腺苷酸核糖基化因子1對奶牛乳腺上皮細胞增殖與泌乳的影響
2017-08-22 03:04:08穆瑩鄭冰張宏偉徐漸
中國乳品工業 2017年7期
穆瑩,鄭冰,張宏偉,徐漸
(1.東北農業大學食品學院,哈爾濱150030;2.順德出入境檢驗檢疫局,廣東佛山528000)
腺苷酸核糖基化因子1對奶牛乳腺上皮細胞增殖與泌乳的影響
穆瑩1,鄭冰2,張宏偉1,徐漸1
(1.東北農業大學食品學院,哈爾濱150030;2.順德出入境檢驗檢疫局,廣東佛山528000)
以奶牛乳腺上皮細胞為模型,研究了腺苷酸核糖基化因子1(ARF1)對細胞泌乳及增殖的影響。對細胞分別進行ARF1過表達與抑制、mTOR抑制及mTOR抑制后ARF1過表達處理,用MTT法檢測細胞增殖,試劑盒檢測乳糖和乳脂分泌,蛋白質免疫印跡檢測β-酪蛋白(CSN2)及ARF1,mTOR,p-mTOR,Cyclin D1,SREBP1的表達。結果表明:ARF1過表達或抑制后,細胞增殖與泌乳及p-mTOR,Cyclin D1,SREBP1的表達均顯著增加或降低;mTOR表達變化不顯著。mTOR抑制后,細胞增殖與泌乳及p-mTOR、Cyclin D1、SREBP1的表達均顯著降低,ARF1表達變化不顯著;并且,在mTOR被抑制的細胞中,過表達ARF1,不能使細胞的細胞增殖與泌乳及p-mTOR,Cyclin D1,SREBP1的表達恢復。實驗說明,ARF1是奶牛乳腺上皮細胞中細胞增殖和泌乳的正向調控因子,它通過mTORC1通路調控細胞增殖和泌乳。
腺苷酸核糖基化因子1;奶牛乳腺上皮細胞;細胞增殖;泌乳;mTORC1通路
0 引言
奶牛乳腺細胞的增殖及泌乳受激素、營養素和能量等多種因素的調控[1-3]。在奶牛乳腺上皮細胞中,哺乳動物雷帕霉素靶蛋白復合體1(mTORC1)信號通路對細胞增殖及泌乳有重要調控作用,mTORC1能夠廣泛接收各種信號,調控細胞的增殖及泌乳[4-6]。研究表明,細胞中mTORC1通路的激活、細胞增殖及泌乳的調控是一個復雜的網絡調控過程[7-10]。并且,就目前而言,細胞中有哪些因子參與這一調控過程?它們的調控通路及機制是什么?這些都還知之甚少。本研究以泌乳奶牛乳腺上皮細胞為材料,研究腺苷酸核糖基化因子1(ARF1)對細胞中mTORC1通路的激活及細胞增殖與泌乳的調控作用。本研究完善了mTORC1調控乳腺細胞增殖及泌乳的調控通路,加深了人們對乳腺細胞增殖及泌乳的調控機制的認識。
1 實驗
1.1 材料
奶牛乳腺上皮細胞由本實驗室純化保存。DMEM/F12培養基,胎牛血清(FBS),Trizol試劑,脂質體2000,DMSO,雙抗,Flag標簽表達載體,MTT,反轉錄試劑,PCR試劑,限制性內切酶,T4 DNA連接酶,si-RNA干擾片段,乳糖檢測試劑盒,甘油三酯檢測試劑盒,SREBP1抗體,CyclinD1抗體,ARF1抗體,mTOR抗體,p-mTOR抗體,β-actin抗體,β-casein抗體,HRP標記的二抗。
1.2 奶牛乳腺上皮細胞的培養
奶牛乳腺上皮細胞用DMEM/F12培養基(添加10%FBS和100 U/mL的雙抗),37℃,CO2培養箱中培養[10]。進行實驗的細胞以5.0×104個/mL培養液的密度接種于6孔板,待細胞長至約80%匯合度時按實驗需求進行處理,收樣備用。
