人工模擬降雨格局變化對紅砂種子萌發的影響
2017-10-13 11:29:16單立山張正中段桂芳種培芳
生態學報 2017年16期
關鍵詞:研究
單立山,李 毅,張正中,白 蕾,段桂芳,種培芳
甘肅農業大學林學院, 蘭州 730070
人工模擬降雨格局變化對紅砂種子萌發的影響
單立山,李 毅*,張正中,白 蕾,段桂芳,種培芳
甘肅農業大學林學院, 蘭州 730070
氣候變化改變降雨格局,會影響到種子出苗及幼苗生長,進而影響幼苗的更新動態。為探討降雨格局變化對典型荒漠植物紅砂種子萌發特性的影響,利用環境控制生長箱開展了降雨量和降雨間隔時間的雙因素控制實驗。結果表明:(1)總降雨量增加30%,紅砂種子出苗率和發芽勢分別平均提高45.69%、39.86% (P<0.05),延長降雨間隔時間單次降雨量達到6mm其效果更明顯,出苗率和發芽勢達到最大值,分別為68.33%、63.33%,表明6mm 降水量是促使紅砂萌發的最小降雨閾量。(2)總降雨量增加30%顯著提高了種子萌發指數和活力指數(P<0.05),與自然總降雨量相比,分別平均提高57.67%、121%。總降雨量減少30%雖降低了萌發指數和活力指數,但與自然總降雨量相比差異不顯著(P>0.05)。(3)降雨量增加30%延長降雨間隔時間處理加快了紅砂的萌發進程,縮短了種子的萌發持續時間,其萌發曲線較陡峭。降雨量減少30%對其種子萌發進程影響不大。該研究得到以下主要結論:1)紅砂種子萌發主要受到降雨量的影響,但降雨量效應依賴降雨間隔時間,總降雨量一定降雨間隔延長所形成的單次降雨量增加均提高萌發率,增加了紅砂繁殖成功率,對其幼苗更新起促進效應;2)在自然狀態及降雨減少的情況下紅砂種子具有推遲萌發的特性,使其幼苗在更適宜的環境條件下出現和生長的機會增多,從而提高植物對環境的長期適應性。
降雨間隔時間;降雨量;種子萌發;幼苗更新
Abstract: Seed germination is considered to be the most important and sensitive factor in response to changes in the moisture and heat phases during the natural process of plant regeneration. The northwest arid areas of China are drought areas that are most sensitive to precipitation changes.Reaumuriasoongorica, a super-xerophytic shrub, exhibits a strong tolerance to drought, cold, saline-alkali soil, and barren landscapes, and is a long-lived species. Our objective was to investigate how precipitation changes affect the physiological and ecological processes of seeds ofR.soongoricato determine the regeneration mechanisms and changes in the patterns of this key species. Seeds ofR.soongoricawere collected in the fall of 2013 at the Linze Inland River Basin Research Station of the Chinese Academy of Sciences. We set up a growth-chamber experiment with two factors: precipitation quantity (natural precipitation as a control, reduction of 30%, and increase of 30%) and interval (time elapsed between two precipitation events, 5 and 10 days). Germinated seeds were counted daily until the end of germination. The results showed that increasing precipitation (+30%) resulted in a significantly enhanced germination rate and germination potential by an average of 45.69% and 39.86%, respectively, especially when the precipitation interval was extended from 5 to 10 days (a single rainfall reached up to 6 mm). In addition, the germination rate and potential reached their maxima by an average of 68.33% and 63.33%, respectively. Therefore, the rainfall threshold for the germination ofR.soongoricawas determined to be 6 mm. The germination index and vigor index were significantly increased by an average of 57.67% and 121% (P<0.05), respectively, with increasing precipitation quantity (+30%). Extension of the precipitation interval under increasing precipitation (+30%) resulted in a reduced germination index and vigor index, but the effect was not statistically significant. Increasing precipitation (+30%) under the extended precipitation interval treatments accelerated the course of germination, and the germination periods were shorter. Reducing the precipitation by 30% had no significant effects on the course of germination. Therefore, seed germination was mainly affected by precipitation quantity, but the effect was dependent on the precipitation interval. Overall, the seed germination percentage increased under a precipitation pattern of increasing precipitation (+30%) with an extended precipitation interval, the breeding success rate and seedling establishment ability were enhanced, and seedling regeneration was promoted.R.soongoricashowed characteristics of delayed germination under the condition of natural rainfall and reduced precipitation(-30%), which could increase the seedling emergence and growth of the seedlings under further favorable environmental conditions and improve the long-term adaptability of plants to the environment.
