不同氮效率水稻根系形態和氮素吸收利用與產量的關系
2017-10-13 05:27:22李娜楊志遠代鄒孫永健徐徽何艷嚴田蓉蔣明金郭長春王春雨馬均
中國農業科學 2017年14期
李娜,楊志遠,代鄒,孫永健,徐徽,何艷,嚴田蓉,蔣明金,郭長春,王春雨,馬均
?
不同氮效率水稻根系形態和氮素吸收利用與產量的關系
李娜,楊志遠,代鄒,孫永健,徐徽,何艷,嚴田蓉,蔣明金,郭長春,王春雨,馬均
(四川農業大學水稻研究所/農業部西南作物生理生態與耕作重點實驗室,成都 611130)
【目的】探究不同氮效率水稻根系形態和氮素吸收利用與產量關系的規律,明確水稻高產根系形態特征,提出相應的水氮優化管理措施。【方法】2013年以氮高效品種川農優498和氮低效品種川優6203為試驗材料,進行不同穗肥運籌比例的裂區試驗;2014年以相同試驗材料進行穴苗數和促花肥、保花肥配比的裂裂區試驗;2015年以氮高效品種德香4103和氮低效品種宜香3724為試驗材料,進行水分管理方式和氮肥施用模式的裂裂區試驗;分別以上述3個大田試驗獲得的產量、根系形態及氮素吸收利用相關指標為樣本,通過計算方差膨脹因子診斷變量間存在的多重共線性關系。在多重共線性嚴重的情況下,運用嶺回歸分析研究根系形態、氮素吸收利用和產量三者之間的關系。基于這些關系在年度間的重演性,確定其中普遍存在的共性規律。【結果】(1)氮高效品種拔節期、抽穗期及成熟期的氮素積累量、氮素干物質生產效率和干物重與產量嶺回歸方程的決定系數范圍分別為0.0219—0.3961、0.0452—0.1379和0.0914—0.6694,氮低效品種分別為0.0084—0.6190、0.1224—0.4341和0.0818—0.4881,產量與氮素吸收利用的關系年度間重演性較差,無明顯共性規律;以根干重、不定根數量、長度、表面積、體積、粗分枝根長度、表面積、體積以及細分枝根長度、表面積、體積11項根系形態指標為自變量,氮素積累量、氮素干物質生產效率和干物重三者分別為因變量進行嶺回歸分析,氮高效品種的回歸方程決定系數范圍分別為0.0527—0.2728、0.0653—0.3139和0.0714—0.3158,氮低效品種分別為0.0607—0.5040、0.0555—0.4411和0.0724—0.5449,氮素吸收利用與根系形態的關系規律年度間存在較大差異;2013—2015年,氮高效和氮低效品種抽穗期根系形態與產量嶺回歸方程的決定系數均超過0.8,<0.001,表明抽穗期根系形態對產量具有顯著影響是共性規律。(2)在以抽穗期根系形態指標為自變量,產量為因變量的嶺回歸方程中,氮高效品種的粗分枝根長度標準回歸系數最高,對產量影響最大;對氮低效品種而言,細分枝根表面積的增加對產量提高最有利。(3)常規施氮量(150 kg·hm-2)下,優化施肥模式為穗肥占比40%,且保花肥的比例達到或超過50%,其結合交替灌溉,有利于優化根系形態;SPAD指導施肥模式下,施氮量較優化施肥減少(由150 kg·hm-2降為120 kg·hm-2),采用常規灌溉比交替灌溉更有利于根系形態優化。【結論】水稻抽穗期根系形態與產量關系極為密切,合理的水氮管理措施能夠優化根系形態提高產量。常規灌溉結合SPAD指導施肥或交替灌溉結合優化施肥均有利于氮高效品種抽穗期粗分枝根長度的增長和氮低效品種細分枝根表面積的增大,最終提高產量。
氮效率;水稻;根系形態;產量;嶺回歸分析
0 引言
【研究意義】水稻根系是土壤養分的直接利用者和地上部光合生產主要的養分供應者,其形態和生理特征與地上部生長發育和產量形成有密切關系。氮素是水稻生長發育最重要的元素之一,水稻體內氮素主要由根系從土壤中吸收[1-2]。分別以根系和產量為出發點和落腳點,以氮素吸收利用為紐帶,進行水稻根系性狀和氮素吸收利用與產量關系的定量研究,有助于深入剖析根系在水稻產量形成過程中發揮作用的機理。【前人研究進展】圍繞水稻根系與氮素吸收和利用的關系,及其對產量形成的影響,前人已經開展了比較系統的研究,從水稻根系對氮素吸收利用的基因型差異[3-7],到根系對氮肥調控的響應[8-10],再到農藝措施對根系的影響及其與地上部生長及產量形成的關系[11-12]等方面均有涉及,這些研究在根系形態特征、生理活性、基因表達等層面取得了豐碩的理論成果,同時針對性地提出了一系列栽培技術措施,促進了水稻氮肥利用效率和產量的同步提高。【本研究切入點】已有研究多以包含較少變量的環境控制性重復試驗為樣本,通過簡單相關分析或普通最小二乘法回歸分析,獲得水稻根系形態和氮素吸收利用與產量的關系。這種研究策略針對性較強,得到的結果對特定條件下水稻生產的指導價值較大,但由于重復試驗工作量大,故這類研究包含的控制變量普遍較少,導致最終的試驗結果存在理論局限性較大,技術可移植性不高的問題。