1.3 表達載體構建與si-RNA片段的合成
Trizol法提取奶牛乳腺上皮細胞總RNA,反轉錄成cDNA作為模板,根據NCBI基因庫中奶牛ARF1的基因序列(NM_176653.4)設計特異性引物,進行PCR擴增ARF1基因,擴增的目的條帶回收、酶切、克隆進真核表達載體pCMV-C-Flag、測序。測序正確的陽性克隆擴大培養、提取質粒備用(ARF1過表達載體標記為AO)。PCR擴增引物及酶切位點如下:
上游引物:5’-CCGGAATTCATGGGGAATATC TTTGCA-3’下劃線為EcoR I酶切位點;
下游引物:5’-GCGTCGACTTTCTGGTTCCG?GAGCTG-3’下劃線為Sal I酶切位點。
ARF1及mTOR的si-RNA特異性干擾片段、陰性對照片段均由蘇州吉瑪基因股份有限公司合成。
1.4 細胞轉染
細胞在6孔板中培養至80%匯合度,換成無血清無雙抗的培養基培養,空載體(EV)、ARF1過表達載體(AO)、陰性對照片段(NC)、ARF1干擾片段(si-A)及mTOR干擾片段(si-M)均按實驗需求用脂質體2000進行轉染,實驗操作參照說明書。轉染培養6 h后,更換能有血清有雙抗的正常培養基常規培養24 h,收樣待檢。
ARF1過表達實驗分組為:空白對照組(B)、轉空質粒組(EV)和ARF1過表達組(AO);ARF1抑制實驗分組為:空白對照組(B)、轉陰性片段組(NC)和ARF1抑制組(si-A);ARF1和mTOR雙處理實驗分組為:空白對照組(B)、轉陰性片段組(NC)、mTOR抑制組(si-M)和mTOR抑制/ARF1過表達組(si-M/ AO)組。
1.5 細胞增殖檢測(MTT法)
細胞以5.0×103個/孔的密度接種于96孔板中,正常培養12 h,根據實驗需要進行轉染處理20 h,然后在培養液中添加MTT至終質量濃度為0.5 g/L(MTT母液用PBS稀釋),繼續培養4 h。棄培養液,加150 μL/孔的DMSO,室溫震蕩孵育10 min,用酶標儀檢測樣品在450 nm的吸光度值[11]。
1.6 細胞活力分析
細胞處理24 h后,細胞樣品用CASY-ton緩沖液1∶100稀釋,然后用CASY細胞分析儀檢測細胞活力。
1.7 乳糖與乳脂檢測
細胞處理24 h后收集細胞培養液,用乳糖和甘油三酯檢測試劑盒檢測培養液中的乳糖和甘油三酯。
1.8 蛋白質免疫印跡(WB)分析
細胞用6孔板正常培養,至所需匯合度(70%~80%),然后按實驗需要進行添加氨基酸或基因過表達與抑制處理。各組細胞處理24 h后,常規方法進行蛋白質免疫印跡(WB)分析。WB實驗中,兔多克隆β-casein一抗使用濃度為1∶1000;兔多克隆mTOR一抗、p-mTOR一抗、GLUT1一抗、SREBP-1c一抗、p-SREBP-1c一抗、GPR87一抗、鼠多克隆β-actin使用濃度均為1∶500;所有HRP標記的二抗使用濃度為1∶2000。
1.9 數據統計與分析