KeyWords: precipitation interval; precipitation quantity; seed germination; seedling regeneration
全球氣候模型預測,降雨格局將發生改變,以間隔時間增大、單次降雨量增多為特征的降雨事件將會增加[1- 3]。干旱區降雨稀少,一般由高頻率、小于5mm的小雨量降雨組成,但降雨格局變異較大[4- 5],主要表現為,單次降雨量增加、干旱間隔期延長[3]。水是干旱荒漠植物種子萌發及生長的主要限制因子,降雨是其主要的水分來源[6- 7],降雨變化必將影響到種子萌發過程,進而對植物幼苗更新產生顯著的影響[8- 9]。種子萌發對降雨變化的響應以及其適應性特征會直接影響后續的幼苗建立與補充的成功與否,進而影響種群的更新動態[9]。因此,加強干旱區降雨格局變化對種子萌發影響的研究,對于預測全球氣候變化背景下干旱區植被的發展變化趨勢以及種質資源保育及撫育更新均具有重要意義。
在干旱和半干旱區,年降雨量是植被建立的主要限制因子,其變化對必將對其種子萌發出苗產生有顯著的影響[10]。近年來,降雨量對種子萌發出苗影響的研究已有大量報道,Zheng等[11]研究發現,在毛烏素沙地,當地許多本地種其萌發出苗隨降雨量的增加而增加,González-Astorga和Núez-Farfán[12]在墨西哥、Ambrose和Wilson[13]在加拿大大平原北部邊緣就不同降雨量條件下的Tagetesmicrantha、冰草(Agropyroncristatum)和格蘭馬草(Boutelouagracilis)進行了研究,得出了類似的結果。但有研究指出,春季降雨量適當的增加并不會改變種子出苗率,但會提前其出苗物候[14-15];而降雨量減少導致的土壤干旱化將降低種子的存活力及出苗率,阻礙幼苗的建立和補充,進而抑制其種群更新[16-18]。也有研究發現不同植物在相同降雨量條件下其種子萌發出苗表現出不同的策略,如降雨量增加1倍的條件下均可導致蒙古櫟(Quercusmongolica)種子的萌發率的下降;而降雨量增加1倍卻有利于紅松(Pinuskoraiensis)種子打破休眠和萌發[19]。然而,有研究指出總降雨量這單一因素并不能決定植物的種子萌發,而生長季內由降雨(包括生長季內的平均單次降雨量、降雨次數和平均降雨間隔時間3個因素)分布狀況對植物種子萌發出苗的影響力更大[20- 21]。近年來,降雨格局變化對種子萌發出苗的生態效應研究開始受到較多的關注,特別是降雨間隔時間變化顯得更重要。Zhu等[22]研究發現賴草(Leymussecalinus)在總降雨量一致的條件下,其種子出苗率隨降雨頻率的減少(降雨間隔時間的縮短單次降雨量減少)而減少;科爾沁沙地尖頭葉藜(ChenopodiumacuminatumWilld)在總降雨量一致的情況下,其種子萌發出苗也隨降雨頻率的減少而減少,8mm降雨量是促使尖頭葉藜萌發的最小降雨閾量[21]。可見,每次降雨量及降雨次數的分布狀況對種子萌發出苗顯得更重要。荒漠植物種子萌發所依賴的最重要的環境因素是降雨的分布和雨量[23]。Loria和Noy-Meir[24]研究發現,生長在以色列Negev沙漠齒稃草(Schismusarabicus),其種子能在冬天少于10 mm 的降雨條件下萌發,而在溫度很高、濕度很低的夏天,只有在大于90 mm 的人工灌溉條件下種子才能萌發,當雨量增加到200 mm 以上,且分配適當,出苗的數量和比率都較高。荒漠中一些種子的萌發不但依賴雨量,而且依賴雨水的次數[23],Juhren等[25]研究發現沙漠中生長的一年生植物,其幼苗數量由降雨的次數所控制;Boerhaviaspicata的種子在第一場雨后不萌發,而在第二場雨后才能萌發[26]。綜上所述,目前關于降雨格局變化對種子萌發出苗的研究還很有限,研究結果之間差異也很大,沒有規律性的結論,需要進行更加廣泛深入的研究,且以往的研究多集中在降雨量變化對種子萌發和出苗的影響,而就降雨頻率和強度對種子萌發和影響的研究相對較少,這妨礙了人們對種子萌發和出苗對未來降雨格局變化響應機理的深入探討。
