福建黃兔INHBA基因5’端cDNA克隆及序列分析
2017-10-11 07:32:38桑雷孫世坤陳冬金陳巖峰謝喜平福建省農業科學院畜牧獸醫研究所350013
中國畜禽種業 2017年1期
關鍵詞:分析
桑雷 孫世坤 陳冬金 陳巖峰 謝喜平 (福建省農業科學院畜牧獸醫研究所 350013)
福建黃兔INHBA基因5’端cDNA克隆及序列分析
桑雷 孫世坤 陳冬金 陳巖峰 謝喜平 (福建省農業科學院畜牧獸醫研究所 350013)
為擴增福建黃兔INHBA基因5'端,根據GenBank上已公布的家兔序列KC831577,設計了2條巢式引物,利用5'RACE技術克隆福建黃兔INHBA基因5'端序列,獲得1個682bp片段,對該片段克隆測序,獲得包括5'UTR、第1外顯子及部分第1內含子的片段。利用在線軟件預測在-229~-180bp處存在可能的啟動子,同時還發現9個潛在轉錄因子結合位點,其中包含對轉錄有重要調控作用的TATA-box位點。
福建黃兔;片段;克隆;INHBA;5'RACE;啟動子;序列分析
抑制素屬于TGF-β (transforming growth factor-bata,轉化生長因子-β)超家族成員之一,由α、β兩個亞基組成的二聚體糖蛋白激素,此類激素通過負反饋調節FSH(follicle stimulating hormone,卵泡雌激素)合成與分泌,從而影響卵泡發育和精子發生[1,2]。抑制素β亞基有βA和βB兩種形式,其中α亞基和βA組成抑制素A,此外βA與自身通過二硫鍵連接組成活化素A(Activin A,ACTA)以及與βB亞基組成活化素B(Activin B,ACTB)[3]。在母畜中,抑制素βA是影響山羊、綿羊產羔數的重要基因[4,5]。在公畜中,抑制素βA是影響荷斯坦公牛射精量及精子密度的重要因素[6]。目前對福建黃兔INHBA基因研究甚少,尤其cDNA 5’UTR(untranslated region,非翻譯區)序列未見有報導,本研究目的是解析福建黃兔INHBA基因啟動子序列及結構特征,為進一步研究福建黃兔INHBA基因調控與繁殖性狀的關聯分析提供依據。
1 材料與方法
1.1 動物及組織
性成熟的福建黃兔母兔由福建連江玉華山生態試驗場購買。黃兔母兔屠宰后立刻取卵巢組織,放入液氮罐中,備用。
1.2 總RNA提取
使用RNAiso Plus(Takara)提取黃兔卵巢組織中的總RNA,然后用不含RNA酶的Recombiant DNase I(Taraka)去除基因組DNA,隨后溶于DEPC水,中保存于-20℃備用。
1.3 引物設計及合成
本試驗以已獲得的INHBA cDNA部分序列 (KC831577)設計引物擴增5’端序列[7],由上海生工合成,引物的部分序列見表1。
1.4 5'RACE前處理及反轉錄
