BKR基因啟動子鑒定及表達調控
2017-10-18 02:59:44潘琳褚春旭李梓文孫基豐張昊
長春理工大學學報(自然科學版) 2017年4期
關鍵詞:檢測
潘琳,褚春旭,李梓文,孫基豐,張昊
(長春理工大學 生命科學技術學院,長春 130022)
BKR基因啟動子鑒定及表達調控
潘琳,褚春旭,李梓文,孫基豐,張昊
(長春理工大學 生命科學技術學院,長春 130022)
BKR基因是睪丸酮叢毛單胞菌中的一個重要的甾體化合物降解酶基因,TetR、LysR和LuxR蛋白是睪丸酮叢毛單胞菌中的調控蛋白。以EGFP為報告基因,構建pK-BKR-EGFP質粒,通過與pK-EGFP分別轉化到大腸桿菌JM109,檢測熒光信號,確定預測的序列中包含BKR基因的啟動子。然后通過質粒共轉化技術,檢測TetR、LysR和LuxR蛋白與pK-BKR-EGFP的熒光信號強度,探討TetR、LysR和LuxR三個蛋白與BKR基因之間的調控關系。實驗結果表明:BKR基因的啟動子在該基因前699bp內;LysR與BKR基因共轉后熒光信號增強;TetR和LuxR分別與BKR基因共轉后熒光信號減弱。由此可以得出結論,LysR對BKR基因的調控起增強作用;TetR和LuxR對BKR基因的調控起抑制作用。該實驗揭示了睪丸酮叢毛單胞菌中BKR基因的調控序列,為CT菌利用甾體激素作為唯一的碳源及能源機理提供理論基礎。
啟動子;增強綠色熒光蛋白;BKR基因
甾體類化合物對生態環境的污染日趨嚴峻,引發人和動物的一系列不良的生理反應[1,2]。睪丸酮叢毛單胞菌ATCC11996(Comamonas testosteroni ATCC11996)具有降解多種甾體激素的能力[3],在降解環境中甾體激素污染上具有很高的應用價值[4],使人類能夠避免一些環境中類固醇激素對人健康的危害。對睪丸酮叢毛單胞菌早期研究發現幾個類固醇代謝酶[5,6,7],對類固醇代謝起關鍵作用[8]。
BKR基因通過DNA序列分析表明,屬于短鏈脫氫酶(SDR)家族,并參與脂肪酸的合成。先前的研究表明,BKR基因是睪丸酮叢毛單胞菌降解激素所必須的基因[9]。TetR[10]、LysR[11,12]和LuxR[13]基因是菌體中的一種調控基因,可能對BKR基因具有調控作用,但目前尚未有明確的認識。為深入了解TetR、LuxR和LysR基因對BKR基因的調控作用,該實驗構建了以增強綠色熒光蛋白(Enforcement green fluorescence protein,EGFP)[14]為報告基因的啟動子鑒定質粒,并在此基礎上明確了TetR、LuxR和LysR基因對BKR基因的調控作用。以期了解BKR基因的調控基因,進一步了解睪丸酮叢毛單胞菌的調控模型。
1 材料與方法
1.1 材料