實驗數據用Microsoft Excel進行統計分析,用單因素方差分析分析組間差異,結果用“平均值±標準差”表示。WB灰度值用ImageJ2x軟件分析。
2 結果與分析
2.1 ARF1對細胞增殖的影響
對培養的奶牛乳腺上皮細胞進行ARF1的過表達和抑制,用MTT法檢測細胞增殖,CASY細胞活力分析儀檢測細胞活力。ARF1過表達實驗分為空白對照組(B)、轉空質粒組(EV)和ARF1過表達組(AO);ARF1抑制實驗分為空白對照組(B)、轉陰性片段組(NC)和ARF1抑制組(si-A)。結果如圖1所示,在ARF1過表達實驗中,與B組和EV組比較,AO組細胞增殖與細胞活力顯著增加(p<0.05)(圖1(a)(c));在ARF1抑制實驗中,與B組和NC組比較,si-A組細胞增殖與細胞活力顯著降低(p<0.05)(圖1(b)(d))。結果說明,ARF1能正向調控奶牛乳腺上皮細胞的細胞增殖和細胞活力。

圖1 ARF1過表達與抑制對細胞增殖及細胞活力的影響
圖1中,(a)和(b)為MTT法檢測ARF1過表達(a)和抑制(b)對細胞增殖的影響。空白組(B組)細胞增殖率定義為“1”;(c)和(d)為CASY細胞活力分析儀檢測ARF1過表達(c)和抑制(d)對細胞活力的影響。B表示空白對照組,EV表示轉空質粒組,AO表示ARF1過表達組,NC表示陰性對照組,si-A表示ARF1抑制組。“??”表示與空白組比較差異顯著(p<0.05)。
2.2 ARF1對細胞泌乳的影響
對培養的奶牛乳腺上皮細胞進行ARF1的過表達和抑制,收樣。用試劑盒檢測細胞培養液中乳糖和乳脂的濃度,蛋白質免疫印跡檢測細胞中乳蛋白的合成。實驗分組同細胞增殖與活力檢測實驗,結果如圖2所示。由圖2可以看出,在ARF1過表達實驗中,與B組和EV組比較,AO組細胞分泌乳糖、乳脂及合成CSN2的能力顯著增加(p<0.05)(圖2(a)(b)(c)(d));在ARF1抑制實驗中,與B組和NC組比較,si-A組細胞分泌乳糖、乳脂及合成CSN2的能力顯著降低(p<0.05)(圖2(e)(f)(g)(h))。結果說明,ARF1能正向調控奶牛乳腺上皮細胞的乳糖、乳脂分泌及乳蛋白合成。
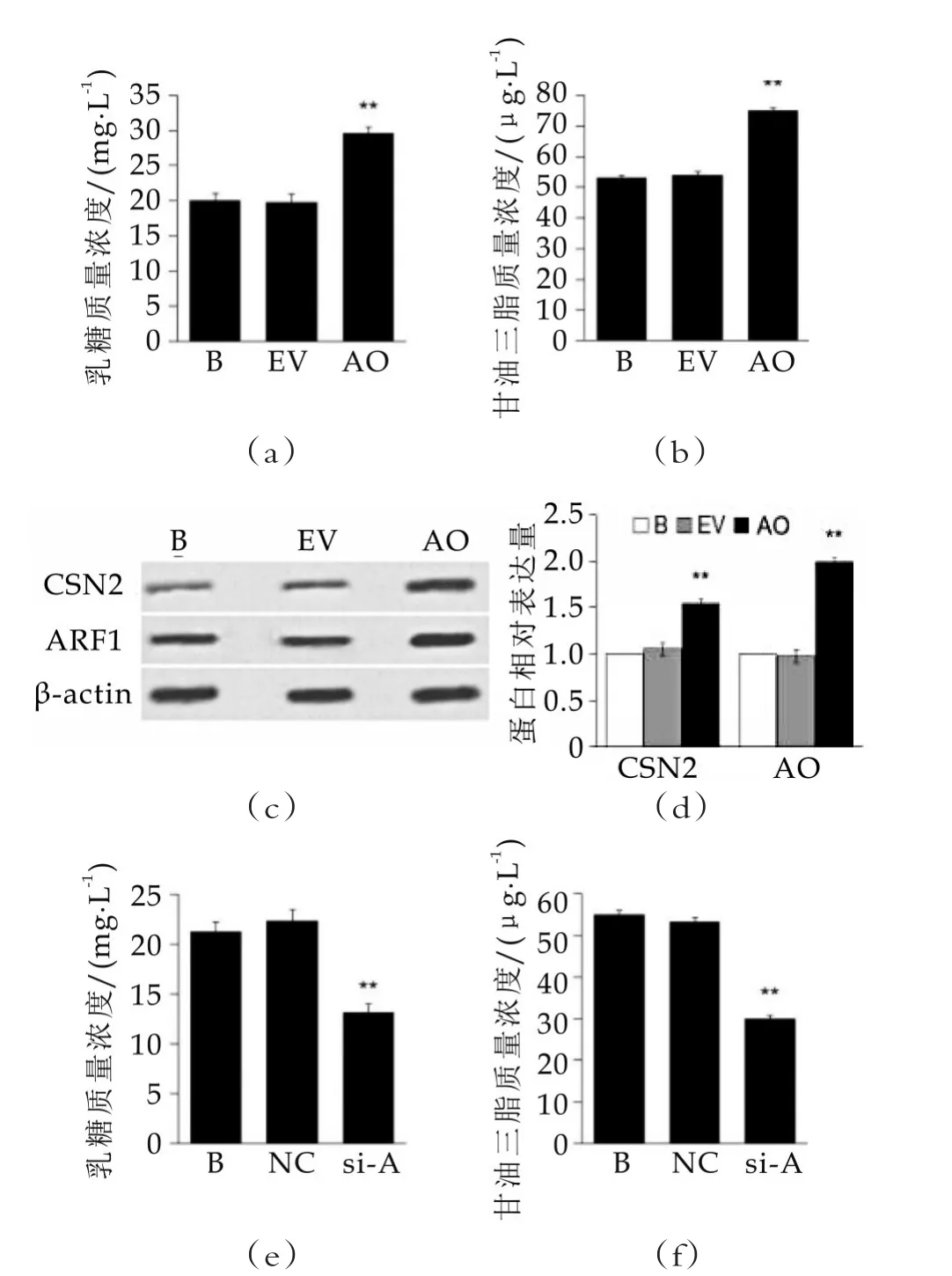
圖2 ARF1過表達與抑制對細胞泌乳能力的影響
圖2中,(a)和(b)為試劑盒檢測ARF1過表達對細胞分泌乳糖(a)和乳脂(b)的影響;(c)和(d)為蛋白質免疫印跡檢測ARF1過表達對細胞中CSN2表達的影響;(e)和(f)為試劑盒檢測ARF1抑制對細胞分泌乳糖(e)和乳脂(f)的影響;(g)和(h)為蛋白質免疫印跡檢測ARF1抑制對細胞中CSN2表達的影響,空白組(B組)蛋白灰度值定義為“1”。B表示空白對照組,EV表示轉空質粒組,AO表示ARF1過表達組,NC表示陰性對照組,si-A表示ARF1抑制組。“??”表示與空白組比較差異顯著(p<0.05)。
2.3 ARF1對mTOR、p-mTOR、CyclinD1和SREBP1表達的影響