使用Takara 5’Full RACE Kit with TAP對5μg的總DNA進行CIAP、TAP處理,并與5’RACE Adaptor連接后,反轉錄合成cDNA。cDNA反轉錄體系:連接好的RNA 6μl,隨機 引 物( 50μM) 0.5μl, dNTPs Mixture(各 10mM)1μl,RNase Inhibitor (40U/μl) 0.25μl, 5×M-MLV buffer 2μl, Reverse Transcriptase M-MLV (200U/μl) 0.25μl。 RT 反應條件為:30℃下10min,42℃下60min,70℃下15min,反應結束后將cDNA產物保存于-20℃備用。
1.5 PCR擴增
取上述反轉錄反應液稀釋5倍取2μl,10μM上、下游引物 (INHBA 1, Outer primer) 各 2μl, 2×GC buffer 25μl, 5U/μl LA Taq?(Takara) 0.5μl, 用水補足至 50μl體系。 反應條件為: 94℃下 3min; 94℃下 30s, 60℃下 30s, 72℃下 2min,30個循環;72℃下10min。取上述PCR產物1μl作為模板,上、 下游引物 (INHNA 2, Inner primer) 各取 2μl, dNTP(各 2.5mM) 8μl, 2×GC buffer25μl, 5U/μl LA Taq?(Takara)0.5μl,用水補足至50μl體系。反應條件為:94℃下3min;94℃下 30s, 60℃下 30s, 72℃下 2min, 30個循環; 72℃下10min。取5μl進行1%瓊脂糖凝膠電泳檢測。
1.6 克隆測序
將膠回收試劑盒切膠回收的PCR產物 (約700bp)與pMD18-T載體連接后轉染至DH5α細胞中,涂平板37℃培養12h,挑選陽性克隆用PCR法鑒定重組質粒。隨后將鑒定過質粒送至寶生物工程 (大連)有限公司進行測序。
2 結果與分析
2.1 兔INHBA基因5'UTR區擴增
經過1%瓊脂糖凝膠電泳,結果顯示,3’RACE產物為大約700bp的條帶 (圖1),與預期的產物大小一致。將擴增產物送至寶生物工程 (大連)有限公司進行測序,最后得到1個682bp片段。從獲得序列分析結果來看,所獲得序列包含1個5’UTR(280bp),第1外顯子 (288bp)以及部分第1內含子 (14bp)(見圖2)。
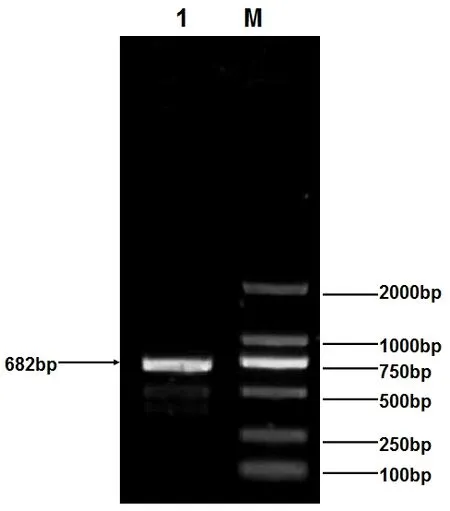
圖1 黃兔INHBA 5’端擴增產物瓊脂糖電泳

圖2 福建黃兔INHBA基因5’UTR區序列分析
2.2 兔INHBA基因的生物信息學分析
將測序得到的兔INHBA基因5’UTR區序列用BDGP∶NeuralNetwork PromoterPrediction ( http∶//www.fruitfly.org/seq_tools/promoter.html)進行啟動子位點預測,發現在-229~-180bp處存在可能的啟動子位點,得分為0.92,具體位置見圖2。