質粒pK-EGFP及睪丸酮叢毛單胞菌由德國基爾大學熊光明教授贈送。質粒Puc19、Puc-TetR、pET 15b、pET-LuxR及pET-LysR為長春理工大學實驗室所保存。pUC 18-T,連接酶及PCR反應所以試劑均為Takara公司產品。限制性內切酶 Sal I,BamH I及 buffer為 Thermo公司產品。質粒小量抽提試劑盒及膠回收試劑盒為上海生工產品。JM109菌株為promega公司產品。其他常用試劑為實驗室自行配制。
1.2 方法
1.2.1 帶EGFP報告基因的啟動子鑒定質粒pKBKR-EGFP的構建
如圖1所示,通過PCR及TA克隆構建BKRT質粒,用限制性內切酶Sal I和BamH I對pKEGFP和BKR-T進行雙酶切,分別回收大片段和小片段,以T4DNA連接酶連接后獲得質粒pKBKR-EGFP。
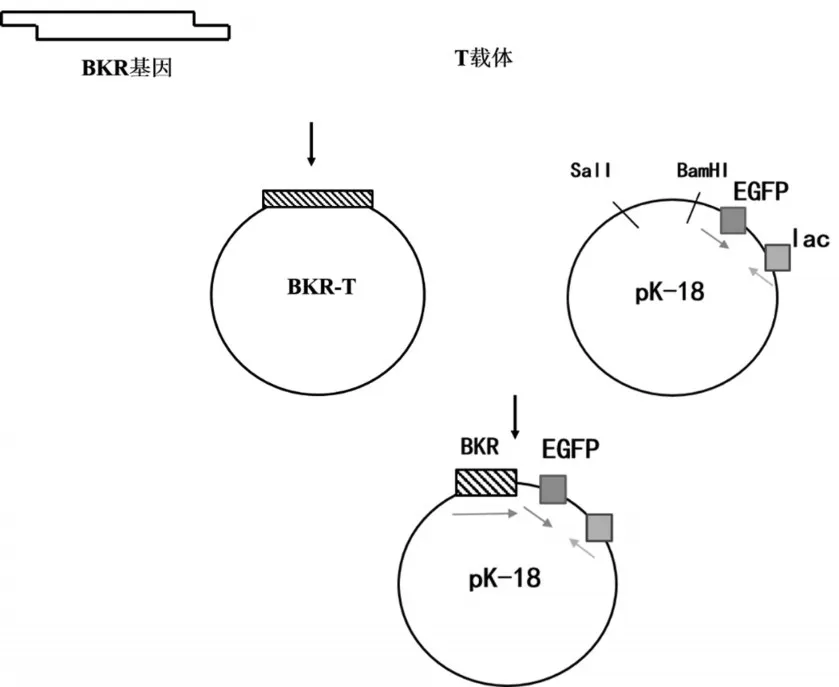
圖1 質粒Pk-BKR-EGFP構建路線圖
1.2.2 BKR-T質粒的構建
1.2.2.1 引物設計與合成
根據睪丸酮叢毛單胞菌基因組序列,設計合成一對擴增引物。上游引物為5’-GTCGACCAGCGACCAGCTGCTGCAAAAG-3’,含有Sal I酶切位點;下游引物為5’-GGATCCTGCTGTCTCCTTGGGTGCG-3’,含有 BamH I酶切位點。擴增產物為BKR-699bp片段。引物有吉林省庫美生物科技有限公司合成。
1.2.2.2 PCR擴增
反應體系:25μl,2xTaq Maste Mix,上下游引物(10pmol/μl)各 1μl,睪丸酮叢毛單胞菌模版 1μl,ddH2O補足至25μl。反應條件為:94℃預變性10min;94℃變性45s,58℃退火45s,72℃延伸50s,循環35次后72℃延伸10min。擴增產物用1%瓊脂糖凝膠電泳,觀察并成像。
1.2.2.3 質粒構建
PCR產物經膠回收后與pUC 18-T進行連接。連接產物轉化到感受態JM109細菌內,接種至LB平板,37℃過夜培養。
1.2.2.4 克隆鑒定
挑取LB平板上生長的單個菌落至LB液體培養基,37℃,180rpm/min震搖培養16h,取1.5ml細菌用質粒小量抽提試劑盒提取質粒,并進行序列分析。DNA序列分析由吉林省庫美生物科技有限公司完成。
1.2.3 pK-BKR-EGFP基因的構建
使用Sal I和BamH I將BKR-T和pK-EGFP進行雙酶切,用T4DNA連接酶將回收后的片段進行連接,過夜后轉化到感受態JM109中并進行克隆鑒定。
1.2.4 BKR基因啟動子的熒光檢測
將pK-EGFP和pK-BKR-EGFP復蘇后,分別取1ml菌液進行離心,13000rpm,1min后,使用生理鹽水洗滌2次,13000rpm,1min,用1ml生理鹽水將菌體懸起。通過紫外分光光度計在OD600處將pK-EGFP和pK-BKR-EGFP調至相同OD值。后用酶標儀在485nm激發光和530nm發射光處進行熒光檢測。
