
DNA識別受體在乙型肝炎發病機制中的作用
2017-09-18 01:04:27龐秀青謝冬英
臨床肝膽病雜志 2017年9期
關鍵詞:機制
黃 勁, 龐秀青, 謝冬英
(中山大學附屬第三醫院 肝病實驗室, 廣州 510630)
DNA識別受體在乙型肝炎發病機制中的作用
黃 勁, 龐秀青, 謝冬英
(中山大學附屬第三醫院 肝病實驗室, 廣州 510630)
乙型肝炎作為一種全球性疾病至今仍危害著人類健康,但是HBV導致的肝炎發病機制仍未明確。肝臟中的固有免疫系統能監測HBV感染,并且運用各種策略清除病毒,其中DNA識別受體發揮了重要的作用。DNA識別受體在胞質或者胞核中識別HBV DNA或pgRNA,通過各種信號通路激活固有免疫從而產生炎性因子及IFN,最終發揮抗病毒作用。總結了各種參與HBV致炎及HBV清除的DNA識別受體,闡述了它們的具體作用途徑,并對DNA識別受體在HBV誘導的肝臟固有免疫作用中存在的問題作出討論及展望。
肝炎, 乙型; 受體, 模式識別; 綜述
現階段全球約有20億人感染過HBV,其中約4億人是HBV慢性攜帶者,而且每年約有5千萬至1百萬人死于HBV相關終末期肝病或肝癌[1-2]。HBV感染導致的肝臟疾病早已進入全球疾病致死榜前10位。但是,HBV感染導致的肝炎發病機制至今仍未明確[3]。既往大量研究[4]表明,盡管HBV不會對肝細胞造成直接破壞,但是HBV感染機體后所觸發的免疫應答在肝炎的發生發展以及病毒的清除過程中發揮了至關重要的作用。而關鍵的問題是機體的免疫系統是如何識別HBV的。隨著對DNA識別受體作用認識的不斷更新,為這個問題的解答帶來了新的方向。本文主要總結了現階段關于DNA識別受體參與HBV導致肝炎的發病機制的研究,并對DNA識別受體存在的問題作出討論和展望。
1 DNA識別受體
既往研究[5]表明核苷酸在免疫應答中發揮著重要的作用,但是人們對核苷酸具體如何激發機體的免疫應答了解很少。繼2000年Hemmi等[6]發現胞膜DNA識別受體——Toll樣受體9后,DNA識別受體開始進入人們視線,并成為免疫學領域的一大熱點。至今為止已有10余種DNA識別受體被報道,它們在對外源或內源損傷DNA識別后通過不同的方式觸發機體免疫反應[7-8]。細胞中的DNA識別受體有很多,它們能識別各種不同來源的DNA,包括細菌、病毒和凋亡細胞。作為一種病原相關分子模式,外源DNA如HBV DNA能夠被細胞中的模式識別受體識別,激活固有免疫應答信號通路,誘導并上調IFN和促炎性細胞因子基因的轉錄和表達,從而啟動固有免疫防御機制,同時促進產生特異性免疫應答[9-10]。
DNA識別受體參與固有免疫的作用在HBV相關肝炎的發病機制中是否也同樣存在,已有許多相關的研究證實了這一猜想。作為一種雙鏈DNA(dsDNA)病毒,HBV感染機體后通過血液到達肝臟,其基因組DNA在肝臟中首先被固有免疫系統識別,隨后誘導產生各種炎性因子,并且通過固有免疫細胞如自然殺傷細胞、自然殺傷T淋巴細胞等引起特異性免疫,最終發揮抗病毒作用[11]。其中肝臟內不同的DNA識別受體參與誘發肝臟炎癥的通路可能不同,而具體的機制仍有待進一步明確。
2 參與HBV相關肝臟炎癥發生的DNA識別受體
2.1 黑素瘤缺乏因子(absent in melanoma,AIM)2與人干擾素誘導蛋白(interferon-inducible protein,IFI)16 2009年,多家研究機構幾乎同時報道發現AIM2[12-14]。AIM2屬于PYHIN蛋白家族成員,分子結構中包含了1個HIN 200結構域和1個Pyrin結構域。HIN 200結構域能夠識別dsDNA,促使AIM2寡聚化和纖維化;Pyrin結構域則介導招募凋亡相關斑點樣蛋白,進而激活半胱氨酸蛋白酶(caspase)1。活化的caspase-1對IL-1β和IL-18的成熟具有重要作用[15-16]。筆者團隊研究[17]發現,AIM2在慢性乙型肝炎患者的肝組織中高表達,并且其表達水平與肝炎的嚴重程度呈正相關。而在體外實驗中,AIM2能夠在轉染了HBV DNA質粒的HepG2細胞中誘導產生IL-18,并且隨著加入的AIM2抑制劑濃度越高,IL-18產生的量越低;而同樣加入caspase-1抑制劑量越多,IL-18表達也越低。這就表明,AIM2作為一種胞漿內的DNA識別受體,可能在肝細胞中識別了HBV DNA,激活了caspase-1通路,最終誘導產生炎性因子,導致肝炎的發生[18]。