紅砂[Reaumuriasoongorica(Pall.) Maxim]為我國西北荒漠地區優勢種、建群種。在干旱區,稀少、多變的降雨量對其生活史、生理特征影響很大,由于荒漠植物幼苗的早期階段最脆弱,對水土條件變化最敏感,所以紅砂種子萌發特性對環境的響應直接影響其幼苗存活及種群的更新動態。有研究預測,未來我國西北干旱地區降水量將有不同程度增加、降雨變幅震蕩將更為劇烈、極端降雨事件發生概率將大大增加[3]。目前就紅砂種子萌發對降雨格局變化的研究還很有限,且以往的研究大多數是在實驗室采用PEG6000 模擬干旱脅迫進行種子萌發試驗[27-28],而對降雨格局變化的效應研究相對較少。本研究提出以下科學問題: (1) 未來我國西北干旱區降雨格局變化對紅砂的種子萌發的影響如何? (2)降雨格局變化對紅砂從種子開始的幼苗更新是促進還是抑制作用?剖析降雨格局變化紅砂種子萌發的影響,對揭示紅砂對環境變化的響應和適應規律至關重要,也為預測未來我國西北干旱區降雨格局變化下,我國西北干旱區優勢荒漠植物紅砂的演化方向與速率提供理論支撐和數據支持。
1 材料與方法
1.1 實驗材料
2013年10月于中國生態系統研究網絡臨澤內陸河流域研究站附近收集新成熟的紅砂種子,將其帶回實驗室進行處理,冷藏于超低溫冰箱儲存過冬。2014年5月中旬將種子取出后,挑選健康、飽滿、大小均一的種子進行實驗。實驗用土均取自生態站附近紅砂原始生境表層土,過1 cm篩去除雜質后,在100 ℃條件下烘干48 h,殺死其中可能存在的雜草種子以及害蟲幼蟲。實驗用塑料圓柱形容器上底外側直徑10 cm,下底外側直徑約7 cm, 高20 cm(外徑),底部有排水孔,并在排水孔下套塑料袋,以防止水分溢出滴落到其他容器中。
1.2 實驗設計
根據中國科學院寒區旱區環境與工程研究所臨澤內陸河流域研究站多年(1967—2008)的氣象資料統計,該區多年平均降雨量為117.1 mm多年,多集中于7—9月(7月31.7mm、8月26.1mm、9月31.7mm)約占全年65%。降雨量較高的年份多為160 mm左右,比多年平均水平高出約30%,降雨量最低為82.9 mm,比多年平均水平低30%,因而以多年月平均降雨量為對照,在此基礎上增減水30%,分別用W、W-、W+表示。劉冰等[29]研究發現該區小于10d的降雨間隔期所占比率最大,為67.56%,且頻率基本穩定,而大于10 d的降雨間隔期頻率明顯下降且變異較大,未來我國西北干旱半干旱區的降雨間隔期將延長,因此,本試驗以5d為降雨間隔期來模擬自然降雨頻率(T1),適當延長間隔期至10d來模擬由降雨間隔時間延長導致的降雨量增加的大降雨事件(T2)。基于花盆開口的面積確定各處理平均每次模擬降雨量,通過澆水間隔時間和平均每次澆水量的控制實現對月總降雨量梯度和降雨間隔時間梯度的模擬,各處理模擬降水頻次降水量如表1。6個處理的代碼為:W-T1, 總降雨量在月多年月平均量基礎上減少30%,降雨間隔時間為5d(T1);W-T2,總降雨量在月多年月平均量基礎上減少30%,降雨間隔時間為10d(T2);WT1,總降雨量為多年月平均量,降雨間隔時間為5d(T1);WT2,總降雨量為多年月平均量,降雨間隔時間為10d(2);W+1,總降雨量在月多年月平均量基礎上增加30%,降雨間隔時間為5d(T1);W+T2,總降雨量在月多年月平均量基礎上增加30%,降雨間隔時間為10d(T2)。
6—9月這4個月是研究區域內紅砂幼苗種子萌發最為關鍵的時期,5月10日將360粒大小均一、健康的種子,播種于18個上述塑料花盆中,播種深度為0.5cm[30],每盆20粒,在人工氣候培養箱中進行培養,濕度為50%—70%,根據紅砂萌發所需的溫度設置(20±2)℃、 24 h無光照[20],每個處理3個重復。
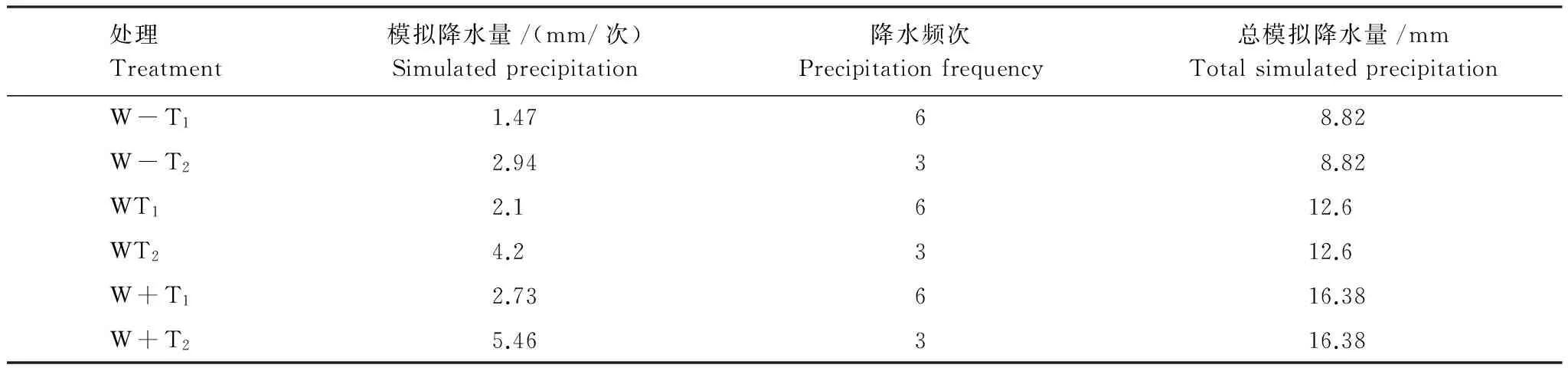
表1 實驗中的模擬降水設置
W,平均月降水量Monthly precipitation;W-,減水30% Water reduction by 30%;W+,加水30% Water addition by 30%;T1,降雨間隔時間為5d Precipitation interval is 5 days;T2,降雨間隔時間為10d precipitation interval is 10 days
1.3 萌發特征指數的測定及計算