將測序得到的兔INHBA基因5’UTR序列用BIMAS網站的 Promoter Scan (https∶//www-bimas.cit.nih.gov/molbio/) 預測轉錄結合位點,發現9個潛在的轉錄因子結合位點 (表2),其中-273bp處存在 3種TFFIID結合位點,分別為TATA-box.2、 Ad2MLP US.5、 TATA-box-CS, 其中 TATA-box.2為真核生物共有的轉錄結合位點,Ad2MLP US.5、TATA-box-CS為哺乳動物特有的TFIID結合位點,-208bp、-206bp存在激活蛋白AP-2(activator protein-2)結合位點,此外還存在 glucagon-G3A (-230bp)、 UCE.2 (-227bp)、JCV_repeated_sequence(-58bp)轉錄結合位點。

表1 PCR引物擴增序列
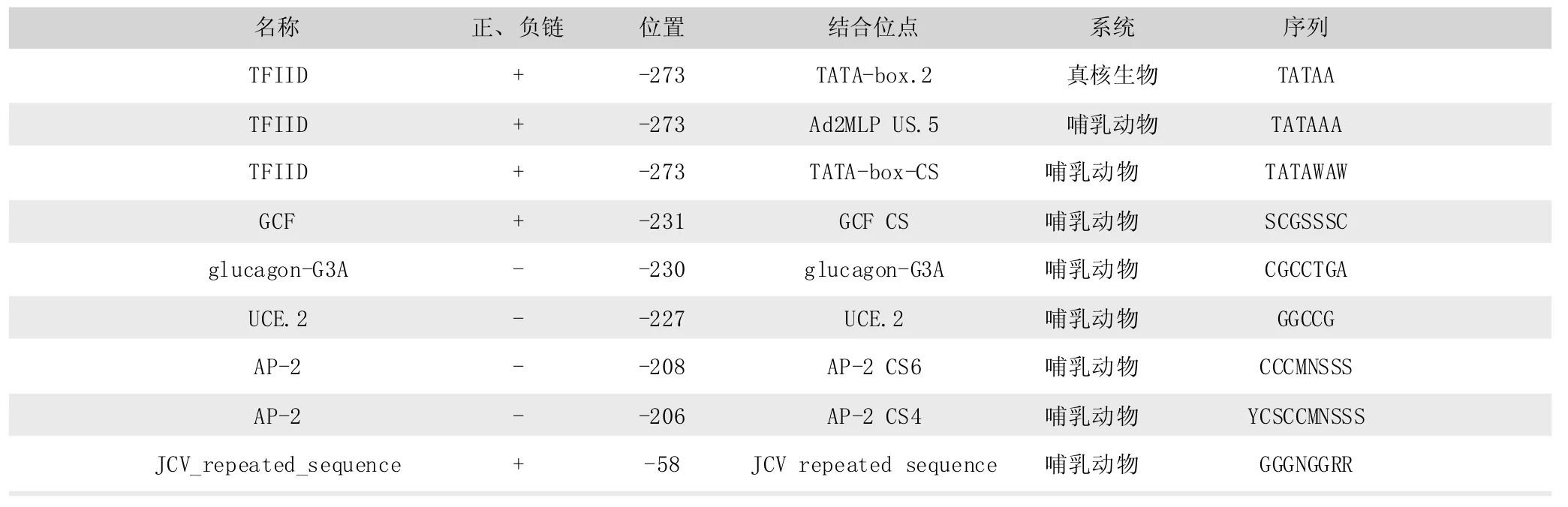
表2 轉錄結合因子
3 討論
TFIID(Transcription factor IID,轉錄因子IID)由TBP(TATA-box binding protein,TATA結合蛋白)和多種TBP協同因子 (TBP-associated factors,TAFs)組成,是真核生物中唯一具有位點特異的DNA結合能力的因子。TFIID能識別TATA位點并與之結合,形成TFIID-啟動子復合物,指導其他轉錄因子與RNA聚合酶II形成轉錄起始復合物 (transcriptional preinitiation complex),引發RNA的合成過程[8]。UCE(upstream control element,上游控制元件),與核心啟動子(core promoter) 一起被 UBF (upstream binding factor, 上游結合因子)結合,再與SL1(selectivity factor 1,選擇性因子1)一起促進轉錄[9]。GCF(GC binding factor,GC結合因子)是一種轉錄抑制因子,通過與上游啟動子中富含GC序列的區域特異結合,進而抑制基因的轉錄[10]。AP-2(activator protein-2,激活蛋白-2)是一種DNA結合蛋白,參與調控多種基因轉錄及細胞增殖、分化、凋亡[11]。Csh1、PRP-I及PAG1等基因上游有AP-2結合位點,相關研究發現Csh1、PRP-I及PAG1與胎盤子葉和胚胎發育相關,敲除小鼠胚胎細胞AP-2基因會導致合體滋養層細胞喪失增殖能力[12,13]。研究發現人滋養層細胞的hCG、hCSH1和hGRH等基因上游區域存在AP-2結合位點,這些基因的特異表達受AP-2家族的調控[14,15]。INHBA基因上GCF和AP-2位點是否與表達調控相關,有待進一步的驗證。
4 小結
本研究利用5’RACE技術獲得INHBA基因5’端1個682bp片段,通過克隆測序獲得了其5’UTR序列。通過在線分析軟件預測福建黃兔INHBA基因轉錄因子結合位置及啟動子序列,為進一步研究福建黃兔INHBA在生殖軸中的表達機制奠定了理論基礎。
[1]程萍,張昌軍.抑制素B與生殖[J].生殖醫學雜志,2007,16(3):205-208.