同時在統計分析中由于未對研究對象內部(自變量間)的共線性問題予以足夠重視,使得試驗結果中變量間關系的穩定性較差,加劇了不同研究結果的不一致性[6-7, 13]。針對上述研究不足,擬從不同試驗組成的包含較多變量因子的樣本中探求水稻根系形態、氮素吸收利用和產量三者之間的關系,與少變量重復試驗樣本相比,從多變量非重復試驗樣本中發掘三者關系的共性規律難度更大,但若三者的關系在排除共線性的干擾后仍能夠呈現出共性規律,則該研究結果的生產指導價值將更加廣泛。【擬解決的關鍵問題】本研究以課題組前期篩選出的在四川盆地稻區廣泛種植的氮高效和氮低效品種為試驗材料[14-16],通過3年大田試驗,探究水稻根系形態、氮素吸收和利用、產量形成三者之間關系的規律,以及實現高產的根系特征及相應水氮調控措施,為水稻高產高效育種及栽培提供理論依據及技術參考。
1 材料與方法
1.1 試驗地點
試驗于2013—2015年在四川農業大學水稻研究所試驗農場(30°43’N,103°47’E)進行。試驗點地處成都平原,屬亞熱帶濕潤氣候區。2006—2015年水稻生長季氣象資料來自四川省氣象局(圖1)。試驗地前茬為油菜,土壤質地為砂壤土,耕層土壤養分含量見表1。
1.2 供試材料
本研究以課題組前期篩選出的在農學利用率上有顯著差異的品種為供試材料。2013年和2014年為氮高效品種川農優498和氮低效品種川優6203[16],2015年為氮高效品種德香4103和氮低效品種宜香3724[14-15]。4個品種均為中秈遲熟雜交稻,生育期150 d左右。
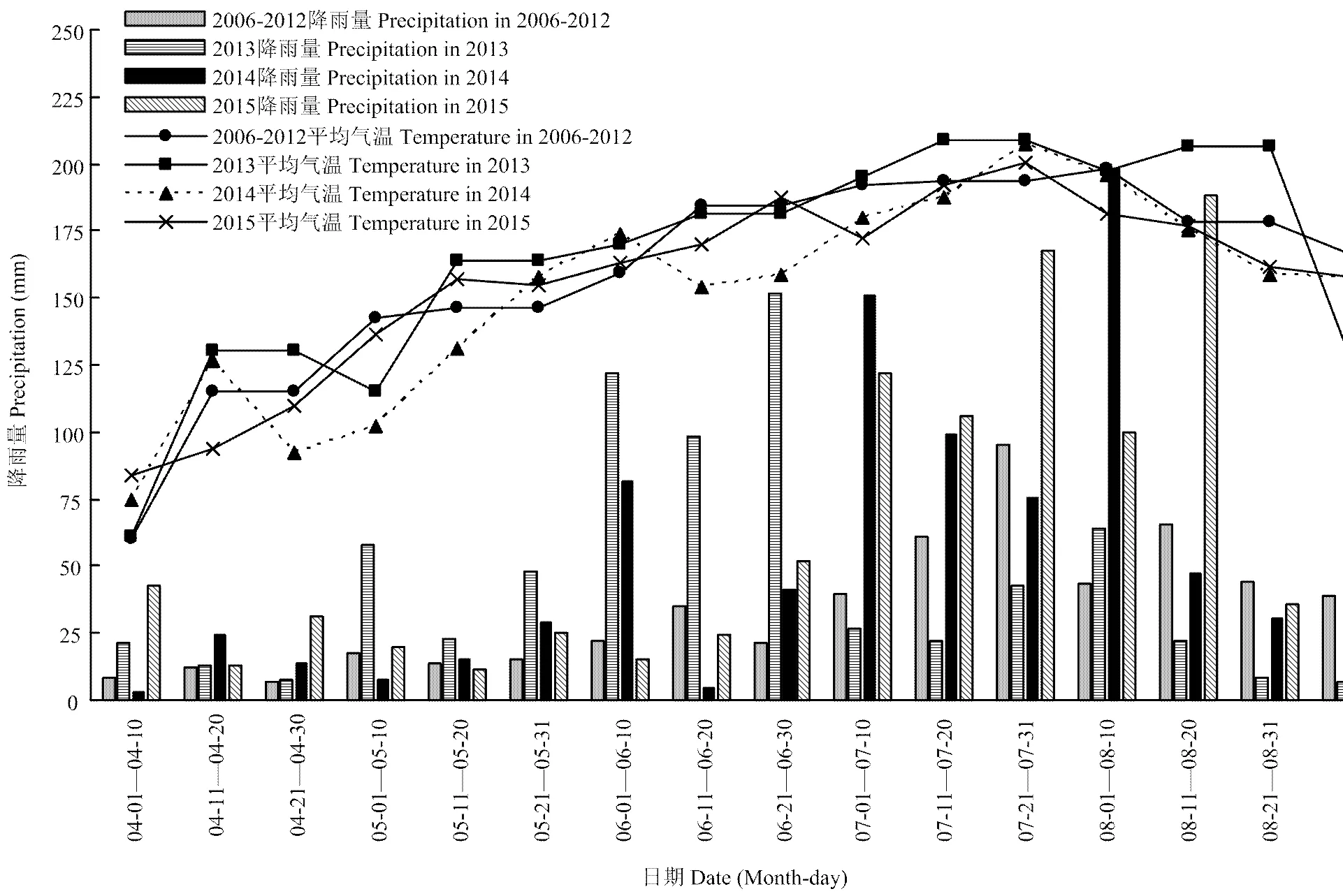
圖1 2006—2015年水稻生長季平均氣溫和降雨量

表1 2013—2015年耕層土壤養分含量表
1.3 試驗設計
2013年試驗采用裂區設計,品種為主區,氮肥運籌為副區,設基蘗肥與穗肥配比為B1(90﹕10)、B2(75﹕25)、B3(60﹕40)、B4(45﹕55)、B5(30﹕70),共計10個處理,3次重復,施氮量為150 kg·hm-2。
2014年試驗為裂裂區設計,品種為主區,每穴苗數為副區,設單苗(S)和雙苗(D)2個水平,穗肥氮運籌為裂裂區,設促花肥與保花肥配比為P1(40﹕0)、P2(20﹕20)、P3(0﹕40),共計12個處理,3次重復,施氮量為150 kg·hm-2,其中穗肥占比40%。
2015年試驗為裂裂區設計,品種為主區,水分管理為副區,設常規灌溉(W1)和交替灌溉(W2)2個水平[17],氮肥管理為裂裂區,設SPAD指導施肥(N1)、優化施肥(N2)及農民習慣施肥(N3)3種模式(表2),共計12個處理,3次重復。
2013—2015年試驗中,基肥(包含75 kg·hm-2磷肥和150 kg·hm-2鉀肥)、蘗肥、促花肥和保花肥分別于移栽前1 d、移栽后7 d、幼穗分化期(倒四葉)和抽穗前(倒二葉)施用。
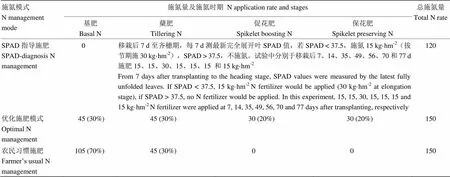
表2 2015年氮肥施用量及施肥時期
括號內百分數代表該日期氮肥施入量占總施氮量的比例 Values in the brackets indicate the percentage of N fertilizer applied to total N rate
1.4 測定項目與方法
1.4.1 根系形態指標測定 分別于拔節期、抽穗期和成熟期,每小區按平均莖蘗數標記5株具有代表性稻株,采用原狀土柱法,用鐵板取根器以稻株為中心掘取長等于行距(33.3 cm),寬等于株距(16.7 cm),深20 cm的土柱,裝入孔徑為0.4 mm的尼龍網袋中,浸泡6 h后洗去泥土雜質,獲得單株完整根系。用Epson Expression 10000XL掃描后,采用WinRHIZO Prov.2009c軟件分析不定根、粗分枝根、細分枝根各形態參數,而后置于80℃下烘干至恒重,測得根系總干重。不定根(0.3 mm<D≤1.65 mm)、粗分枝根(0.1 mm<D≤0.3 mm)、細分枝根(D≤0.1 mm)的界定參照顧東祥等[18]的方法。
1.4.2 氮素積累測定 將根系形態指標測定剩余的地上部,分莖、葉、穗各器官烘干,稱質量,再粉碎、過篩,用凱氏定氮儀(FOSS—8400)測定各器官的全氮含量。氮素干物質生產效率(nitrogen dry matter production efficiency,NDMPE)用單位氮素生產的干物質量表示。
1.4.3 考種與計產 成熟期各小區單獨收割,按實收株數計產。
1.5 數據處理