IFI16與AIM2同屬于PYHIN蛋白家族成員,分子結構中包含2個HIN結構域( HIN-A和HIN-B)和1個Pyrin結構域。 IFI16能夠識別多種病毒的dsDNA,通過STING-TBK1-干擾素調節因子(interferon regulatory factor,IRF)3信號通路上調IFNβ的表達。IFI16高表達于慢性乙型肝炎患者肝組織的免疫細胞中,而IFI16的表達水平同樣與肝臟炎癥和纖維化程度呈正相關。但是IFI16在肝臟免疫細胞中如何發揮對HBV DNA的識別,以及如何觸發后續的固有免疫反應仍有待明確,這可能需要更深入的體外實驗,如利用免疫細胞和肝細胞共培養來進一步探索。
2.2 環鳥苷酸-腺苷酸合成酶(cyclic GMP-AMP synthetase,cGAS) cGAS于2013年被發現,同時其也被認為在介導胞質DNA識別中起關鍵作用。cGAS蛋白結構與2′-5′-寡腺苷酸合成酶(2′-5′-oligoadenylatesynthase,OAS)1高度相似,OAS1能特異性識別胞內雙鏈RNA(dsRNA),而cGAS主要識別的是細胞質中的dsDNA。未活化的cGAS以自抑制形式的二聚體存在,當外源或內源DNA進入細胞質后,每個cGAS分子都能識別并結合1個DNA分子,形成2∶2的cGAS-DNA復合物。cGAS-DNA復合物能激活環鳥苷酸-腺苷酸,并且使STING磷酸化[19-20]。活化的STING從內質網遷移至高爾基體,然后停留在位于核周的內涵體上。STING通過其C端結構域結合并磷酸化TBK1。隨后,STING-TBK1復合體進入內涵體,結合并激活IRF3和核因子-κB(NF-κβ),活化的IRF3和NF-κB進入細胞核,啟動固有免疫相關基因的轉錄[21]。2016年,日本學者Dansako等[22]運用體外實驗發現cGAS能夠識別HBV DNA,并且通過cGAS-STING-IRF3通路激活固有免疫反應,最后產生免疫血清球蛋白和IFN。實驗還得出,cGAS-STING可以通過抑制HBV的復制起到抗病毒作用。關于cGAS-STING通路參與HBV相關肝炎的發病機制仍需要進一步證實。
2.3 DNA依賴的干擾素調節因子激活物(DNA-dependent activation of interferon regulatory factor,DAI) 2007年,日本學者[23]研究發現胞質蛋白DLM-1(又名ZBP1)能夠識別胞質DNA,激活IRF3并上調Ⅰ型IFN的表達,因此將這種蛋白質命名為DAI[24]。HBV復制型質粒轉染HepG2細胞后能誘導Ⅰ型IFN應答,王秋景等[25]運用RNA干擾技術下調DAI在HepG2的表達后,HBV的復制和蛋白分泌受到了抑制,并且抑制效果有劑量依賴性。接著在DAI下調后檢測HBV質粒誘導的IFIT1和IL-6,結果顯示二者均不受影響,說明DAI分子并沒有在其中發揮作用。另一方面,當同時下調TBK1和DAI后,HBV抗原表達受到明顯抑制,表明DAI分子非但不能通過Ⅰ型IFN信號通路抑制HBV,反而能支持HBV的蛋白表達。Upton等[26]早在2012年就發現DAI既不通過IFN調節因子-Ⅰ型IFN信號通路,也非通過受體相互作用蛋白激酶1-NF-κB通路,而是通過DAI與核糖體鈍化蛋白(ribosome-inactivating protein,RIP)3形成復合物介導小鼠巨細胞病毒誘導的病毒感染細胞的程序性死亡,但是DAI是否真的通過RIP3參與調節HBV的復制仍有待進一步證實。