1.2.5 質粒共轉化
通過質粒共轉化技術將pK-BKR-EGFP質粒 分別與 pUC 19、pUC-TetR、pUC-LuxR 及pUC-LysR質粒進行共轉化。其中pUC 19為對照。
1.2.6 熒光檢測
將共轉化的質粒進行復蘇,分別去1ml菌液分別取1ml菌液進行離心,13000rpm,1min后,使用生理鹽水洗滌2次,13000rpm,1min,用1ml生理鹽水將菌體懸起。通過紫外分光光度計在OD600處將共轉化的質粒調至相同OD值。后用酶標儀在485nm激發光和530nm發射光處進行熒光檢測。
2 結果與討論
2.1 BKR-T質粒的構建
2.1.1 BKR基因啟動子片段PCR
根據引物的設計,PCR應擴增出699bp的產物,為完整的BKR基因的5’非翻譯區。如圖2所示,PCR產物位置與設計一致。
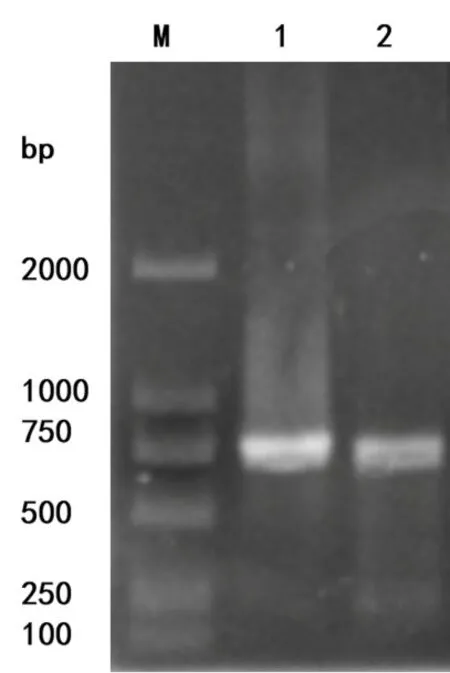
圖2 BKR基因啟動子PCR
孔道M:DNA的標準Marker;孔道1,2:BKR基因PCR的目的片斷
2.1.2 BKR-T陽性克隆鑒定
挑取5個克隆做質粒初篩實驗,如圖3所示,以T載體自連為對照可以看出質粒1,2的條帶大小與目的質粒大小相同。送去吉林省庫美生物科技有限公司測序出的基因序列,與目的基因序列一致。結果表明已經成功構建BKR-T質粒。

圖3 1%瓊脂糖凝膠電泳鑒定BKR-T質粒
孔道T:T載體自連;孔道1,2,3,4,5,6:BKRT目的片斷;孔道M1:DNA的標準Marker
2.2 pK-BKR-EGFP質粒的構建
將構建成功的BKR-T質粒通過限制性內切酶Sal I和BamH I將BKR片段酶切下來與通過限制性內切酶Sal I和BamH I酶切的質粒pK-EGFP,連接成功,轉化后獲得多個陽性克隆,其結果如圖4所示,送去吉林省庫美生物科技有限公司測序出的基因序列,與目的基因序列一致。結果表明已經成功構建pK-BKR-EGFP。

圖4 1%瓊脂糖凝膠電泳鑒定BKR-EGFP質粒
孔道 M1:DNA 的標準 Marker;孔道 pKEGFP:pK-EGFP 質粒;孔道 1,2,3,4,5:BKREGFP目的片斷
2.3 BKR基因啟動子的熒光檢測
將攜帶有重組質粒pK-BKR-EGFP和pKEGFP的JM109菌株分別涂布于LB固體培養基,37℃,過夜培養。通過紫外分光光度計在OD600處將共轉化的質粒調至相同OD值。后用酶標儀在485nm和530nm處進行熒光檢測。檢測結果如圖5所示。由圖可以看出含有BKR基因啟動子的BKR-EGFP的熒光信號是對照組pK-EGFP的二倍,由此可以得出該5’非翻譯區為BKR基因的啟動子區域。