方差膨脹因子(variance inflation factor,VIF)用于診斷變量間存在的共線性關系,當某組變量的VIF平均值大于1且最大值大于10時,認為該組變量共線性嚴重。本研究用SPSS 20計算自變量的方差膨脹因子,結果顯示自變量間共線性嚴重(表3),符合進行嶺回歸分析的條件。嶺回歸分析(ridge regression)是一種用于共線性數據分析的有偏估計回歸,通過在自變量信息矩陣的主對角線元素上加入一個非負因子(嶺回歸參數k),使回歸系數的估計稍有偏差,但估計的穩定性明顯提高。本研究中當k=0.2時各自變量的嶺跡都基本穩定,因此嶺參數都取k=0.2。本研究用DPS 7.05進行嶺回歸分析和多重比較,用Excel 2003進行圖表制作。
2 結果
2.1 根系形態與氮素吸收利用及物質積累的嶺回歸分析
氮高效品種的產量、氮積累量、氮素干物質生產效率及成熟期干物重均顯著高于氮低效品種(表4)。二者根系形態與氮素吸收利用及物質積累的嶺回歸分析顯示,54組樣本中僅有2組決定系數超過0.5。2015年氮低效品種在拔節期的根系形態對氮素積累量和干物重變化的影響程度分別達到0.5040和0.5449,但在2013年和2014年對應樣本的方程決定系數均不超過0.2(表5)。這表明水稻根系形態與氮素積累量、氮素干物質生產效率及干物重關系的規律性較差。
表3 自變量方差膨脹因子
HN:氮高效;LN:氮低效;JS:拔節期;HS:抽穗期;MS:成熟期;NA:氮素積累;NDMPE:氮素干物質生產效率;DW:干物質量。下同
HN: High N use efficiency; LN: Low N use efficiency; JS: Jointing stage; HS: Heading stage; MS: Maturity stage; NA: N accumulation; NDMPE: N dry matter production efficiency; DW: Dry matter weight. The same as below

表4 氮高效和氮低效品種產量和氮素吸收利用比較
同列數據后不同字母表示在5 %水平差異顯著。下同
Values within a column followed by different letters are significantly different at<0.05. The same as below
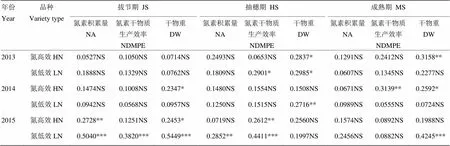
表5 根系形態與氮素吸收利用及物質積累的嶺回歸分析
*, **, ***分別表示在0.05, 0.01, 0.001水平差異顯著;NS表示二者差異未達顯著水平。下同
*, ** and *** indicate significance at 0.05, 0.01 and 0.001 levels, respectively; NS means that the difference was not significant. The same as below
2.2 各生育時期氮素吸收利用及物質積累與產量的嶺回歸分析
表6表明,氮高效和氮低效水稻在拔節期、抽穗期和成熟期的氮素積累量、氮素干物質生產效率及干物重與產量嶺回歸分析方程的決定系數大多低于0.5,僅有的決定系數超過0.6的3組(共計18組)樣本在年份間或品種間的重演性均較差。2013年和2014年氮高效品種成熟期的氮素積累量、氮素干物質生產效率及干物重對產量的影響程度分別為0.6694和0.6596,其中成熟期干物重對產量影響最大,氮素干物質生產效率次之,氮素積累量最小,但2015年嶺回歸方程的決定系數僅為0.0914,且干物重對產量的影響遠小于氮素干物質生產效率和氮素積累量。2015年氮低效品種在拔節期的氮素積累量、氮素干物質生產效率及干物重對產量的影響程度達到0.6190,但2013年和2014年嶺回歸方程的決定系數僅為0.0084和0.0680。這表明水稻拔節期、抽穗期及成熟期的氮素吸收、利用及生物量對產量的影響可能具有較強的品種特異性或受氣象條件影響較大,因而相互關系的規律重演性較差。
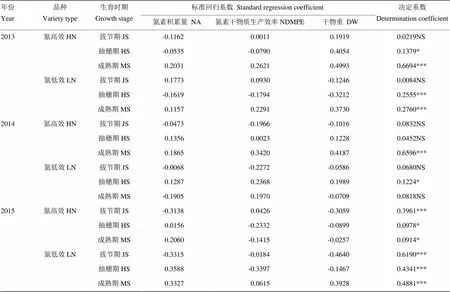
表6 產量與各生育時期氮素吸收利用及物質積累的嶺回歸分析
2.3 抽穗期根系形態與產量的嶺回歸分析
以水稻拔節期、抽穗期及成熟期的根系形態為自變量與產量作嶺回歸分析,結果顯示不同氮效率水稻均表現為抽穗期根系形態結構特征與產量關系的密切程度遠大于拔節期、成熟期,且該結果在3年大田試驗中重演性較好(數據未列出),本文僅對抽穗期根系形態與產量的關系進行分析(表7)。6組樣本的方程決定系數均超過0.8,達到極顯著水平,2013年和2014年氮高效品種抽穗期根系形態結構對產量的解釋程度均高于氮低效品種,2015年二者嶺回歸方程的決定系數相近。從各根系形態指標分析,氮高效品種的粗分枝根長度與產量關系最密切,氮低效品種的細分枝根表面積對產量的影響居所有指標之首。

表7 抽穗期根系形態與產量的嶺回歸分析