播種1周后,即自5月17日起每天統計一次出土幼苗數并對其總數做詳細記錄,當出苗高峰過后連續觀測出土幼苗數均未發生變化時,結束實驗,發芽結束,統計種子出苗率,每個處理隨機挑選10粒發芽種子測量苗高。本實驗評價種子活力的參數包括出苗率(GR)、發芽勢(GP)、萌發指數(GI)和活力指數(VI),分別按下述公式計算有關萌發參數:GR=n×100%/N,式中n和N分別為萌發種子數和試驗過程中所用種子總數;GP=m×100%/N,式中m和N分別為發芽高峰期發芽的種子數和試驗過程中所用種子總數;GI=∑(Gt/Dt),式中Gt為在第t天的種子萌發個數,Dt為Gt對應的發芽天數[31],GI是反映種子萌發速率的指標,萌發指數越大,種子萌發速度就越快,在自然條件下便能快速出苗;VI=GI×S,S為幼苗平均總長度。萌發開始時間,即從播種后到第一粒種子萌發所需的天數。萌發高峰期,在種子萌發過程中日萌發率最高的時期。萌發持續時間,從開始萌發到萌發結束所需的天數。
1.4 數據處理
采用有重復觀測值的雙因素方差分析(two-way ANOVA)檢驗不同降雨量和降雨間隔時間水平下各參數的差異顯著性,當交互作用顯著時,采用單因素方差分析(one-way ANOVA)檢驗不同降雨量或降雨間隔時間處理對各參數的影響;當交互作用不顯著時,直接進行多重比較。最小顯著差數法(LSD)驗證相應數據的差異顯著性,檢驗的顯著性水平定為0.05。
2 結果
2.1 降雨格局變化對種子萌發出苗的影響
雙因素方差分析表明,總降雨量對種子出苗率和發芽勢均有顯著影響,但降雨間隔時間及兩者的交互作用對種子萌發并沒有產生顯著影響(P> 0.05)(表2)。從圖1可以看出,降雨間隔時間一致時,隨總降雨量的增加種子出苗率和發芽勢均呈增加的變化趨勢,單因子方法分析表明,降雨量增加30%顯著提高了種子萌發率(P﹤0.05),與自然總降雨量相比,其出苗率和發芽勢分別平均提高45.69%、39.86%;然而,降雨量減少30%使出苗率和發芽勢分別平均降低了12.86%、13.36%,但差異不顯著(P> 0.05)。從圖1還可以看出,相同降雨量條件下降雨間隔時間的延長,即總降雨量一致增大了單次降雨量(表1)均使紅砂種子出苗率和發芽勢分別平均提高了11.61%、11.65%,但差異不顯著(P> 0.05),說明總降雨量相同條件下降雨量間隔時間適當延長即單次降雨量增加能提高了種子的出苗率,也表明單次降雨事件增大有利于紅砂種子的萌發。
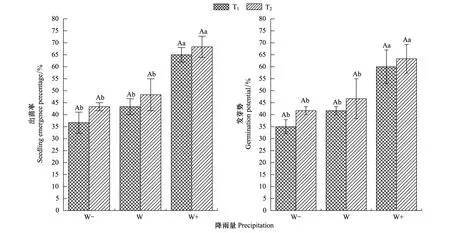
圖1 紅砂種子的出苗率和發芽勢(平均值±標準誤差)Fig.1 The germination rate and germination potential of R.soongorica seeds (mean ± SE)不同小寫字母表示在相同的降水間隔處理下,降水量增加或減少與對照間差異顯著(P<0.05);不同大寫字母表示相同降雨量下,降水間隔延長與對照間的差異顯著(P<0.05); T:降雨間隔時間為5d;T+:降雨間隔時間為10d;W:平均降雨量;W-:降雨量減少30%;W+,降雨量增加30%
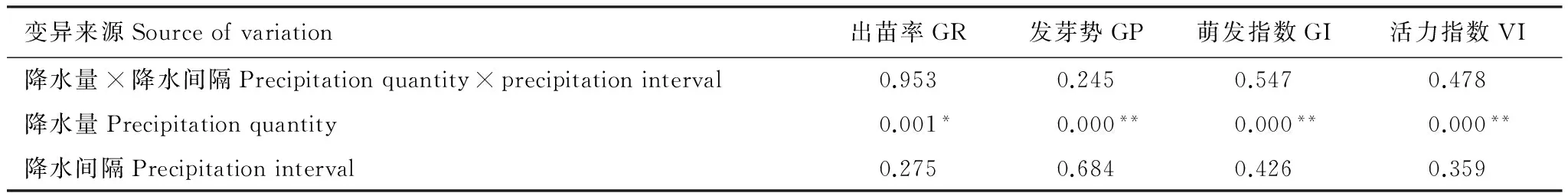
變異來源Sourceofvariation出苗率GR發芽勢GP萌發指數GI活力指數VI降水量×降水間隔Precipitationquantity×precipitationinterval0.9530.2450.5470.478降水量Precipitationquantity0.001*0.000**0.000**0.000**降水間隔Precipitationinterval0.2750.6840.4260.359
* 表示顯著水平(P< 0.05); **表示極顯著水平(P< 0.01);SRL:比根長 Specific Root Length;SRA:比表面積 Specific Root Area
2.2 降雨變化對種子萌發指數和活力指數的影響
雙因素方差分析表明,總降雨量對種子萌發指數有顯著影響,但降雨間隔時間及兩者的交互作用對種子萌發并沒有產生顯著影響(P> 0.05)(表2)。從表2可以看出,總降雨量增加30%顯著提高了種子萌發指數,降水間隔時間延長其效果更顯著,與自然總降雨量相比,其萌發指數平均提高57.67%。總降雨量減少30%雖降低了萌發指數,但與自然總降雨量相比差異不顯著(P> 0.05)。從表3還可以看出,同一總降雨量條件下,隨降水間隔時間的延長萌發指數均呈現出增加的變化趨勢。表明減少降雨次數增大單次降水量將更有利于提高荒漠植物種子的萌發速度。