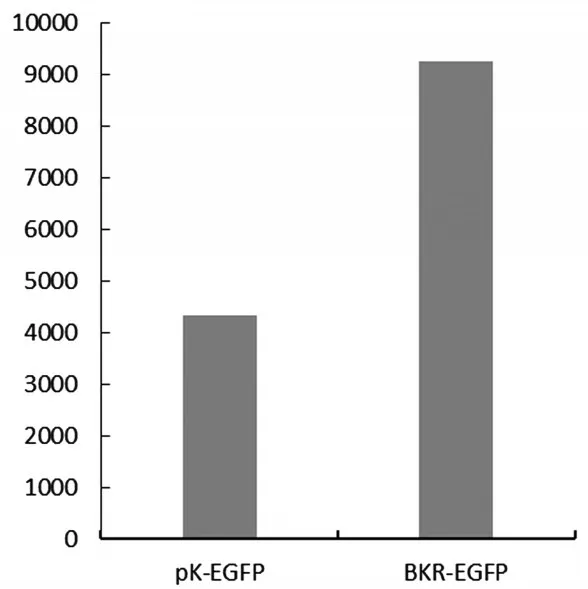
圖5 BKR基因啟動子熒光檢測
2.4 TetR、LuxR和LysR對BKR基因調控作用的熒光檢測
以pUC 19為對照,分別將pUC 19-TetR,pUC 19-LysR,pUC 19-LuxR與BKR-EGFP進行共轉化。將共轉化的質粒通過紫外分光光度計在OD600處將共轉化的質粒調至相同OD值。后用酶標儀在485nm和530nm處進行熒光檢測。檢測結果如圖6所示,可以看出TetR和LuxR的熒光信號低于對照組,而LysR的熒光信號高于對照組,故得出結論LysR對BKR基因的調控起增強作用;TetR和LuxR對BKR基因的調控起抑制作用。
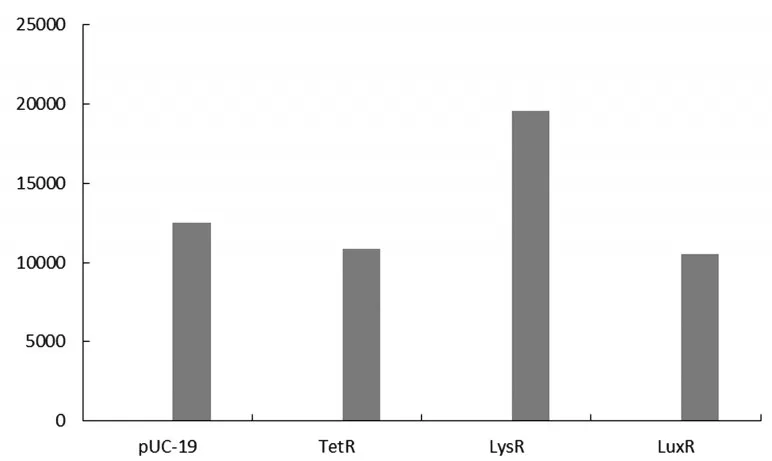
圖6 TetR、LysR、LuxR和BKR基因共轉的熒光檢測
3 結論
實驗通過熱擊法將重組載體轉入至JM109中,成功構建了pK-BKR-EGFP工程菌。通過對目的基因進行熒光檢測,檢測結果表明該非翻譯區確定為啟動子區域。在此基礎上以pUC 19為對照進行共轉化,得到含BKR-pUC 19-EGFP、BKR-pUC19-TetR-EGFP、pK-pUC19-LysREGFP、BKR-pUC 19-LuxR-EGFP 共轉 化 菌株。進行熒光檢測得到以下結論:LysR對BKR基因的調控起增強作用;TetR和LuxR對BKR基因的調控起抑制作用。
實驗通過質粒共轉化,熒光檢測等生物技術手段,對BKR基因的調控機制進行研究,揭示睪丸酮叢毛單胞菌中BKR基因的調控序列,為睪丸酮叢毛單胞菌利用甾體激素作為唯一的碳源及能源機理提供理論基礎,為了進一步分析睪丸酮叢毛單胞菌的調控模型,后續將進行ELISA和EMSA實驗對上述結論進行驗證,為開發環境中痕量甾體激素的檢測試劑提供基礎。
[1]Colborn T,vom Saal F S,Soto A M.Developmental effects of endocrine disrupting chemicals in wildlife and humans[J].Environ.Health Perspect,1993(101):378-384.
[2]Khan S,Cao Q,Lin A J,et al.Concentrations and bioaccessibility of polycyclic aromatic hydrocarbons in wastewater-irrigated soil using in vitro gastrointestinal test[J].Environmental Science and Pollution Research,2008,15(4):344-353.