TRW:根干重。下同 TRW: Total root dry weight. The same as below
2.4 水肥管理對抽穗期根系形態的影響
2.4.1 氮肥運籌對抽穗期根系形態的影響 由表8可見,氮高效水稻抽穗期根系形態指標較氮低效水稻平均高21.81%—23.08%。隨著穗肥占比升高,根系生物量呈逐漸增大趨勢,穗肥占比超過40%,根系干重變化較小。穗肥占比提高促進了不定根數量、長度、表面積及體積的增長,穗肥占比超過40%,氮高效品種不定根數量及表面積等指標呈小幅下降趨勢,氮低效品種基本保持穩定。不定根上分出的粗分枝根以及后者分出的細分枝根也延續了不定根對氮肥運籌的響應規律,大都在B3處理時達到最大值。
2.4.2 穗肥氮運籌及每穴苗數對抽穗期根系形態的影響 氮高效品種抽穗期根系指標較氮低效品種平均高21.76%—22.18%,單苗或雙苗栽插對根系形態影響較小(表9)。施用保花肥能顯著提高根系生物量,但不同保花肥施用比例間差異較小。不定根的數量、長度、表面積和體積及其上分出的粗分枝根和細分枝根的各項指標也基本符合根系干重對穗肥運籌的響應規律。施用保花肥使氮高效品種粗分枝根長度和氮低效品種細分枝根表面積分別增加5.21%—20.78%和12.19 %—16.74 %。
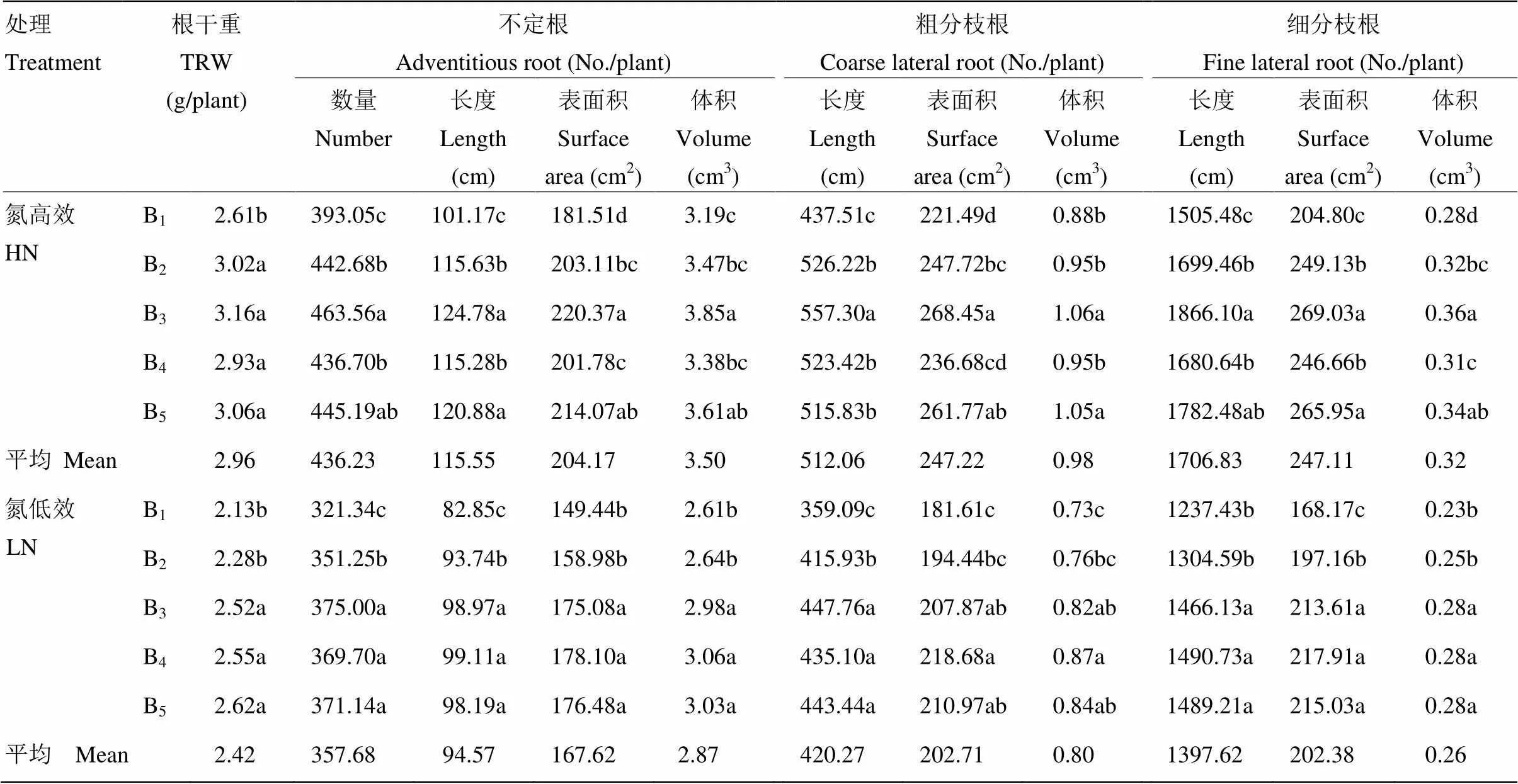
表8 氮肥運籌對抽穗期根系形態的影響(2013年)
B1:基蘗肥﹕穗肥=90 : 10;B2:基蘗肥﹕穗肥=75 : 25;B3:基蘗肥﹕穗肥=60 : 40;B4:基蘗肥﹕穗肥=45 : 55;B5:基蘗肥﹕穗肥=30 : 70
B1: Basal - tillering N : panicle N = 90 : 10; B2: Basal - tillering N : panicle N = 75 : 25; B3: Basal - tillering N : panicle N = 60 : 40; B4: Basal - tillering N : panicle N =45 : 55; B5: Basal - tillering N : panicle N = 30 : 70
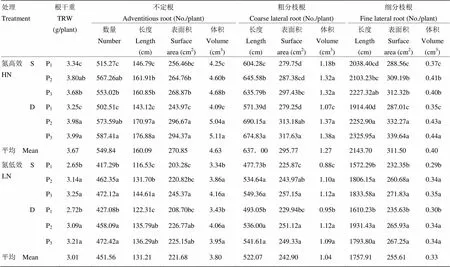
表9 穗肥氮運籌及每穴苗數對抽穗期根系形態的影響(2014年)
S:單苗;D:雙苗;P1:促花肥﹕保花肥=40 : 0;P2:促花肥﹕保花肥=20 : 20;P3:促花肥﹕保花肥=0 : 40
S: Single seedling; D: Double seedlings; P1: The ratio of nitrogen dressing at 4thand 2ndleaf from top stretching was 40 : 0; P2: The ratio of nitrogen dressing at 4thand 2ndleaf from top stretching was 20 : 20; P3: The ratio of nitrogen dressing at 4thand 2ndleaf from top stretching was 0 : 40
2.4.3 水氮管理對抽穗期根系形態的影響 表10表明,無論在常規灌溉還是交替灌溉條件下,均以農民習慣施肥(N3)抽穗期根系形態指標最小。常規灌溉條件下SPAD指導施肥氮高效品種根系形態指標均顯著高于優化施肥,采用SPAD指導施肥或優化施肥對氮低效品種根系形態指標影響差異較小。交替灌溉條件下,氮高效品種優化施肥比SPAD指導施肥在根系形態指標上更具優勢,而氮低效品種采用SPAD指導施肥更有利根系生長發育。對氮高效品種而言,常規灌溉下采用SPAD指導施肥和交替灌溉下采用優化施肥對粗分枝根長度增長更有利,比農民習慣施肥分別提高35.87%和36.26%;對氮低效品種而言,常規灌溉下SPAD指導施肥或交替灌溉下優化施肥均能有效促進細分枝根表面積增加,較農民習慣施肥分別提高29.91%和28.98%。
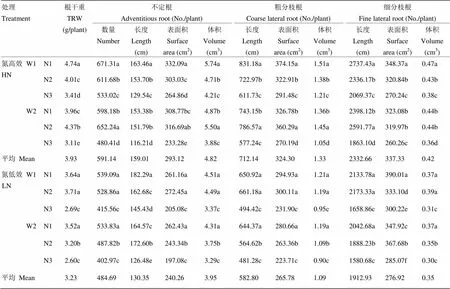
表10 水氮管理對抽穗期根系形態的影響(2015年)
W1:常規灌溉,W2:交替灌溉;N1、N2、N3分別代表SPAD指導施肥、優化施肥模式、農民習慣施肥
W1: Conventional irrigation, W2: Controlled alternate irrigation; N1: SPAD-diagnosis N management; N2: Optimal N management; N3: Farmer’s usual N management
3 討論
3.1 根系形態與氮素吸收利用的關系