種子活力指數可以體現種子是否生長和生長整齊度兩個因素,在一定降雨條件下,種子可以萌發但生長會受到抑制,導致種子活力指數降低。從表中可以看出,總降雨量減少30%,顯著降低了種子的活力指數,與自然總降雨量相比,其活力指數平均降低了25.92%。然而,在總降雨增加降雨間隔時間延長的情況下種子活力指數達到了最大值,顯著提高了種子的活力指數(P<0.05)。
2.3 降水變化對種子萌發進程的影響
經過20d的動態監測發現,紅砂種子開始萌發出苗主要集中在播種后第2周(第7天—第14天)(表2),萌發高峰期均出現在第10天左右(表2),第3周(第15天—第26天)基本趨于穩定(圖2)。在第2周,在降雨間隔時間下相同的情況下,總降雨量增加30%均提高了幼苗的出苗速率,降雨間隔時間延長尤為顯著,在出苗的第2天,其出苗速率就達到了38.33%,其種子出苗結束期也延遲了5d;總降雨量減少30%條件下,隨降水間隔時間的延長,種子出苗結束期也推遲了4d,表明延長降水間隔期增加大降水事件有利于提高種子的出苗速率。從表2還可以看出,在總降雨量增加降雨間隔時間延長的情況下,紅砂種子在第7天開始萌發,并在第8天就達到了萌發高峰期,表明荒漠區未來大降雨事件將有利于紅砂的幼苗自然更新。
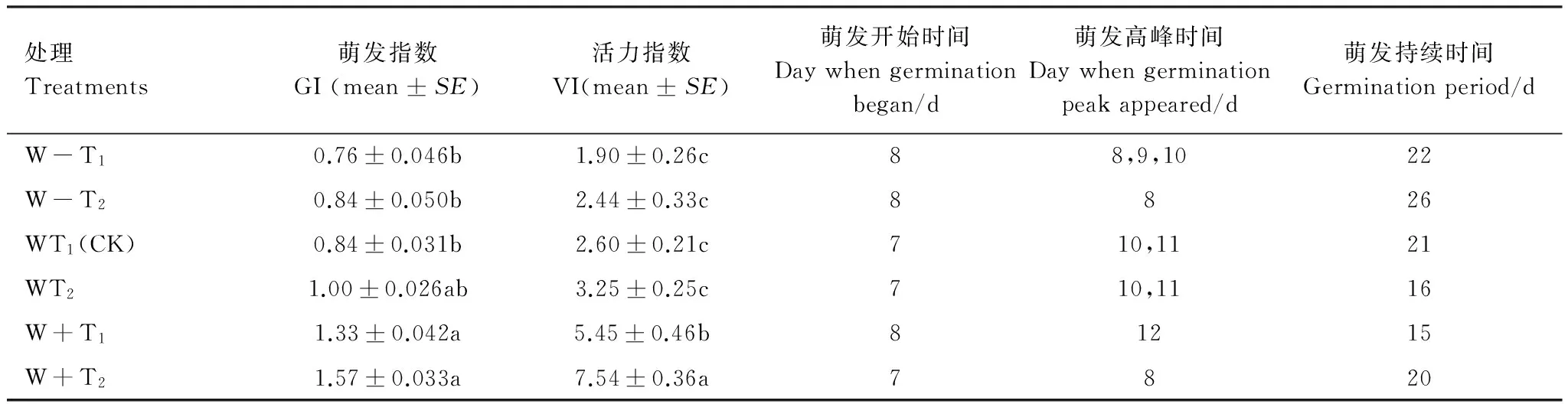
表3 不同降雨格局對種子萌發特性的影響
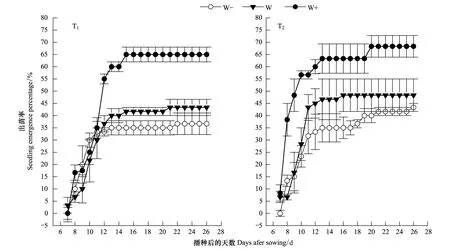
圖2 不同降水條件下播種1周后的出苗動態(平均值±標準誤差)Fig.2 Dynamic of seeding emergence after sowing in one week in a different precipitation (mean ± SE)
3 討論與結論
3.1 降雨格局變化對紅砂種子出苗的影響
在干旱和半干旱區,降雨是植物生長發育的主要限制因子,降雨量大小可以影響到該區荒漠植物的種子萌發,進而影響接下來幼苗的生長和存活能力[10]。本研究發現,隨總降雨量的增加紅砂種子萌發呈現出增加的變化趨勢,這與我國西北干旱半干旱區沙蓬(Agriophyllumsquarrosum(L.) Moq.)、油蒿(ArtemisiaordosicaKrasch.)、檸條錦雞兒(CaraganakorshinskiiKom.)等荒漠植物的研究結果一致[22],表明降雨量增加明顯改善了土壤的水分條件,提高了種子的萌發率。總降雨量減少30%條件下,降低了紅砂種子出苗率,說明總降雨量減少加劇了土壤的干旱化,繼而造成干旱脅迫,降低了種子出苗率,這與Hovenden等[16]和Gómez-Aparicio等[17]的研究結果一致。未來我國西北干旱區夏季降雨量將有不同程度增加[3],紅砂作為夏萌植物,這必將提高其種子的出苗率,增加其繁殖成功率。在干旱區種子萌發出苗不僅決定于降雨量,而且決定于降雨頻率,本研究發現,在總降雨量一致的情況下,適當延長降雨間隔時間(即減少了降雨頻率增大了單次降雨量)均提高了紅砂種子出苗率,總降雨量增加(30%)降雨間隔延長的處理其種子出苗率達到了最大值,表明降雨間隔時間增長而平均單次降雨量增多的降雨格局更有利于紅砂種子出苗率的提高,也說明紅砂種子萌發出苗主要受到降雨量的影響,但降雨量效應依賴降雨間隔時間,適當地增加降水量延長降水間隔時間會提高種子萌發出苗率,增加了繁殖成功率,對其幼苗更新起促進效應。這可能是因為紅砂種子萌發需水量高、種子萌發緩慢、種子萌發往往需要積水條件和較長的吸水時間[27,30],而降雨間隔時間增長而平均單次降雨量增多的降雨格局正好滿足了紅砂種子萌發出苗所需水量高的這種特性,進而提高了種子出苗率。有研究預測未來全球降雨將表現為降雨頻率減少、大降雨事件增加[32- 34],可見,未來降雨頻率減少單次降雨量增加的降雨事件將促進紅砂種子的萌發出苗。