2.4 Ku70/80復合體 Ku70/80復合體在細胞中具有多種生物學功能,其中包括免疫細胞V(D)J重組、穩定端粒結構和受損DNA修復等[27]。2016年Li等[28]發現Ku70/80復合體能在細胞質中識別HBV DNA,并且在HBV DNA轉染模型中上調趨化因子CCL3和CCL5的表達。Ku70/80復合體被廣泛認為是損傷DNA的修復分子,它能在細胞核內監測DNA的破壞和非同源末端連接。盡管Ku蛋白早在10年前就被報道能夠轉移至細胞質中,但是它識別胞質DNA的能力是近年才被發現的。Zhang等[29]揭示Ku70能識別不同構型的DNA,并且通過IRF1/IRF7依賴通路促進Ⅲ型IFN的表達。另外,Ferguson等[27]研究發現Ku蛋白通過協同DNA-PKcs識別DNA,而后激活TBK1-IRF3信號通路。不僅如此,當Ku70/80復合體的DNA結合能力被病毒疫苗、細胞因子IL-6、CXCL10等阻斷后,下游的固有免疫信號分子會隨之下調。在轉染了HBV的小鼠中,CCL3和CCL5能促進免疫效應細胞聚集在肝臟中而誘發肝臟炎癥。其中可能的機制是Ku70/80復合體通過協同DNA-PKcs和聚腺苷二磷酸-核糖聚合酶(poly-ADP-ribose polymerase,PARP)1識別HBV DNA后激活IRF1依賴的信號通路,最后上調CCL3和CCL5的表達。而敲除了Ku70基因能顯著抑制CCL3和CCL5的分泌。由于CCR5在乙型肝炎的致病中起著關鍵作用,所以Ku70/80參與識別HBV DNA后誘導CCL3和CCL5上調的機制很可能是HBV相關肝炎的致病機制之一[30]。
2.5 維甲酸誘導基因(retinoic acid-inducible gene,RIG)1 RIG-1最早被認作是HCV RNA的識別受體,并且能激活固有免疫應答發揮抗HCV作用。RIG-1通過其poly-U/UC模體識別HCV RNA,最后誘導產生Ⅰ型IFN[31]。另一方面,更早的研究[32-33]表明與HCV感染相比,HBV感染的動物模型中固有免疫應答的激活受損,而且如 IFNα、 IFNβ的Ⅰ型IFN誘導能力很難被檢測出。然而,HBV是如何被人肝細胞識別以及Ⅲ型IFN的作用仍未被明確。研究[34]發現HBV感染主要誘導Ⅲ型IFN而非Ⅰ型IFN,并且這是通過RIG-1對HBV復制過程中的pgRNA 5′-ε端識別調節的。RIG-1在識別pgRNA后能阻礙HBV多聚酶(P蛋白)與pgRNA 5′-ε端相互作用而抑制HBV復制。在移植了人肝組織的小鼠體內,脂質體調節轉運和表達5′-ε端的RNA能抑制HBV的復制。因此,RIG-1和病毒RNA的相互作用能激活固有免疫應答,從而誘導IFN分泌抗病毒,且RIG-1起到直接抗病毒作用。
3 總結和展望
綜上所述,雖然既往的研究已經發現有DNA識別受體能參與HBV DNA或pgRNA的識別并觸發固有免疫抗病毒反應(圖1),但是它們各自的具體機制仍未完全明確,而且目前仍存在著許多問題。第一,DNA識別受體主要存在于細胞質中,它們是如何穿過HBV核衣殼識別基因組DNA的?近期有研究[35]發現IFI16亦存在于細胞核中,它們是否直接在細胞核內識別HBV DNA?第二,肝臟免疫細胞可吞噬壞死的肝細胞,而IFI16在HBV感染的肝臟免疫細胞中高表達,其是否可以在吞噬作用后識別壞死肝細胞釋放的自身DNA加強炎癥反應作用?第三,cGAS大量表達于肝衰竭患者肝細胞內,而且AIM2與IFI16的表達與肝臟炎癥呈正相關,提示固有免疫與肝衰竭密切相關,這些識別受體是否協同加強固有免疫作用從而促進肝衰竭的發生發展?此外,這些不同的DNA識別受體之間是否有相互拮抗、協同或者調控作用?上述問題均有待未來更多的研究來解答。如果DNA識別受體參與HBV誘導的肝炎機制清晰化,那么DNA識別受體很可能成為乙型肝炎治療的新靶點。