[2]劉賀山,袁陶燕,吳兵兵,等.抑制素基因免疫在提高動物繁殖力中的應用[J].浙江畜牧獸醫,2011(1):12-14.
[3]張成云,曾憲垠.抑制素免疫在家畜繁殖中的應用[J].畜牧獸醫雜志,2008,27(2):35-37.
[4]彭志蘭,儲明星,陳宏權,等.抑制素βA基因多態性及其與濟寧山羊高繁殖力關系[J].農業生物技術學報,2007,15(5):901-902.
[5]陳寶麒,趙宗勝,賈斌,等.新疆肉用綿羊抑制素βA基因多態性與產羔數關聯分析[J].石河子大學學報,2013,31(2):159-163.
[6]Sang L,Du Q,Yang W,et al.Polymorphisms in follicle stimulation hormone receptor,inhibin alpha,inhibin bata A,and prolactin genes,and their association with sperm quality in Chinese Holstein bulls[J].Anim Reprod Sci,2011,126(3-4):151-156.
[7]桑雷,孫世坤,陳冬金,等.福建黃兔卵泡抑制素βA亞基cDNA的克隆與序列分析[J].家畜生態學報,2013,34(9):17-20.
[8]王軍,賈弘緹.真核轉錄因子TFIID研究進展[J].生物化學與生物物理進展,1993,20(5):329-333.
[9]俞峻,王均,賈弘.TBP與TBP相關因子[J].生物化學與生物物理進展,1995,22(5):408-414.
[10]馬平,晁耐霞,潘復澤,等.GC結合因子2在肝細胞癌中的表達和意義[J].廣西醫科大學學報,2013,30(4):493-495.
[11]張翔,蘇波.轉錄因AP-2與腫瘤的研究進展[J].國際病理科學與臨床雜志,2012,32(1):40-44.
[12]Werling U,Schorle H.Transcription factor gene AP-2 gamma essential for early murine development[J].Molecular Cell Biology,2002,35:442-450.
[13]Auman H,Nottoli T,Lakiza O,et al.Transcription factor AP-2 gamma is essential in the extraembryonic lineages for early postimplantation development[J].Placenta,2007,5:16-35.
[14]Richdardson B,Cheng Y,Langland R,et al.Differential expression of AP-2 gamma and Ap-2 alpha during human trophoblast differentiation[J].Life Science,2001,69:2157-2165.
[15]Cheng Y,Aronow B,Hossain S,et al.Critical role for transcription factor AP-2 alpha in human trophoblast differentiation[J].Physiology Genomics,2004,18:99-107.
項目資金:福建省公益類研究院所專項 “閩西南黑兔FABP4基因的克隆、多態性及與生長、肉質性狀的關聯分析”(2015R1023-13);福建省農業科學院科技創新團隊PI項目 “家兔優良品種資源創新育種研究”(2016PI-11);福建農業科學院杰出青年人才基金 “閩西南黑兔FABP4基因的克隆、多態性及與生長、肉質性狀的關聯分析”(2014JQ-1)。
桑雷 (1981-),男,漢族,博士,助理研究員,從事草食動物遺傳育種與繁殖工作。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