同時,本研究也發現:由于降水頻次的改變,在相同降雨量條件下延長降水間隔時間減少降雨頻率,其單次降雨量增加(表1),則出苗率均呈現出增加的趨勢,在單次降雨為5.46mm≈6mm時,其萌發率達到最大值68.33%,這與科爾沁沙地尖頭葉藜(ChenopodiumacuminatumWilld)在總降雨量不變降雨頻次由2d /1次增加到8d/1次,其種子出苗率達到最大值一致,表明紅砂種子萌發出苗主要決定于單次降雨量,單次降雨量是其種子出苗率高低的決定因素,也說明促使植被萌發存在最小降雨閾量[21]。然而,在不同的生態環境下,促使植被萌發的最小降雨閾量亦不同[21]。在科爾沁沙地促進尖頭葉藜萌發的最小降雨量是8mm[21];在莫哈韋沙漠和內蓋夫沙漠,促使一些沙漠物種萌發的最小降雨量范圍是10—15mm[24,35],而有研究發現在科威特當最少降雨量達到4mm時,足以促使車前屬物種的萌發[36]。本研究中當單次降雨量達到6mm時,才顯著促進了紅砂種子出苗,可見,在河西走廊荒漠區當降雨總量大于6mm,紅砂種子才能吸收足夠萌發所需的水分。同時,該試驗也驗證了筆者野外發現,在河西走廊降雨較大山前戈壁有大量的紅砂實生幼苗,而在荒漠綠洲過渡帶幾乎無紅砂實生幼苗,這可能是因山前戈壁降雨量大,且多為大于5mm大降雨事件,夏季單次較大的降雨量滿足紅砂種子的萌發的最少降雨量,既而出現了大量的種子萌發幼苗。因此,加強單次降雨量對荒漠植物種子萌發影響的研究,找到萌發閾值,對認識未來降雨格局變化荒漠植物自然更新過程顯得更有意義。
3.2 降雨格局變化對紅砂萌發進程的影響
降雨格局變化對紅砂的萌發進程產生了一定的影響。總降雨量增加降雨間隔延長所形成的大降雨事件顯著提高了紅砂的萌發指數(表3),萌發指數越大表示種子萌發速度越快。總降雨增大降雨間隔延長處理其萌發進程曲線較陡峭(從圖2),種子在第8天達到萌發高峰期,表明增大單次降雨縮短了種子的萌發持續時間,提早了種子的萌發高峰時間。在相同降雨間隔條件下,與對照相比,總降雨減少30%對種子的萌發進程影響不大。目前的研究認為,植物種子萌發表現出兩種不同的萌發機制:1)快速萌發和早期定居,從而能有效提高植物的競爭優勢[37-38];2)推遲萌發,是植物在不可預測環境中形成的另一種有效的風險分散策略[39]。紅砂為典型荒漠植物,在氣候惡劣、干旱少雨的荒漠地區,水分(特別是降雨)是其種子萌發的限制條件。本研究發現,在自然狀態(對照組)及降雨減少情況下紅砂種子出苗的變化趨勢則是先少量萌發,之后慢慢上升維持一個穩定的水平,這與科爾沁沙地尖頭葉藜在高頻率低降雨量條件下種子萌發出苗動態一致,表明這兩種荒漠植物在低降雨量條件下幼苗萌發采取的分批萌發類似K-對策[40],也說明目前干旱環境條件下紅砂采取的可能是第2種萌發策略,推遲種子萌發使其幼苗在更適宜的環境條件下出現和生長的機會增多,從而提高紅砂對環境的長期適應性,這說明低降雨量條件下的分批萌發對策是極端生境下植物長期適應自然選擇的結果[41]。一定的大降雨事件(即總降雨量增大降雨間隔時間延長的降雨格局即增大單次降雨量)提高了紅砂種子萌發指數,加快了種子的萌發進程,這也與科爾沁沙地尖頭葉藜通過延長降雨間隔增大單次降雨量,單次降雨量達到8mm時其幼苗大批萌發策略一致(類似r-對策)。這可能是因為大的降雨事件有利于打破種子的休眠,提高了紅砂種子萌發的整齊度。然而,有研究指出如果荒漠植物種子萌發強烈地響應于某一次降水,即有活力的種子同時全部萌發,干旱干擾就可能導致全部個體死亡[42]。紅砂分布區屬典型的大陸干旱氣候,有研究指出全球變化導致我國西北干旱區的降雨量逐漸增加[43],特別是春夏季降雨增加[44],這必將提高紅砂種子萌發率,加快紅砂種子的萌發進程,但西北干旱地區氣候特征為大氣干旱,大氣干旱是西北荒漠區種子萌發后幼苗死亡的直接原因,最終真正能補充到種群種的個體數量非常有限,幼苗更新受限[45]。由此可見,西北干旱區降雨量增加雖提高了紅砂在空間和時間上的競爭優勢,但因大氣干旱在一定程度上也降低了其抗干擾的能力。
[1] Houghton J T, Ding Y, Griggs D J, Noguer M, Van Der Linden P J, Dai X, Maskell K, Johnson C A. Climate Change 2001: The Scientific Basis. Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK:Cambridge University Press, 2001.