Cassman等[6]研究表明氮高效水稻在根系生物量、體積、總吸收表面積和活性吸收面積方面均具有較大優勢。戢林等[3]研究認為水稻粗分枝根的發育情況會直接影響氮素的吸收,進而影響產量和氮素利用效率。Samejima等[7, 19]和石慶華等[13]研究認為部分在根系形態、生理活性方面較優的品種,在氮素吸收利用率方面并未表現明顯優勢。本研究水稻根系形態與氮素吸收利用嶺回歸方程的決定系數波動范圍較大(0.0527≤2≤0.5449),且相同指標在不同試驗中表現也差異較大,這反映了水稻根系形態與氮素吸收利用關系的復雜性。首先,水稻根系吸收養分的過程中,氮、磷、鉀等元素存在顯著地協同或拮抗作用,而磷等對根系生長發育具有較強的調節作用[20-22];其次,根系吸收養分的同時會以根泌的方式釋放出不同類型化合物,反饋調節根系生長發育及對氮、磷、鉀等的吸收利用[23];因此土壤性狀、肥水管理等都可能對根系形態與氮素吸收利用的關系產生重要影響。本研究中,不同試驗間的氣象條件、土壤性狀及水氮管理等均存在一定差異,這可能是根系性狀與氮素吸收利用之間關系在3個試驗中無共性規律的主要原因。
3.2 氮素吸收利用及物質積累與產量的關系
氮素干物質生產效率是衡量水稻氮素利用效率的重要指標之一[24],過量施氮或穗肥占比過高易導致植株氮積累尤其是花后氮積累偏高,削弱其碳同化能力,使氮素干物質生產效率降低[25]。Chen等[26]研究認為不同氮效率水稻的氮素籽粒生產效率及氮素干物質生產效率差異并不顯著,氮高效品種發揮產量優勢的關鍵在于吸收更多的氮素。Qiao等[27]研究認為在高地力稻田,施氮雖能增加植株氮積累量,但較多氮素會滯留在秸稈內,增產效果較差。本研究顯示在決定系數較高的嶺回歸方程里,氮素吸收和利用對產量的影響較小,普遍低于生物量,與前人研究存在一定差異,其原因可能在兩方面:首先,四川盆地稻田土壤基礎肥力高,對產量貢獻大,屬地力貢獻率高值區,施肥發揮的作用較小[28-29]。其次,四川盆地晝夜溫差小、光照時數少,水稻產量潛力小。本研究中絕大多數處理產量已經超過該地區產量潛力估值,此時氮素可能已經不是產量的決定因素[30-31]。
2013年和2014年試驗氮高效品種干物重對產量的影響遠大于2015年試驗,除了品種間的基因型差異外,氣象條件的差異亦可能是重要原因。2015年試驗水稻灌漿期降水頻繁,日照時數較近十年平均值減少27%,較2013年和2014年分別減少35%和29%,光照不足限制了水稻群體花后光合生產能力,增強了花前物質輸出對產量的影響,導致生物量對產量的影響減弱[32]。
3.3 抽穗期根系形態與產量的關系
目前研究普遍認為抽穗期水稻根系已經完成生長且對產量影響最大[33-34]。本研究顯示抽穗期根系形態對產量的影響在所有樣本中均較大,表明該時期根系形態與產量的密切關系具有普遍性。就常見形態指標來說,不定根數量、根系吸收表面積及根直徑等對產量影響較大,采用根系分析系統細化到分枝根層面,則是粗分枝根長度、表面積及細分枝根表面積等與產量關系密切[3, 35]。本研究獲得相近的結果,即粗分枝根長度和細分枝根表面積分別對氮高效和氮低效品種產量影響較大。不同氮效率品種間的差異可能與二者對養分的需求差異有關,氮高效品種更高產量的實現需要較多的養分支撐,粗分枝根數量增加有利于擴展根系吸收空間,增加土壤養分供應量[3]。
根系對土壤環境的適應具有可塑性,合理的水氮管理能夠通過與土壤互作優化根系形態提高產量,在此過程中分枝根的表現尤為突出[10,18,36-38]。本研究中,合理施氮(基蘗肥﹕促花肥﹕保花肥= 60﹕20﹕20)和水氮耦合管理(SPAD指導施肥采用常規灌溉、優化施肥結合交替灌溉)能夠優化抽穗期氮高效品種粗分枝根長度和氮低效品種細分枝根表面積,實現產量增長。
4 結論
水稻根系形態對產量具有重要影響,尤其是抽穗期根系形態與產量關系極為密切。合理施氮與控水可優化各類型根的生長與分布特征,但需考慮不同品種之間的差異,促進抽穗期粗分枝根長度及細分枝根表面積增長分別是實現氮高效和氮低效品種根系形態優化的首要目標。常規施氮量(150 kg·hm-2)下,穗肥比例為40%,且其中保花肥的占比達到或超過50%,同時結合交替灌溉,有利于優化根系形態;在SPAD指導施肥模式(施氮量為120 kg·hm-2)下,采用常規灌溉更有利于根系形態優化。
References
[1] PENG S B, HUANG J L, ZHONG X H, YANG J C, WANG G H, ZOU Y B, ZHANG F S, ZHU Q S, ROLAND B, CHRISTIAN W.Challenge and opportunity in improving fertilizer-nitrogen use efficiency of irrigated rice in China., 2002, 1(7): 776-785.