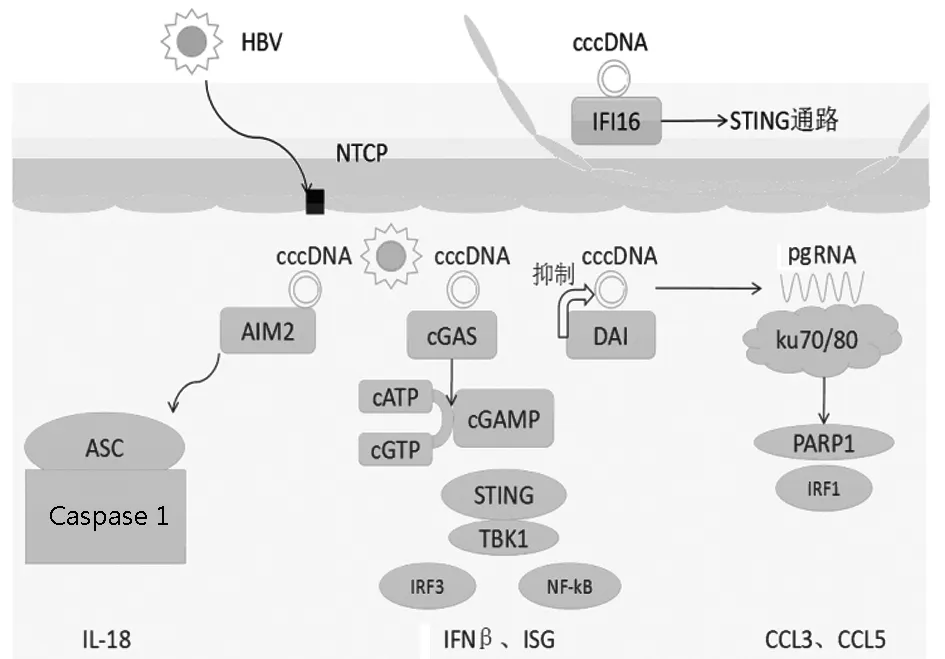
圖1 DNA識別受體在肝臟中參與HBV誘導固有免疫作用
[1] GONZALEZ SA, KEEFFE EB. Chronic viral hepatitis: epidemiology, molecular biology, and antiviral therapy[J]. Front Biosci (Landmark Ed), 2011, 16: 225-250.
[2] FARAZI PA, DEPINHO RA. Hepatocellular carcinoma pathogenesis: from genes to environment[J]. Nat Rev Cancer, 2006, 6(9): 674-687.
[3] FU WZ, YANG L, HU XQ. Relationship between HBV DNA load and HBV-M, ALT, AST content in the serum of chronic hepatitis B patients[J]. Int J Virol, 2015, 22(2): 111-114. (in Chinese) 付萬智, 楊麗, 胡曉勤. 慢性乙肝患者血清HBV DNA載量與HBV-M、ALT、AST含量的關系研究[J]. 國際病毒學雜志, 2015, 22(2): 111-114.
[4] BUSCA A, KUMAR A. Innate immune responses in hepatitis B virus (HBV) infection[J]. Virol J, 2014, 11: 22.
[5] HORNUNG V, ELLEGAST J, KIM S, et al. 5′-Triphosphate RNA is the ligand for RIG-I[J]. Science, 2006, 314(5801): 994-997.
[6] HEMMI H, TAKEUCHI O, KAWAI T, et al. A Toll-like receptor recognizes bacterial DNA[J]. Nature, 2000, 408(6813): 740-745.
[7] DEMPSEY A, BOWIE AG. Innate immune recognition of DNA: a recent history[J]. Virology, 2015, 479-480: 146-152.
[8] PALUDAN SR. Activation and regulation of DNA-driven immune responses[J]. Microbiol Mol Biol Rev, 2015, 79(2): 225-241.
[9] TAKEUCHI O, AKIRA S. Pattern recognition receptors and inflammation[J]. Cell, 2010, 140(6): 805-820.
[10] WU J, CHEN ZJ. Innate immune sensing and signaling of cytosolic nucleic acids[J]. Annu Rev Immunol, 2014, 32: 461-488.
[11] REVILL P, YUAN Z. New insights into how HBV manipulates the innate immune response to establish acute and persistent infection[J]. Antivir Ther, 2013, 18(1): 1-15.