對培養的奶牛乳腺上皮細胞進行ARF1的過表達和抑制,收樣。用蛋白質免疫印跡檢測細胞中mTOR、p-mTOR、Cyclin D1和SREBP1的表達。實驗分組同細胞增殖與活力檢測實驗。結果如圖3所示,在ARF1過表達實驗中,與B組和EV組比較,AO組細胞中p-mTOR、Cyclin D1和SREBP1的表達顯著增加(p<0.05),但mTOR的表達無顯著變化(p>0.05)(圖3(a)(b));在ARF1抑制實驗中,與B組和NC組比較,si-A組細胞中p-mTOR、Cyclin D1和SREBP1的表達顯著降低(p<0.05),但mTOR的表達無顯著變化(p>0.05)(圖3(c)(d))。結果說明,ARF1能促進奶牛乳腺上皮細胞中的mTOR磷酸化,激活mTOR通路,上調Cyclin D1和SREBP1的表達。
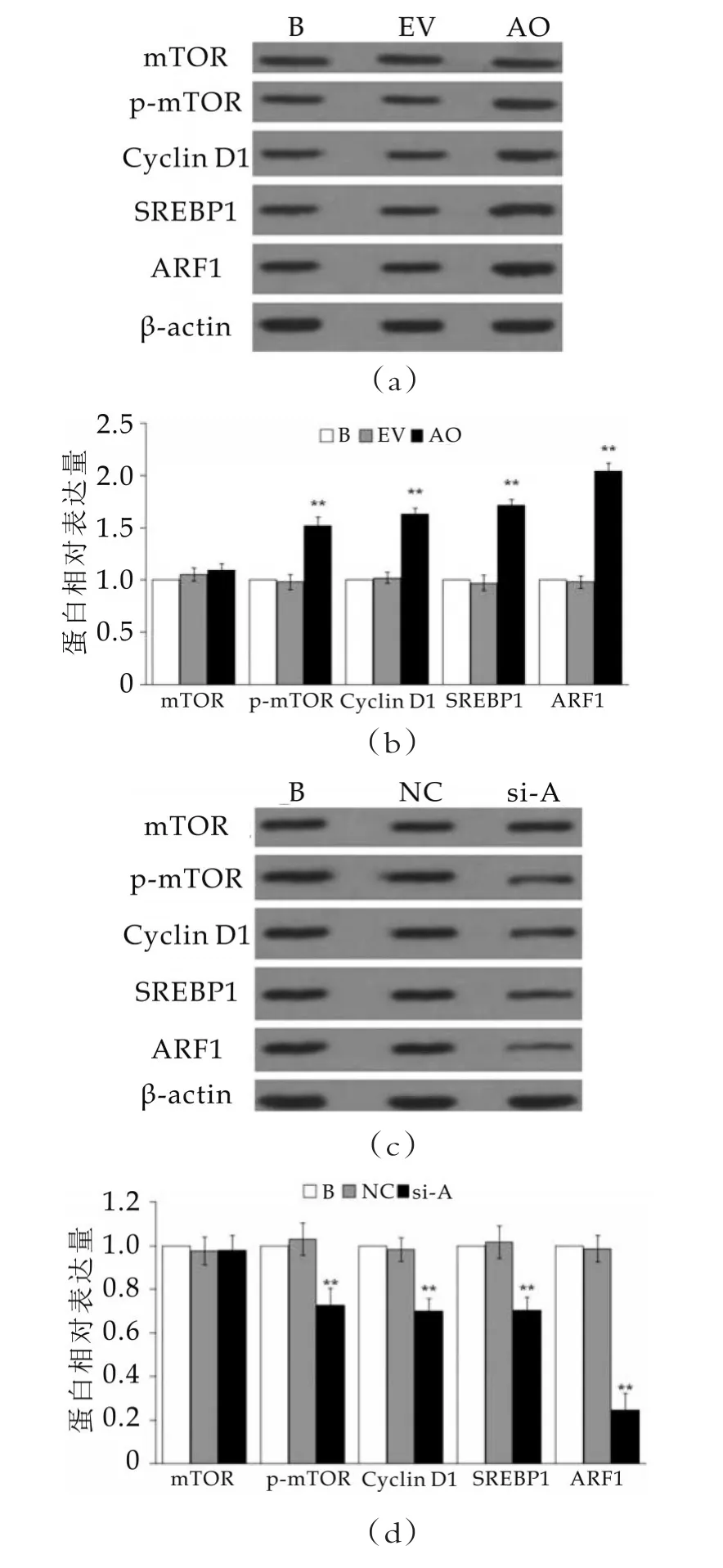
圖3 ARF1過表達與抑制對mTOR,p-mTOR,Cyclin D1和SREBP1表達的影響
圖3中,(a)和(b)為蛋白質免疫印跡檢測ARF1過表達對細胞中mTOR,p-mTOR,Cyclin D1和SREBP1表達的影響;(c)和(d)為蛋白質免疫印跡檢測ARF1抑制對細胞中mTOR,p-mTOR,Cyclin D1和SREBP1表達的影響,空白組(B組)蛋白灰度值定義為“1”。B表示空白對照組,EV表示轉空質粒組,AO表示ARF1過表達組,NC表示陰性對照組,si-A表示ARF1抑制組。“??”表示與空白組比較差異顯著(p<0.05)。
2.4 ARF1通過mTORC1通路調控細胞增殖與泌乳
對培養的奶牛乳腺上皮細胞進行mTOR抑制、mTOR抑制后過表達ARF1處理,收樣。用MTT法檢測細胞增殖,CASY細胞活力分析儀檢測細胞活力,試劑盒檢測細胞培養液中乳糖和乳脂的濃度,蛋白質免疫印跡檢測細胞中ARF1、CSN2、Cyclin D1和SREBP1的表達。實驗分為空白對照組(B)、陰性對照組(NC)、mTOR抑制組(si-M)和mTOR抑制/ ARF1過表達組(si-M/AO)組。結果如圖4所示,在細胞增殖方面,與B組和NC組比較,si-M組細胞增殖與細胞活力顯著下降(p<0.05),并且mTOR抑制后,過表達ARF1并不能使細胞增殖與細胞活力恢復(圖4(a)(b));在泌乳能力方面,與B組和NC組比較,si-M組細胞分泌乳糖、乳脂及合成CSN2的能力顯著下降(p<0.05),并且mTOR抑制后,過表達ARF1并不能使細胞分泌乳糖、乳脂及合成CSN2的能力恢復(圖4(c)(d)(e)(f))。此外,與B組和NC組比較,si-M組細胞中Cyclin D1和SREBP1的表達顯著下降(p<0.05),并且mTOR抑制后,過表達ARF1并不能使細胞中Cyclin D1和SREBP1的表達恢復(圖4(e)(f));但是,與B組和NC組比較,si-M組細胞中ARF1的表達無顯著變化(p>0.05)。結果說明,ARF1對奶牛乳腺上皮細胞增殖與泌乳及Cy?clin D1、SREBP1表達的調控是通過mTORC1通路實現的。
