取勝模式向強調分化持續時間的時間取勝模式轉變。升溫使興安落葉松的生長季結束延遲,間接延長了生長季長度。興安落葉松小枝解剖特征相比主干解剖特征對升溫響應更敏感,小枝相比主干而言更適合被用于氣候變化研究。
[1] IPCC. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK: Cambridge University Press, 2007.
[2] Marotzke J, Forster P M. Forcing, feedback and internal variability in global temperature trends. Nature, 2015, 517(7536): 565- 570.
[3] Hansen J, Ruedy R, Sato M, Reynolds R. Global surface air temperature in 1995: return to pre-Pinatubo level. Geophysical Research Letters, 1996, 23(13): 1665- 1668.
[4] 《氣候變化國家評估報告》編寫委員會. 氣候變化國家評估報告. 北京: 科學出版社, 2007.
[5] Bonsal B R, Zhang X, Vincent L A, Hogg W D. Characteristics of daily and extreme temperatures over Canada. Journal of Climate, 2001, 14(9): 1959- 1976.
[6] Vincent L A, Mekis E. Changes in daily and extreme temperature and precipitation indices for Canada over the twentieth century. Atmosphere-Ocean, 2006, 44(2): 177- 193.
[7] Krankina O N, Dixon R K. Forest management options to conserve and sequester terrestrial carbon in the Russian Federation. World Resource Review, 1994, 6(1): 88- 101.
[8] 周廣勝, 王玉輝, 張新時. 中國植被及生態系統對全球變化反應的研究與展望. 中國科學院院刊, 1999, 14(1): 28- 32.
[9] 周以良, 董世林, 聶紹荃. 黑龍江樹木志. 哈爾濱: 黑龍江科學出版社, 1986.
[10] 徐化成. 中國大興安嶺森林. 北京: 科學出版社, 1998.
[11] 周玉榮, 于振良, 趙士洞. 我國主要森林生態系統碳貯量和碳平衡. 植物生態學報, 2000, 24(5): 518- 522.
[12] 張新時. 研究全球變化的植被-氣候分類系統. 第四紀研究, 1993, 13(2): 157- 169.
[13] 程肖俠. 氣候變化背景下中國東北森林的演替動態[D]. 北京: 中國科學院研究生院(大氣物理研究所), 2007.
[14] 何維明, 董鳴. 升高氣溫對旱柳光合和生長的影響. 林業科學, 2003, 39(1): 160- 164.
[15] 劉春延, 谷建才, 李吉躍, 陳平, 陸貴巧, 田國恒. 塞罕壩華北落葉松生長與氣候因子的相關分析. 北京林業大學學報, 2009, 31(4): 102- 105.
[16] 熊偉, 王彥輝, 于澎濤, 劉海龍, 時忠杰, 管偉. 六盤山南坡華北落葉松(Larixprincipis-rupprechtii) 樹干直徑生長及其對氣象因子的響應. 生態學報, 2007, 27(2): 432- 441.
[17] 張先亮, 何興元, 陳振舉, 崔明星, 黎娜. 大興安嶺山地樟子松徑向生長對氣候變暖的響應——以滿歸地區為例. 應用生態學報, 2011, 22(12): 3101- 3108.
[18] 王雨茜, 高露雙, 趙秀海. 長白山山楊年輪年表及其與氣候變化的關系. 東北林業大學學報, 2013, 41(1): 10- 13.
[19] Olano J M, Eugenio M, García-Cervigón A I, Folch M, Rozas V. Quantitative tracheid anatomy reveals a complex environmental control of wood structure in continental Mediterranean climate. International Journal of Plant Sciences, 2012, 173(2): 137- 149.
[20] Carrer M, von Arx G, Castagneri D, Petit G. Distilling allometric and environmental information from time series of conduit size: the standardization issue and its relationship to tree hydraulic architecture. Tree Physiology, 2015, 35(1): 27- 33.
[21] DeSoto L, De la Cruz M, Fonti P. Intra-annual patterns of tracheid size in the Mediterranean treeJuniperusthuriferaas an indicator of seasonal water stress. Canadian Journal of Forest Research, 2011, 41(6): 1280- 1294.