[12] BüRCKSTüMMER T, BAUMANN C, BLüML S, et al. An orthogonal proteomic-genomic screen identifies AIM2 as a cytoplasmic DNA sensor for the inflammasome[J]. Nat Immunol, 2009, 10(3): 266-272.
[13] FERNANDES-ALNEMRI T, YU JW, DATTA P, et al. AIM2 activates the inflammasome and cell death in response to cytoplasmic DNA[J]. Nature, 2009, 458(7237): 509-513.
[14] HORNUNG V, ABLASSER A, CHARREL-DENNIS M, et al. AIM2 recognizes cytosolic dsDNA and forms a caspase-1-activating inflammasome with ASC[J]. Nature, 2009, 458(7237): 514-518.
[15] RATHINAM VA, JIANG Z, WAGGONER SN, et al. The AIM2 inflammasome is essential for host defense against cytosolic bacteria and DNA viruses[J]. Nat Immunol, 2010, 11(5): 395-402.
[16] MORRONE SR, MATYSZEWSKI M, YU X, et al. Assembly-driven activation of the AIM2 foreign-dsDNA sensor provides a polymerization template for downstream ASC[J]. Nat Commun, 2015, 6: 7827.
[17] PANG XQ, LI XH, MO ZS, et al. Expression of absent in melanoma 2 in the liver and correlation with the degrees of liver damage in patients with chronic hepatitis B[J/CD]. Chin J Exp Clin Infect Dis: Electronic Edition, 2015, 9(3): 326-330. (in Chinese) 龐秀青, 李新華, 莫志碩, 等. 慢性乙型肝炎患者肝組織中黑色素瘤缺乏因子2表達與病變程度的相關性[J/CD]. 中華實驗和臨床感染病雜志: 電子版, 2015, 9(3): 326-330.
[18] PAN X, XU H, ZHENG C, et al. Human hepatocytes express absent in melanoma 2 and respond to hepatitis B virus with interleukin-18 expression[J]. Virus Genes, 2016, 52(4): 445-452.
[19] CIVRIL F, DEIMLING T, de OLIVEIRA MANN CC, et al. Structural mechanism of cytosolic DNA sensing by cGAS[J]. Nature, 2013, 498(7454): 332-337.
[20] ZHANG X, WU J, DU F, et al. The cytosolic DNA sensor cGAS forms an oligomeric complex with DNA and undergoes switch-like conformational changes in the activation loop[J]. Cell Rep, 2014, 6(3): 421-430.
[21] ISHIKAWA H, MA Z, BARBER GN. STING regulates intracellular DNA-mediated, type I interferon-dependent innate immunity[J]. Nature. 2009, 461(7265): 788-792.
[22] DANSAKO H, UEDA Y, OKUMURA N, et al. The cyclic GMP-AMP synthetase-STING signaling pathway is required for both the innate immune response against HBV and the suppression of HBV assembly[J]. FEBS J, 2016, 283(1): 144-156.
[23] TAKAOKA A, WANG Z, CHOI MK, et al. DAI (DLM-1/ZBP1) is a cytosolic DNA sensor and an activator of innate immune response[J]. Nature, 2007, 448(7152): 501-505.
[24] KAISER WJ, UPTON JW, MOCARSKI ES. Receptor-interacting protein homotypic interaction motif-dependent control of NF-kappa B activation via the DNA-dependent activator of IFN regulatory factors[J]. J Immunol, 2008, 181(9): 6427-6434.
[25] WANG QJ, LI SB, HUANG HP, et al. Effect of the cytoplasmic DNA sensor DAI on replication of hepatitis B virus[J]. Chin J Hepatol, 2015, 23(12): 934-938. (in Chinese) 王秋景, 李世波, 黃紅平, 等. 胞質內DNA依賴的干擾素調節因子激活物在HBV復制中的作用[J]. 中華肝臟病雜志, 2015, 23(12): 934-938.
[26] UPTON JW, KAISER WJ, MOCARSKI ES. DAI/ZBP1/DLM-1 complexes with RIP3 to mediate virus-induced programmed necrosis that is targeted by murine cytomegalovirus vIRA[J]. Cell Host Microbe, 2012, 11(3): 290-297.
[27] FERGUSON BJ, MANSUR DS, PETERS NE, et al. DNA-PK is a DNA sensor for IRF-3-dependent innate immunity[J]. Elife, 2012, 1: e47.