[3]Garcíavaldés E,Cozar E,Rotger R,et al.New naphthalene-degrading marine Pseudomonas strains[J].Applied&Environmental Microbiology,1988,54(10):2478–2485.
[4]Coulter A W,Talalay P.Studies on the microbiological degradation of steroid ring A[J].Journal of Biological Chemistry,1968,243(12):3238-3247.
[5]戴藝民.睪丸酮叢毛單胞菌3α-HSD/CR等基因的表達調控研究[D].福州:福建農林大學,2005.
[6]Wu Y,Huang P,Xiong G,et al.Identification and isolation of a regulator protein for 3,17β-HSD expressional regulation in Comamonas testosteroni[J].Chemico-Biological Interactions,2015,234(9):197-204.
[7]Maser E,M?bus E,Xiong G.Functional expression,purification,and characterization of 3alpha-hydroxysteroid dehydrogenase/carbonyl reductase from Comamonas testosteroni[J].Biochemical&Biophysical Research Communications,2000,272(2):622-628.
[8]Wu Y,Huang P,Xiong G,et al.Identification and isolation of a regulator protein for 3,17β-HSD expressional regulation in Comamonas testosteroni[J].Chemico-biological interactions,2015,234(9):197-204.
[9]Zhang H,Ye J,Wang Y,et al.Cloning and characterization of a novel β-ketoacyl-ACP reductase from Comamonas testosteroni[J].Chem Biol Interact.2015,234(9):213-220.
[10]Pan T,Huang P,Xiong G,et al.Isolation and identification of a repressor TetR for 3,17β-HSD expressional regulation in Comamonas testosteroni[J].Chemico-biological interactions,2015(234):205-212.
[11]Gong W,Xiong G,Maser E.Oligomerization and negative autoregulation of the LysR-type transcriptional regulator HsdR from Comamonas testosteroni[J].Journal of Steroid Biochemistry&Molecular Biology,2012,132(3-5):203-211.
[12]李明堂,王清爽,于源華.睪丸酮叢毛單胞菌LysR基因的克隆及對3α-HSD/CR表達的調節[J].吉林農業大學學報,2011,33(6):624-627.
[13]Pruneda-Paz J L,Linares M,Cabrera J E,et al.TeiR,a LuxR-type transcription factor required for testosterone degradation in Comamonas testosteroni[J].Journal of Bacteriology,2004,186(5):1430-1437.
[14]Xiong G,Maser E.Construction of a biosensor mutant of Comamonas testosteroni for testosterone determination by cloning the EGFP gene downstream to the regulatory region of the 3,17β-HSD gene[J].Chemico-Biological Interactions,2014(234):188-196.
Identification and Expression Regulation of BKR Gene Promoter
PAN Lin,CHU Chunxu,LI Ziwen,SUN Jifeng,ZHANG Hao
(School of Life Science and Technology,Changchun University of Science and Technology,Changchun 130022)
BKR gene is an important steroid degrading enzyme gene in Comamonas testosteroni.TetR,LysR and LuxR proteins are the regulatory proteins in Comamonas testosteroni.In this experiment,EGFP was used as reporter gene and pK-BKREGFP plasmid was constructed.By converting the pK-EGFP to E.coli JM109,we detected fluorescence signals and determined that our predicted sequence contained the promoter of the BKR gene.Then,the signal intensity of TetR,LysR and LuxR protein and pK-BKR-EGFP was detected by plasmid CO transformation technique,and the relationship between the three proteins of etR,LysR and LuxR and BKR gene was investigated.The results showed that the promoter of BKR gene was enhanced in the pre gene 699bp,and the fluorescence signal was enhanced when LysR and BKR genes were co transferred;the fluorescence signals of TetR and LuxR were decreased when they were co transferred with BKR gene.From this we can conclude that LysR plays an important role in the regulation of BKR gene;TetR and LuxR inhibit the regulation of BKR gene.This experiment has revealed the regulation sequence of BKR gene in the testis,and provided the theoretical basis for the use of steroid hormones as the sole carbon source and energy mechanism for Comamonas testosteroni.
promoter;EGFP;BKR gene
Q789
A
1672-9870(2017)04-0129-04
2017-06-22
潘琳(1996-),女,本科,E-mail:1009670449@qq.com
張昊(1985-),女,博士,講師,E-mail:853287300@qq.com
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