[2] Meehl G A, Arblaster J M, Tebaldi C. Understanding future patterns of increased precipitation intensity in climate model simulations. Geophysical Research Letters, 2005, 32(18): L18719.
[3] IPCC. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK: Cambridge University Press, 2007.
[4] Noy-Meir I. Desert ecosystems: environment and producers. Annual Review of Ecology and Systematics, 1973, 4: 25- 51.
[5] Reynolds J F, Kemp P R, Ogle K, Fernández R J. Modifying the ‘pulse-reserve’ paradigm for deserts of North America: precipitation pulses, soil water, and plant responses. Oecologia, 2004, 141(2): 194- 210.
[6] Austin A T, Yahdjian L, Stark J M, Belnap J, Porporato A, Norton U, Ravetta D A, Schaeffer S M. Water pulses and biogeochemical cycles in arid and semiarid ecosystems. Oecologia, 2004, 141(2): 221- 235.
[7] Sponseller R A. Precipitation pulses and soil CO2flux in a Sonoran Desert ecosystem. Global Change Biology, 2007, 13(2): 426- 436.
[8] Vile D, Pervent M, Belluau M, Vasseur F, Bresson J, Muller B, Granier C, Simonneau T.Arabidopsisgrowth under prolonged high temperature and water deficit: Independent or interactive effects? Plant, Cell and Environment, 2012, 35(4): 702- 718.
[9] Walck J L, Hidayati S N, Dixon K W, Thompson K, Poschlod P. Climate change and plant regeneration from seed. Global Change Biology, 2011, 17(6): 2145- 2161.
[10] Gillespie I G, Loik M E. Pulse events in Great Basin Desert shrublands: physiological responses ofArtemisiatridentataandPurshiatridentataseedlings to increased summer precipitation. Journal of Arid Environments, 2004, 59(1): 41- 57.
[11] Zheng Y R, Xie Z X, Yu Y, Jiang L H, Shimizu H, Rimmington G. Effects of burial in sand and water supply regime on seedling emergence of six species. Annals of Botany, 2005, 95(7): 1237- 1245.
[12] González-Astorga J, Núez-Farfán J. Variable demography in relation to germination time in the annual plantTagetesmicranthaCav. (Asteraceae). Plant Ecology, 2000, 151(2): 253- 259.
[13] Ambrose L G, Wilson S D. Emergence of the introduced grassAgropyroncristatumand the native grassBoutelouagracilis in a mixed-grass prairie restoration. Restoration Ecology, 2003, 11(1): 110- 115.
[14] Weltzin J F, McPherson G R. Implications of precipitation redistribution for shifts in temperate savanna ecotones. Ecology, 2000, 81(7): 1902- 1913.
[15] Adams H D, Kolb T E. Tree growth response to drought and temperature in a mountain landscape in northern Arizona, USA. Journal of Biogeography, 2005, 32(9): 1629- 1640.
[16] Hovenden M J, Newton P C D, Wills K E, Janes J K, Williams A L, Schoor J K V, Nolan M J. Influence of warming on soil water potential controls seedling mortality in perennial but not annual species in a temperate grassland. New Phytologist, 2008, 180(1): 143- 152.
[17] Gómez-Aparicio L, Pérez-Ramos I M, Mendoza I, Matías L, Quero J L, Castro J, Zamora R, Maraón T. Oak seedling survival and growth along resource gradients in Mediterranean forests: implications for regeneration in current and future environmental scenarios. Oikos, 2008, 117(11): 1683- 1699.
[18] 董麗佳, 桑衛國. 模擬增溫和降水變化對北京東靈山遼東櫟種子出苗和幼苗生長的影響. 植物生態學報, 2012, 36(8): 819- 830.
[19] 趙娟, 宋媛, 孫濤, 毛子軍, 劉傳照, 劉林磬, 劉瑞鵬, 侯玲玲, 李興歡. 紅松和蒙古櫟種子萌發及幼苗生長對升溫與降水綜合作用的響應. 生態學報, 2012, 32(24): 7791- 7800.
[20] Swemmer A M, Knapp A K, Snyman H A. Intra-seasonal precipitation patterns and above-ground productivity in three perennial grasslands. Journal of Ecology, 2007, 95(4): 780- 788.
[21] 馬赟花, 張銅會, 劉新平, 毛偉, 岳祥飛. 春季小降雨事件對科爾沁沙地尖頭葉藜萌發的影響. 生態學報, 2015, 35(12): 4063- 4070.
[22] Zhu Y J, Yang X J, Baskin C C, Baskin J M, Dong M, Huang Z Y. Effects of amount and frequency of precipitation and sand burial on seed germination, seedling emergence and survival of the dune grass Leymus secalinus in semiarid China. Plant and Soil, 2014, 374(1): 399- 409.
[23] 張勇, 薛林貴, 高天鵬, 晉玲, 安黎哲. 荒漠植物種子萌發研究進展. 中國沙漠, 2005, 25(1): 106- 112.