[28] LI Y, WU Y, ZHENG X, et al. Cytoplasm-translocated Ku70/80 complex sensing of HBV DNA induces hepatitis-associated chemokine secretion[J]. Front Immunol, 2016, 7: 569.
[29] ZHANG X, BRANN TW, ZHOU M, et al. Cutting edge: Ku70 is a novel cytosolic DNA sensor that induces type III rather than type I IFN[J]. J Immunol, 2011, 186(8): 4541-4545.
[30] SANCHOOLI J, SANADGOL N, KAZEMI ARABABADI M, et al. CCR5 plays important roles in hepatitis B infection[J]. Viral Immunol, 2014, 27(1): 2-6.
[31] SAITO T, OWEN D M, JIANG F, et al. Innate immunity induced by composition-dependent RIG-I recognition of hepatitis C virus RNA[J]. Nature, 2008, 454(7203): 523-527.
[32] WIELAND S, THIMME R, PURCELL RH, et al. Genomic analysis of the host response to hepatitis B virus infection[J]. Proc Natl Acad Sci U S A, 2004, 101(17): 6669-6674.
[33] NAKAGAWA S, HIRATA Y, KAMEYAMA T, et al. Targeted induction of interferon-λ in humanized chimeric mouse liver abrogates hepatotropic virus infection[J]. PLoS One, 2013, 8(3): e59611.
[34] SATO S, LI K, KAMEYAMA T, et al. The RNA sensor RIG-I dually functions as an innate sensor and direct antiviral factor for hepatitis B virus[J]. Immunity, 2015, 42(1): 123-132.
[35] KERUR N, VEETTIL MV, SHARMA-WALIA N, et al. IFI16 acts as a nuclear pathogen sensor to induce the inflammasome in response to Kaposi Sarcoma-associated herpesvirus infection[J]. Cell Host Microbe, 2011, 9(5): 363-375.
引證本文:HUANG J, PANG XQ, XIE DY. Role of DNA recognition receptors in pathogenic mechanism of hepatitis B[J]. J Clin Hepatol, 2017, 33(9): 1794-1797. (in Chinese) 黃勁, 龐秀青, 謝冬英. DNA識別受體在乙型肝炎發病機制中的作用[J]. 臨床肝膽病雜志, 2017, 33(9): 1794-1797.
(本文編輯:葛 俊)
RoleofDNArecognitionreceptorsinpathogenicmechanismofhepatitisB
HUANGJin,PANGXiuqing,XIEDongying.
(HepatopathyLaboratory,TheThirdAffiliatedHospitalofSunYat-SenUniversity,Guangzhou510630,China)
As a global disease, hepatitis B still threatens human health. However, the pathogenesis of hepatitis caused by HBV remains unclear. The innate immune system in the liver can detect HBV infection and use every strategy to eliminate the virus. DNA recognition receptors play an important role in this process; they recognize HBV DNA or pgRNA in cytoplasm or nucleus, activate innate immunity through various signaling pathways to produce inflammatory cytokines and interferon, and finally exert their antiviral effect. This article summarizes the DNA recognition receptors involved in inflammation induced by HBV and HBV clearance, elaborates on their detailed pathways, and discusses the issues regarding the role of DNA recognition receptors in liver innate immunity induced by HBV and related perspectives.Keywords: hepatitis B; receptors, pattern recognition; review
10.3969/j.issn.1001-5256.2017.09.037
2017-04-25;
:2017-05-12。
廣東省自然科學基金項目(2015A030313022)
黃勁(1990-),男,主要從事病毒性肝炎致病機制及防治的研究。
謝冬英,電子信箱:xdy-gz@163.com。
R512.62
:A
:1001-5256(2017)09-1794-04
猜你喜歡
四川勞動保障(2021年9期)2022-01-18 05:11:08
文苑(2018年21期)2018-11-09 01:23:06
當代陜西(2018年9期)2018-08-29 01:21:00
當代陜西(2017年12期)2018-01-19 01:42:33
暨南學報(哲學社會科學版)(2016年9期)2017-01-15 13:52:00
中國衛生(2016年9期)2016-11-12 13:28:08
中國衛生(2015年9期)2015-11-10 03:11:12
醫學研究雜志(2015年12期)2015-06-10 06:57:46
中國衛生(2014年3期)2014-11-12 13:18:12
中國火炬(2014年4期)2014-07-24 14:22:19