圖4中,(a)為MTT法檢測mTOR抑制及mTOR抑制/ARF1過表達對細胞增殖的影響;(b)為CASY細胞活力分析儀檢測mTOR抑制及mTOR抑制/ARF1過表達對細胞活力的影響;(c)和(d)為試劑盒檢測mTOR抑制及mTOR抑制/ARF1過表達對細胞分泌乳糖(c)和乳脂(d)的影響;(e)和(f)為蛋白質免疫印跡檢測mTOR抑制及mTOR抑制/ARF1過表達對細胞中CSN2、Cyclin D1及SREBP1表達的影響,空白組(B組)蛋白灰度值定義為“1”。B表示空白對照組,NC表示陰性對照組,si-M表示mTOR抑制組,si-M/AO表示mTOR抑制/ARF1過表達組。“??”表示與空白組比較差異顯著(p<0.05)。
3 討論
3.1 ARF1對奶牛乳腺上皮細胞增殖的調控

圖4 通過mTOR通路調控細胞增殖與泌乳
腺苷酸核糖基化因子(ARF)廣泛存在于真核生物中,是在動植物演化發展中高度保守的GTP激酶,屬于小G蛋白Ras家族[11-13]。ARF1是ARF家族中重要的一員,它廣泛定位于高爾基體上,與高爾基復合體相關[14-15]。目前研究表明,ARF1在囊泡運輸、磷脂代謝、細胞內物質運輸與信號轉導過程中具有重要生物學功能[16-18]。此外,近年來研究表明,ARF1還與多種腫瘤的發生密切相關,它對癌細胞的細胞增殖、PI3K信號通路及對MAPK信號通路均有調控作用[19-20]。在本實驗中,對奶牛乳腺細胞中進行了ARF1的過表達和抑制,結果顯示,ARF1過表達后,細胞生長率和細胞活力均顯著提高;ARF1抑制后,細胞生長率和細胞活力均顯著降低,這說明ARF1能夠促進奶牛乳腺上皮細胞的增殖。
本實驗還對ARF1促進乳腺上皮細胞增殖的分子機制做了初步的探究。實驗對ARF1過表達或抑制后的細胞中的mTOR、p-mTOR和Cyclin D1等與細胞增殖有關的信號分子進行了檢測。結果表明,ARF1的過表達和抑制能使細胞中的p-mTOR與Cyclin D1的表達均上升,但mTOR的表達沒變化,這說明ARF1可能是通過促使細胞中mTOR的磷酸化,激活mTORC1信號通路及促進Cyclin D1的表達來正向調控細胞增殖的。此外,實驗還進一步對細胞進行了抑制mTOR,然后過表達ARF1的雙重處理,結果顯示,mTOR抑制后,細胞的生長及活力顯著降低,細胞中p-mTOR及Cyclin D1的表達也顯著下降,但ARF1的表達無明顯變化。并且,在抑制了mTOR的細胞中過表達ARF1并不能使細胞生長及活力得到恢復,也不能使細胞中p-mTOR及Cyclin D1的表達回升。這個結果進一步說明了在奶牛乳腺上皮細胞中,ARF1是通過激活細胞中的mTORC1通路來調控細胞增殖的。
3.2 ARF1調控奶牛乳腺上皮細胞泌乳的調控
奶牛乳腺細胞泌乳的調控是一個復雜的網絡調控過程,它涉及到多個信號分子及信號通路。有研究表明,在奶牛乳腺上皮細胞中,SREBP1是細胞合成與分泌乳脂的關鍵調控因子[21];還研究表明,細胞中的mTOR及其參與的mTORC1通路對乳腺細胞的泌乳有重要調控作用,并且細胞中很多蛋白酶及具有GTP酶活性的小G蛋白都能激活細胞中的mTORC1通路[22]。有研究表明,ARF1對細胞中mTORC1信號通路及SREBP1有調控作用[23-24]。在本實驗中,對奶牛乳腺細胞中進行了ARF1的過表達和抑制,結果顯示,ARF1過表達和抑制后,細胞分泌乳糖和乳脂及合成酪蛋白的能力顯著提高和降低,這說明ARF1能夠促進奶牛乳腺上皮細胞中乳糖、乳脂的分泌和乳蛋白的合成,上調細胞的泌乳能力。