[2] 蔡昆爭, 駱世明, 段舜山. 水稻根系在根袋處理條件下對氮養分的反應. 生態學報, 2003, 23(6): 1109-1116.
CAI K Z, LUO S M, DUAN S S. The response of the rice root system to nitrogen conditions under root confinement., 2003, 23(6): 1109-1116. (in Chinese)
[3] 戢林, 李廷軒, 張錫洲, 余海英. 氮高效利用基因型水稻根系形態和活力特征. 中國農業科學, 2012, 45(23): 4770-4781.
JI L, LI T X, ZHANG X Z, YU H Y. Root morphologyical and activity characteristics of rice genotype with high nitrogen utilization efficiency.2012, 45(23): 4770-4781. (in Chinese)
[4] 魏海燕, 張洪程, 張勝飛, 杭杰, 戴其根, 霍中洋, 許軻, 馬群, 張慶, 劉艷陽. 不同氮利用效率水稻基因型的根系形態與生理指標的研究. 作物學報, 2008, 34(3): 429-436.
WEI H Y, ZHANG H C, ZHANG S F, HANG J, DAI Q G, HUO Z Y, XU K, MA Q, ZHANG Q, LIU Y Y.Root morphological and physiological characteristics in rice genotypes with different N use efficiencies.2008, 34(3): 429-436. (in Chinese)
[5] 李敏, 張洪程, 楊雄, 葛夢婕, 馬群, 魏海燕, 戴其根, 霍中洋, 許軻, 曹利強, 吳浩. 水稻高產氮高效型品種的根系形態生理特征. 作物學報, 2012, 38(4): 648-656.
LI M, ZHANG H C, YANG X, GE M J, MA Q, WEI H Y, DAI Q G, HUO Z Y, XU K, CAO L Q, WU H. Root morphological and physiological characteristics of rice cultivars with high yield and high nitrogen use efficiency.2012, 38(4): 648-656. (in Chinese)
[6] CASSMAN K G, DOBERMANN A, WALTERS D T. Agroecosystems, nitrogen-use efficiency, and nitrogen management.,2002, 31(2): 132-140.
[7] SAMEJIMA H, KONDO M, ITO O, NOZOE T, SHINANO T, OSAKI M. Characterization of root systems with respect to morphological traits and nitrogen-absorbing ability in the new plant type of tropical rice lines.2005, 28(5): 835-850.
[8] 樊小林, 史正軍, 吳平. 水肥(氮)對水稻根構型參數的影響及其基因型差異. 西北農林科技大學學報(自然科學版), 2002, 30(2): 1-5.
FAN X L, SHI Z J, WU P.Effects of nitrogen fertilizer on parameters of rice (L.) root architecture and their genotypic difference., 2002, 30(2): 1-5. (in Chinese)
[9] 孫虎威, 王文亮, 劉尚俊, 候蒙蒙, 謝天寧, 粱志浩, 樊亞男, 張亞麗. 低氮脅迫下水稻根系的發生及生長素的響應. 土壤學報, 2014, 51(5): 1096-1102.
SUN H W, WANG W L, LIU S J, HOU M M, XIE T N, LIANG Z H, FAN Y N, ZHANG Y L. Formation of rice root regulated by nitrogen dificiency.2014, 51(5): 1096-1102. (in Chinese)
[10] WALCH-LIU P, IVANOV I, FILLEUR S Y, GAN Y, REMANS T, FORDE B G. Nitrogen regulation of root branching.2006, 97(5): 875-881.
[11] 楊志遠, 孫永健, 徐徽, 秦儉, 賈現文, 馬均. 栽培方式與免耕對雜交稻Ⅱ優498灌漿期根系衰老和籽粒灌漿的影響. 中國農業科學, 2013, 46(7): 1347-1358.
YANG Z Y, SUN Y J, XU H, QIN J, JIA X W, MA J. Influence of cultivation methods and no-tillage on root senescence at filling stage and grain-filling properties of Eryou 498., 2013, 46(7): 1347-1358. (in Chinese)
[12] 張洪程, 朱聰聰, 霍中洋, 許軻, 蔣曉鴻, 陳厚存, 高尚勤, 李德劍, 趙成美, 戴其根, 魏海燕, 郭保衛. 缽苗機插水稻產量形成優勢及主要生理生態特點. 農業工程學報, 2013, 29(21): 50-59.
ZHANG H C, ZHU C C, HUO Z Y, XU K, JIANG X H, CHEN H C, GAO S Q, LI D J, ZHAO C M, DAI Q G, WEI H Y, GUO B W.Advantages of yield formation and main characteristics of physiological and ecological in rice with nutrition bowl mechanical transplanting.2013, 29(21): 50-59. (in Chinese)
[13] 石慶華, 李木英, 涂起紅. 雜交水稻根系N素營養效率及其生理因素研究. 雜交水稻, 2002, 17(4): 49-52.
SHI Q H, LI M Y, TU Q H. Studies on efficiency of N nutrition and physiological factors in roots of hybrid rice.2002, 17(4): 49-52. (in Chinese)
[14] 秦儉, 楊志遠, 孫永健, 馬均. 不同穗型雜交秈稻物質積累、氮素吸收利用和產量的差異比較. 中國水稻科學, 2014, 28(5): 514-522.
QIN J, YANG Z Y, SUN Y J, MA J. Differential comparison of assimilation products accumulation, nitrogen uptake and utilization and grain yield of hybridrice combinations with different panicle types.,2014, 28(5): 514-522. (in Chinese)
[15] 朱從樺, 孫永健, 嚴奉君, 蔣明金, 徐徽, 趙建紅, 馬均. 曬田強度和氮素穗肥運籌對不同氮效率雜交稻產量及氮素利用的影響. 中國水稻科學, 2014, 28(3): 258-266.