[22] 趙西平, 郭明輝. 落葉松管胞形態與氣候變化關系的實證分析. 東北林業大學學報, 2009, 37(7): 45- 48.
[23] Deslauriers A, Morin H. Intra-annual tracheid production in balsam fir stems and the effect of meteorological variables. Trees, 2005, 19(4): 402- 408.
[24] 趙西平, 郭明輝. 人工林紅松管胞形態對氣候變化的響應. 東北林業大學學報, 2009, 37(8): 1- 4.
[25] 高洪娜, 高瑞馨. 氣象因子對樹木生長量影響研究綜述. 森林工程, 2014, 30(2): 6- 9.
[26] M?kinen H, Njd P, Saranp?? P. Seasonal changes in stem radius and production of new tracheids in Norway spruce. Tree Physiology, 2003, 23(14): 959- 968.
[27] Barnett J R, Miller H. The effect of applied heat on graft union formation in dormantPiceasitchensis(Bong.) Carr. Journal of Experimental Botany, 1994, 45(1): 135- 143.
[29] 霍宏. 氣候暖化對興安落葉松光合和生長影響的研究[D]. 哈爾濱: 東北林業大學, 2007.
[30] 朱良軍, 楊婧雯, 朱辰, 王曉春. 林隙干擾和升溫對小興安嶺紅松和臭冷杉徑向生長的影響. 生態學雜志, 2015, 34(8): 2085- 2095.
[31] 劉實, 閆敏華, 隋波. 東北三省冬季氣溫變化的有關研究進展. 氣候變化研究進展, 2009, 5(6): 357- 361.
[32] Rossi S, Morin H, Deslauriers A, Plourde P Y. Predicting xylem phenology in black spruce under climate warming. Global Change Biology, 2011, 17(1): 614- 625.
[33] 封曉輝, 程瑞梅, 肖文發, 王瑞麗, 王曉榮, 劉澤彬. 基于日均溫度的華山松徑向生長敏感溫度研究. 生態學報, 2012, 32(5): 1450- 1457.
[34] Fonti P, Von Arx G, García-González I, Eilmann B, Sass-Klaassen U, G?rtner H, Eckstein D. Studying global change through investigation of the plastic responses of xylem anatomy in tree rings. New Phytologist, 2010, 185(1): 42- 53.
[35] 方精云. 地理要素對我國溫度分布影響的數量評價. 生態學報, 1992, 12(2): 97- 104.
[36] Chave J, Coomes D, Jansen S, Lewis S L, Swenson N G, Zanne A E. Towards a worldwide wood economics spectrum. Ecology letters, 2009, 12(4): 351- 366.
[37] Zheng J Y, Ge Q S, Hao Z X, Wang W C. Spring phenophases in recent decades over eastern china and its possible link to climate changes. Climatic Change, 2006, 77(3): 449- 462.
[38] Chen X Q, Hu B, Yu R. Spatial and temporal variation of phenological growing season and climate change impacts in temperate eastern China. Global Change Biology, 2005, 11(7): 1118- 1130.
[39] Hasenauer H, Nemani R R, Schadauer K, Running S W. Forest growth response to changing climate between 1961 and 1990 in Austria. Forest Ecology and Management, 1999, 122(3): 209- 219.
[40] Vaganov E A, Hughes M K, Shashkin A V. Growth Dynamics of Conifer Tree Rings. Berlin Heidelberg: Springer, 2006.
[41] Drake J E, Aspinwall M J, Pfautsch S, Rymer P D, Reich P B, Smith R A, Crous K Y, Tissue D T, Ghannoum O, Tjoelker M G. The capacity to cope with climate warming declines from temperate to tropical latitudes in two widely distributedEucalyptusspecies. Global Change Biology, 2015, 21(1): 459- 472.
[42] Vaganov E A, Anchukaitis K J, Evans M N. How well understood are the processes that create dendroclimatic records? A mechanistic model of the climatic control on conifer tree-ring growth dynamics // Hughes M K, Swetnam T W, Diaz H F, eds. Dendroclimatology. Netherlands: Springer, 2011: 11: 37- 75.