此外,本研究還進一步檢測了ARF1對細胞中泌乳相關的信號因子的表達的影響。結果顯示,ARF1過表達或抑制后,細胞中的p-mTOR與SREBP1的表達均上升,但mTOR的表達沒變化,這說明ARF1可能是通過促使細胞中mTOR的磷酸化,激活mTORC1信號通路及促進SREBP1的表達來正向調控細胞泌乳的。實驗還進一步對細胞進行了抑制mTOR,然后過表達ARF1的雙重處理,結果顯示,mTOR抑制后,細胞的泌乳能力顯著降低,細胞中p-mTOR及SREBP1的表達也顯著下降,但ARF1的表達無明顯變化。并且,在抑制了mTOR的細胞中過表達ARF1并不能使細胞的泌乳能力得到恢復,也不能使p-mTOR及SREBP1的表達回升。這個結果進一步說明了在奶牛乳腺上皮細胞中,ARF1是通過激活細胞中的mTORC1通路來調控細胞泌乳的。
4 結論
本研究表明,ARF1是奶牛乳腺上皮細胞中細胞增殖和泌乳的正向調控因子,它通過調節mTORC1通路,促進mTOR的磷酸化、Cyclin D1和SREBP1的表達調控細胞增殖和泌乳。
[1]WANG Z R,HOU X M,QU B,et al.Pten regulates development and lactation in the mammary glands of dairy cows[J].PLoS One,2014,9(7):e102118.
[2]HUANG Y L,ZHAO F,LUO C C,et al.SOCS3-Mediated block?ade reveals major contribution of JAK2/STAT5 signaling pathway to lactation and proliferation of dairy cow mammary epithelial cells in vi?tro[J].Molecules,2013,18(10):12987-13002.
[3]穆瑩,穆辛,鄭冰,等.G蛋白偶聯受體87對奶牛乳腺上皮細胞泌乳的影響[J].中國乳品工業,2016,44:20-23,31.
[4]NAN X M,BU D P,LI X Y,et al.Ratio of lysine to methionine al?ters expression of genes involved in milk protein transcription and translation and mTOR phosphorylation in bovine mammary cells[J]. Physiol Genomics,2014,46(7):268-275.
[5]穆瑩,徐漸,許巖.小G蛋白RagD對奶牛乳腺上皮細胞乳蛋白合成的影響[J].中國乳品工業,2014,42:13-16.