ZHU C H, SUN Y J, YAN F J, JIANG M J, XU H, ZHAO J H, MA J. Effects of different paddy field drainage degrees and panicle nitrogen fertilizer managements on yield and nitrogen utilization of different nitrogen efficiency hybrid rice.2014, 28(3): 258-266. (in Chinese)
[16] 楊世民, 楊志遠, 孫永健, 馬均. 氮肥運籌對2個不同穗重型雜交稻產量及氮肥利用的影響. 浙江大學學報(農業與生命科學版), 2015, 41(6): 685-694.
YANG S M, YANG Z Y, SUN Y J, MA J. Effects of nitrogen on grain yield and nitrogen use efficiency of two hybrid rice varieties with different panicle masses.2015, 41(6): 685-694. (in Chinese)
[17] 孫永健, 馬均, 孫園園, 徐徽, 嚴奉君, 代鄒, 蔣明金, 李玥. 水氮管理模式對雜交秈稻岡優527群體質量和產量的影響. 中國農業科學, 2014, 47(10): 2047-2061.
SUN Y J, MA J, SUN Y Y, XU H, YAN F J, DAI Z, JIANG M J, LI Y. Effects of water and nitrogen management patterns on population quality and yield of hybrid rice Gangyou 527.,2014, 47(10): 2047-2061. (in Chinese)
[18] 顧東祥, 湯亮, 徐其軍, 雷曉俊, 曹衛星, 朱艷. 水氮處理下不同品種水稻根系生長分布特征. 植物生態學報, 2011, 35(5): 558-566.
GU D X, TANG L, XU Q J, LEI X J, CAO W X, ZHU Y. Root growth and distribution in rice cultivars as affected by nitrogen and water supply.2011, 35(5): 558-566. (in Chinese)
[19] SAMEJIMA H, KONDO M, ITO O, NOZOE T, SHINANO T. Root-shoot interaction as a limiting factor of biomass productivity in new tropical rice lines.2004, 50(4): 545-554.
[20] 許飛云, 張茂星, 曾后清, 朱毅勇. 水稻根系細胞膜質子泵在氮磷鉀養分吸收中的作用. 中國水稻科學, 2016, 30(1): 106-110.
XU F Y, ZHANG M X, ZENG H Q, ZHU Y Y. Involvement of plasma membrane H+-ATPase in uptake of nitrogen, phosphorus and potassium by rice root.2016, 30(1): 106-110. (in Chinese)
[21] DINGKUHN M, LUQUET D, KIM H K, TAMBOUR L, CLEMENTVIDAL A. EcoMeristem, a model of morphogenesis and competition among sinks in rice. 2. Simulating genotype responses to phosphorus deficiency.,2006, 33(4): 325-337.
[22] INSALUD N, BELL R W, COLMER T D, RERKASEM B. Morphological and physiological responses of rice () to limited phosphorus supply in aerated and stagnant solution culture.2006, 98(5): 995-1004.
[23] LI M, ZHANG H C, YANG X, GE M J, MA Q, WEI H Y, DAI Q G, HUO Z Y, XU K, LUO D Q. Accumulation and utilization of nitrogen, phosphorus and potassium of irrigated rice cultivars with high productivities and high N use efficiencies.2014, 161: 55-63.
[24] CASSMAN K G, Peng S, OLK D C, LADHA J K, REICHARDT W, DOBERMANN A, SINGH U. Opportunities for increased nitrogen- use efficiency from improved resource management in irrigated rice systems.1998, 56(1): 7-39.
[25] JIANG L G, DAI T B, JIANG D, CAO W X, GAN X Q, WEI S Q. Characterizing physiological N-use efficiency as influenced by nitrogen management in three rice cultivars.2004, 88(2/3): 239-250.
[26] CHEN G, CHEN Y, ZHAO G H, CHENG W D, GUO S W, ZHANG H L, SHI W M. Do high nitrogen use efficiency rice cultivars reduce nitrogen losses from paddy fields?2015, 209: 26-33.
[27] QIAO J, YANG L Z, YAN T M, XUE F, ZHAO D. Nitrogen fertilizer reduction in rice production for two consecutive years in the Taihu Lake area.2012, 146(1): 103-112.
[28] 湯勇華, 黃耀. 中國大陸主要糧食作物地力貢獻率和基礎產量的空間分布特征. 農業環境科學學報. 2009, 28(5): 1070-1078.
TANG Y H, HUANG Y. Spatial distribution characteristics of the percentage of soil fertility contribution and its associated basic crop yield in Mainland China., 2009, 28(5): 1070-1078. (in Chinese)
[29] 梁濤, 陳軒敬, 趙亞南, 黃興成, 李鴻, 石孝均, 張躍強. 四川盆地水稻產量對基礎地力與施肥的響應. 中國農業科學, 2015, 48(23): 4759-4768.
LIANG T, CHEN X J, ZHAO Y N, HUANG X C, LI H, SHI X J, ZHANG Y Q. Response of rice yield to inherent soil productivity of paddies and fertilization in Sichuan basin, 2015, 48(23): 4759-4768. (in Chinese)
[30] VAN WART J, KERSEBAUM K C, Peng S B, MILNER M, CASSMAN K G. Estimating crop yield potential at regional to national scales.2013, 143(1): 34-43.
[31] DOBERMANN A. Fertilizer best management practices: General principles, strategy for their adoption and voluntary initiatives vs regulations//Proceedings of the IFA International Workshop on Fertilizer Best Management Practices. Paris: International Fertilizer Industry Association, 2007: 1-28.
[32] 楊志遠, 胡蓉, 孫永健, 徐徽, 許遠明, 馬均. 三角形強化栽培模式下氮肥運籌對II優498產量及氮肥利用的影響. 作物學報, 2012, 38(6): 1097-1106.
YANG Z Y, HU R, SUN Y J, XU H,XU Y M, MA J. Effects of nitrogen fertilizer management on yield and nitrogen use efficiency of Eryou 498 in triangle-planted system of rice intensification., 2012, 38(6): 1097-1106. (in Chinese)
[33] 劉桃菊, 戚昌瀚, 唐建軍. 水稻根系建成與產量及其構成關系的研究. 中國農業科學, 2002, 35(11): 1416-1419.
LIU T J, QI C H, TANG J J. Studies on relationship between the character parameters of root and yield formation in rice.2002, 35(11): 1416-1419. (in Chinese)
[34] 樊劍波, 沈其榮, 譚炯壯, 葉利庭, 宋文靜, 張亞麗. 不同氮效率品種根系生理生態指標的差異. 生態學報, 2009, 29(6): 3052-3058.