[43] 李牧. 東北三大硬闊年輪與氣候關系[D]. 哈爾濱: 東北林業大學, 2013.
[44] Mikola P. Temperature and tree growth near the northern timberline // Kozlowski T T, ed. Tree Growth. New York: Ronald Press, 1962: 265- 274.
[45] Sun S C, Jin D M, Shi P L. The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient: an invariant allometric scaling relationship. Annals of Botany, 2006, 97(1): 97- 107.
[46] Westoby M, Wright I J. The leaf size-twig size spectrum and its relationship to other important spectra of variation among species. Oecologia, 2003, 135(4): 621- 628.
[47] 楊冬梅. 貢嘎山植物小枝功能性狀在不同生活型和不同海拔間的比較研究[D]. 成都: 中國科學院成都生物研究所, 2009.
[48] 郭慶學, 柴捷, 錢鳳, 張騰達, 劉文丹, 陶建平. 不同木本植物功能型當年生小枝功能性狀差異. 生態學雜志, 2013, 32(6): 1465- 1470.
Impact of simulated climate warming on the radial growth ofLarixgmeliniiin northeast China
LIU Yujia,GU Zhuoxin,WANG Xiaochun*
CenterforEcologicalResearch,NortheastForestryUniversity,Harbin150040,China
Climate warming has been widely reported across the globe, especially in high-latitude regions. There is an increasing body of evidence that a temperature increase would greatly affect tree growth and forest ecosystem. To simulate the effects of climate warming on the radial growth ofLarixgmelinii, eight-year-old young trees in Dailing, Sunwu, Songling, and Tahe (from low to high latitude) were transplanted to Maoershan (the lowest latitude) in 2004. We collected the micro-growing cores from the main trunks and segments of the branchlets from trees growing at the four transplanting origins and the one transplanting destination in the growing season of 2013. The enlarged and mature cells in main trunk and branchlet in each year were identified, and their sizes and quantities were measured by paraffin section methods. The results showed that the size and quantity of enlarged and mature cells in theLarixgmeliniitrunk increased with the simulated temperature increase, but the radial growth increment of the main trunk did not respond to the temperature increase. In the early part of the growing season (June), the quantity of mature cells in Dailing was significantly higher than that of the other two groups, whereas the cell number in August was relatively lower, indicating a pattern of continuous growth. In the late part of the growing season (August), the size and differentiation number of enlarged cells in the transplanting origin of Songling was 5 μm and 4, respectively, whereas these values were 9 μm and 7 in the transplanting destination (Maoershan), representing a nearly 2-fold increase. The size of mature cells in the transplanting origin of Songling was larger than that in the other two groups, which may be related to the change in water status. Temperature increase didn′t directly change the physiological characteristics, such as plant growth period, which might be controlled by genetic factors. We found that the branchlets were more affected by the temperature increase than the main trunk. The radial growth ofLarixgmelinii, including the diameter, number, and growth period of the tracheids, is expected to decline when global temperature increases, especially during epochs with the contemporary precipitation decrease. Our provenance experiments provide important original data to benchmark the predictions on the climate warming effects onLarixgmeliniigrowth and forest ecosystem changes in northeast China.
Larixgmelinii; tree rings; global warming; cambium; slicing technology
國家“十二五”科技支撐項目(2011BAD37B01);教育部新世紀優秀人才支持計劃(NCET- 12-0810);長江學者和創新團隊發展計劃(IRT- 15R09)共同資助
2015- 12- 17; 網絡出版日期:2016- 10- 29
10.5846/stxb201512172515
*通訊作者Corresponding author.E-mail: wangxc-cf@nefu.edu.cn
劉玉佳,顧卓欣,王曉春.模擬氣候變暖對東北興安落葉松徑向生長的影響.生態學報,2017,37(8):2684- 2693.
Liu Y J,Gu Z X,Wang X C.Impact of simulated climate warming on the radial growth ofLarixgmeliniiin northeast China.Acta Ecologica Sinica,2017,37(8):2684- 2693.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