[6]WANG J H,DIAO Q Y,TU Y,et al.The limiting sequence and proper ratio of lysine,methionine and threonine for calves fed milk re?placers containing soy protein[J].Asian-Aust.J.Anim.Sci.,2012,25(2):224-233.
[7]穆瑩,王淼,馮一兵,等.MAP3K9對奶牛乳腺上皮細胞增殖及乳蛋白合成的影響[J].中國乳品工業,2015,43:16-20,61.
[8]PAZ H A,KONONOFF P J.Lactation responses and amino acid utili?zation of dairy cows fed low-fat distillers dried grains with solubles with or without rumen-protected lysine supplementation[J].J Dairy Sci.,2014,97(10):6519-6530.
[9]LU L M,GAO X J,LI Q Z,et al.Comparative Phosphoproteomics Analysis of the Effects of L-methionine on Dairy Cow Mammary Epi?thelial Cells[J].Canadian Journal of Animal Science,2012,92(4):433-442.
[10]LU L M,LI Q Z,HUANG J G,et al.Proteomic and functional anal?yses reveal MAPK1 regulates milk protein synthesis[J].Molecules,2012,18(1):263-275.
[11]BOTTANELLI F,KILIAN N,ERNST AM,et al.A novel physiologi?cal role for ARF1?in the formation of bi-directional tubules from the Golgi[J].Mol Biol Cell,2017,Apr 20.pii:mbc.E16-12-0863.doi:10.1091/mbc.E16-12-0863.[Epub ahead of print].
[12]KOUBEK EJ,SANTY LC.ARF1 and ARF6 regulate recycling of GRASP/Tamalin and the Rac1-GEF Dock180 during HGF-in?duced Rac1 activation[J].Small GTPases,2016,12:1-18.
[13]PACZKOWSKI J E,FROMME J C.Analysis of Arf1 GTPase-De?pendent Membrane Binding and Remodeling Using the Exomer Se?cretory Vesicle Cargo Adaptor[J].Methods Mol Biol,2016,1496:41-53.