FAN J B, SHEN Q R, TAN J Z, YE L T, SONG W J, ZHANG Y L. Difference of root physiological and ecological indicies in rice cultivars with different N use efficiency.2009, 29(6): 3052-3058. (in Chinese)
[35] 李杰, 張洪程, 常勇, 龔金龍, 胡雅杰, 龍厚元, 戴其根, 霍中洋, 許軻, 魏海燕, 高輝. 高產栽培條件下種植方式對超級稻根系形態生理特征的影響. 作物學報, 2011, 37(12): 2208-2220.
LI J, ZHANG H C, CHANG Y, GONG J L, HU Y J, LONG H Y, DAI Q G, HUO Z Y, XU K, WEI H Y, GAO H. Influence of planting methods on root system morphological and physiological characteristics of super rice under high-yielding cultivation condition., 2011, 37(12): 2208-2220. (in Chinese)
[36] DE DORLODOT S, FORSTER B, PAGèS L, PRICE A, TUBEROSA R, DRAYE X. Root system architecture: Opportunities and constraints for genetic improvement of crops.2007, 12(10): 474-481.
[37] DAVIES W J. Root growth response and functioning as an adaptation in water limiting soils//JENKS M A, HASEGAWA P M, JAIN S M.. Dordrecht: Springer Netherlands, 2007: 55-72.
[38] 徐國偉, 呂強, 陸大克, 王賀正, 陳明燦. 干濕交替灌溉耦合施氮對水稻根系性狀及籽粒庫活性的影響. 作物學報, 2016, 42(10): 1495-1505.
XU G W, Lü Q, LU D K, WANG H Z, CHEN M C. Effect of wetting and drying alternative irrigation coupling with nitrogen application on root characteristic and grain-sink activity.2016, 42(10): 1495-1505. (in Chinese)
(責任編輯 楊鑫浩)
The Relationships Between Root Morphology, N Absorption and Utilization and Grain Yield in Rice with Different N Use Efficiencies
LI Na, YANG ZhiYuan, DAI Zou, SUN YongJian, XU Hui, HE Yan, YAN TianRong, JIANG MingJin, GUO ChangChun, WANG ChunYu, MA Jun
(Rice Research Institute, Sichuan Agricultural University/Key Laboratory of Crop Physiology, Ecology, and Cultivation in Southwest, Ministry of Agriculture, Chengdu 611130)
【Objective】The objective of this research is to study the relationships between root morphology, nitrogen (N) absorption and utilization and grain yield in rice with different N use efficiencies, clarify the root morphology characters of high yielding, and put forward the corresponding optimized water-N management practice. 【Method】In 2013, high N use efficiency (HN) variety Chuannongyou 498 and low N use efficiency (LN) variety Chuanyou 6203 were used as trial materials to conduct a split-plot experiment with different panicle fertilizer ratio. In 2014, a split-split plot experiment was performed with the same varieties, where seedling number per hole was assigned to the split plot, and the proportion of spikelet preserving fertilizer and spikelet promoting fertilizer was assigned to the split-split plot. In 2015, a split-split plot experiment was carried out with water management practice as split plot and N-management practice as split-split plot, where HN variety Dexiang 4103 and LN variety Yixiang 3724 were used as trial materials. Grain yield, root morphology characters, and N absorption and utilization characters, obtained from the above 3 field experiments, were performed multiple collinearity diagnostics through calculating the variance inflation factor. After confirming that there were serious multiple collinearity problems, ridge regression analysis was used as the main data analysis method to investigate the relationships between root morphology characters, N absorption and utilization and grain yield. Based on the recurrence of these relationships from 2013 to 2015, common regularities hidden among these relationships were located. 【Result】The results were as follows: (1) For HN varieties, the determination coefficients of ridge regression equation between grain yield and N accumulation (NA), N dry matter production efficiency (NDMPE), and dry matter weight (DW) at elongation stage, heading stage, and maturity stage, were in the range of 0.0219-0.3961, 0.0452-0.1379, and 0.0914-0.6694, respectively. For LN varieties, the determination coefficients were in the range of 0.0084-0.6190, 0.1224-0.4341, and 0.0818-0.4881, respectively. The relationship between grain yield and N uptake and utilization was less repeatable during the year, and there was no obvious common regularity. Ridge regression analysis was performed with 11 root morphology characters of total root dry weight, adventitious root number, length, surface area, volume, coarse lateral root length, surface area, volume, and fine lateral root length, surface area, volume as independent variables and NA, NDMPE, and DW as dependent variables, respectively. For HN varieties, the determination coefficients were in the range of 0.0527-0.2728, 0.0653-0.3139, and 0.0714-0.3158, respectively. For LN varieties, the determination coefficients were in the range of 0.0607-0.5040, 0.0555-0.4411, and 0.0724-0.5449, respectively. There were significant differences in the relationships between N absorption and utilization and root morphology among years. From 2013 to 2015, the determination coefficients of ridge regression equation between grain yield and root morphology characters of both HN varieties and LN varieties all exceeded 0.8 (<0.001), which showed an common regularity that root morphology at heading stage had a significant effect on grain yield. (2) In the ridge regression equation with root morphology characters as independent variables, and grain yield as dependent variable, the coarse lateral root length of HN varieties obtained the largest standard coefficient, and had the greatest impact on grain yield. However, for the LN varieties, the fine lateral root surface area played the most important role in increasing grain yield. (3) With N rate of 150 kg·hm-2, the optimal N management practice (panicle fertilizer ratio was 40% and the ratio of spikelet preserving fertilizer to panicle fertilizer reached or exceeded 50%) combined with dry-wet alternate irrigation was beneficial to optimize the root morphology. For the SPAD-diagnosis N management practice, N rate reduced from 150 kg·hm-2to 120 kg·hm-2, conventional irrigation mode was more favorable to root morphology optimization than dry-wet alternate irrigation.【Conclusion】Rice root morphology at heading stage had close relationship with grain yield, and appropriate water-N management practice could optimize rice morphology to increase grain yield. SPAD-diagnosis N management practice coupled with conventional irrigation or optimal N management combined with dry-wet alternate irrigation mode could extend the coarse lateral root length of HN varieties as well as the fine lateral root surface area of LN varieties at the heading stage to increase their grain yields, respectively.
N use efficiency; rice; root morphology; grain yield; ridge regression analysis
2016-11-09;接受日期:2017-03-07
國家重點研發計劃(2016YFD0300506)、國家糧食豐產科技工程(2013BAD07B13)
李娜,E-mail:lina1409bs@163.com。通信作者馬均,E-mail:majunp2002@163.com
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年14期)2021-10-14 08:35:40
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