[24] Loria M, Noy-Meir I. Dynamics of some annual populations in a desert loess plain. Israel Journal of Botany, 1979, 28(3/4): 211- 225.
[25] Juhren M, Went F W, Phillips E. Ecology of desert plant. IV. Combined field and laboratory work on germination of annuals in the Joshua Tree National Monument, Califolia. Ecology, 1956, 37(2): 318- 330.
[26] Gutterman Y. Seed Germination in Desert Plants. Berlin Heidelberg: Springer-Verlag, 1993.
[27] 曾彥軍, 王彥榮, 薩仁, 田雪梅. 幾種旱生灌木種子萌發對干旱脅迫的響應. 應用生態學報, 2002, 13(8): 953- 956.
[28] 宋雪梅, 楊九艷, 呂美婷, 楊明, 張作如. 紅砂種子萌發對鹽脅迫及適度干旱的響應. 中國沙漠, 2012, 32(6): 1674- 1680.
[29] 劉冰, 常學向, 李守波. 黑河流域荒漠區降水格局及其脈動特征. 生態學報, 2010, 30(19): 5194- 5199.
[30] 曾彥軍, 王彥榮, 莊光輝, 楊鬃山. 紅砂和霸王種子萌發對干旱與播深條件的響應. 生態學報, 2004, 24(8): 1629- 1634.
[31] 高海娟, 云錦鳳, 劉德福. 荒漠草原地區3種冰草種子萌發的研究. 草業科學, 2007, 24(5): 64- 68.
[32] Easterling D R, Meehl G A, Parmesan C, Changnon S A, Karl T R, Mearns L O. Climate extremes: observations, modeling, and impacts. Science, 2000, 289(5487): 2068-2074.
[33] Weltzin J F, Loik M E, Schwinning S, Williams D G, Fay P A, Haddad B M, Harte J, Huxman T E, Knapp A K, Lin G H, Pockman W T, Shaw M R, Small E E, Smith M D, Samith S D, Tissue D T, Zak J C. Assessing the response of terrestrial ecosystems to potential changes in precipitation. BioScience, 2003, 53(10): 941- 952.
[34] Allan R P, Soden B J. Atmospheric warming and the amplification of precipitation extremes. Science, 2008, 321(5895): 1481- 1484.
[35] Beatley J C. Phenological events and their environmental triggers in Mojave desert ecosystems. Ecology, 1974, 55(4): 856- 863.
[36] Brown G. Community composition and population dynamics in response to artificial rainfall in an undisturbed desert annual community in Kuwait. Basic and Applied Ecology, 2002, 3(2): 145- 156.
[37] Grime J P, Mason G, Curtis A V, Rodman J, Band S R. A comparative study of germination characteristics in a local flora. Journal of Ecology, 1981, 69(3): 1017- 1059.
[38] Mandák B. Germination requirements of invasive and non-invasive Atriplex species: a comparative study. Flora-Morphology, Distribution, Functional Ecology of Plants, 2003, 198(1): 45- 54.
[39] Venable D L, Brown J S. The selective interactions of dispersal, dormancy, and seed size as adaptations for reducing risk in variable environments. The American Naturalist, 1988, 131(3): 360- 384.
[40] Andrews J H, Harris R F.r- andK-Selection and microbial ecology // Marshall K C, ed. Advances in Microbial Ecology. New York: Springer, 1986, 9: 99- 147.
[41] 趙惠勛. 群體生態學. 哈爾濱: 東北林業大學出版社, 1990: 40- 52.
[42] 王宗靈, 徐雨清, 王剛. 沙區有限降水制約下一年生植物種子萌發與生存對策研究. 蘭州大學學報: 自然科學版, 1998, 34(2): 98- 103.
[43] 施雅風, 沈永平, 李棟梁, 張國威, 丁永健, 胡汝驥, 康爾泗. 中國西北氣候由暖干向暖濕轉型的特征和趨勢探討. 第四紀研究, 2003, 23(2): 152- 164.
[44] 徐利崗, 周宏飛, 梁川, 吳安琪. 中國北方荒漠區降水多時間尺度變異性研究. 水利學報, 2009, 40(8): 1002- 1011.
[45] 田媛, 李建貴, 趙巖. 梭梭幼苗死亡與土壤和大氣干旱的關系研究. 中國沙漠, 2010, 30(4): 878- 884.
EffectsofsimulatedprecipitationchangeonseedgerminationofReaumuriasoongorica
SHAN Lishan, LI Yi*, ZHANG Zhengzhong, BAI Lei, DUAN Guifang, CHONG Peifang
CollegeofForestry,GansuAgriculturalUniversity,Lanzhou730070,China
國家自然科學基金(41361100,31560135,31360205,41461044);甘肅省科技支撐項目(1604FKCA088);甘肅省高等學校科研項目(2015A-067);甘肅農業大學青年研究生指導教師扶持基金(GAU-QNDS-201605)
2016- 05- 27; < class="emphasis_bold">網絡出版日期
日期:2017- 03- 26
*通訊作者Corresponding author.E-mail: liyi@gsau.edu.cn
10.5846/stxb201605271024
單立山,李毅,張正中,白蕾,段桂芳,種培芳.人工模擬降雨格局變化對紅砂種子萌發的影響.生態學報,2017,37(16):5382- 5390.
Shan L S, Li Y, Zhang Z Z, Bai L, Duan G F, Chong P F.Effects of simulated precipitation change on seed germination ofReaumuriasoongorica.Acta Ecologica Sinica,2017,37(16):5382- 5390.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