[14]RODRIGUES F F,HARRIS T J C.Key roles of Arf small G pro?teins and biosynthetic trafficking for animal development[J].Small GTPases,2017,14:1-8.
[15]RODRIGUES FF,SHAO W,HARRIS TJ.The Arf GAP Asap pro?motes Arf1 function at the Golgi for cleavage furrow biosynthesis in Drosophila[J].Mol Biol Cell,2016,27:3143-3155.
[16]WANG Y,ZHANG H,SHI M,et al.Sec71 functions as a GEF for the small GTPase Arf1 to govern dendrite pruning of Drosophila sen?sory neurons[J].Development,2017 Apr 18.pii:dev.146175.doi:10.1242/dev.146175.
[17]XU X,WANG Q,HE Y,et al.ADP-ribosylation factor 1(ARF1)takes part in cell proliferation and cell adhesion-mediated drug resis?tance(CAM-DR)[J].Ann Hematol,2017,96:847-858.
[18]GENOT E.ARF1 at the crossroads of podosome construction and function[J].J Cell Biol,2017,216:13-15.
[19]GU G,CHEN Y,DUAN C,et al.Overexpression of ARF1 is associ?ated with cell proliferation and migration through PI3K signal path?way in ovarian cancer[J].Oncol Rep,2017,37:1511-1520.
[20]DAVIS J E,XIE X,GUO J,et al.ARF1 promotes prostate tumori?genesis via targeting oncogenic MAPK signaling[J].Oncotarget,2016,7:39834-39845.
[21]LI N,ZHAO F,WEI C,et al.Function of SREBP1 in the milk fat synthesis of dairy cow mammary epithelial cells[J].Int J Mol Sci,2014,15:16998-17013.
[22]WANG L N,LIN Y,BIAN Y J,et al.Leucyl-tRNA synthetase reg?ulates lactation and cell proliferation via mTOR signaling in dairy cow mammary epithelial cells[J].Int.J.Mol.Sci,2014,15(4):5952-5969.
[23]崔依同,王沖,孫大曉,等.ASAP1和ARF1通過調節mTOR重激活調控自噬性溶酶體再生[J].中國科學:生命科學,2014,44:1082-1090.
[24]SMULAN L J,DING W,FREINKMAN E,et al.Cholesterol-Inde?pendent SREBP-1 Maturation Is Linked to ARF1 Inactivation[J]. Cell Rep,2016,16:9-18.
Effects of ADP ribosylation factor 1 on proliferation and lactation in dairy cow mammary epithelial cells
MU Ying1,ZHENG Bing2,ZHANG Hongwei1,XU Jian1
(1.Food Science College of northeast agricultural university,Harbin 150030,China;2.Entry-exit Inspection and Quar?antine Bureau of Shunde,Foshan 528000,China)
The role of ADP ribosylation factor 1(ARF1)in cell proliferation and lactation of bovine mammary epithelial cells(BMECs)was investigated.The BMECs were treated with ARF1 overexpression,ARF1 silencing,mTOR silencing,and mTOR silencing/ARF1 overex?pression,respectively.The cell proliferation was detected by MTT;the secretion of lactose and milk fat was detected by lactose assay kit and triglyceride detection kit,respectively;the expression ofβ-casein(CSN2),ARF1,mTOR,p-mTOR,Cyclin D1,and SREBP1 was detect?ed by western blotting.The result showed that the BMECs treated with ARF1 overexpression and silencing,the cell proliferation,lactation,and the expression of p-mTOR,Cyclin D1,and SREBP1 were significant increased and significant decreased,respectively,but the expres?sion of mTOR was no significant changed.The BMECs treated with mTOR silencing,the cell proliferation,lactation,and the expression of p-mTOR,Cyclin D1,and SREBP1 were significant decreased,but the expression of ARF1 was no significant changed.What’s more,after mTOR silencing,the cell proliferation,lactation,and the expression of p-mTOR,Cyclin D1,and SREBP1 were not restored in the BMECs treated with ARF1 overexpression.These results suggested that ARF1 can positively regulate cell proliferation and lactation via mTORC1 pathway.
ADP ribosylation factor 1;bovine mammary epithelial cells;cell proliferation;lactation;mTORC1 pathway
TS252.1
:A
:1001-2230(2017)07-0018-05
2017-01-16
穆瑩(1982-),女,碩士,研究方向為乳品加工技術。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
