α7神經型尼古丁受體激動劑PNU282987對APP/PS1小鼠海馬組織中AP180蛋白表達的影響
2017-11-02 03:00:16鄧于新王曉玲吳昌學官志忠齊曉嵐
中風與神經疾病雜志 2017年10期
鄧于新, 王曉玲, 吳昌學, 官志忠,3, 齊曉嵐
α7神經型尼古丁受體激動劑PNU282987對APP/PS1小鼠海馬組織中AP180蛋白表達的影響
鄧于新1,2, 王曉玲1,2, 吳昌學1,2, 官志忠1,2,3, 齊曉嵐1,2
目的探討特異性激動APP/PS1轉基因小鼠中的α7nAChR后其海馬組織中突觸后膜蛋白的表達變化。方法選擇16只經過鑒定的APP/PS1小鼠隨機分為APP/PS1組(APP/PS1)和APP/PS1+PNU282987組(AP),每組各8只;另選16只野生型小鼠隨機分為對照組(Control)和野生+PNU282987組(WP),每組各8只。飼養小鼠達到24周齡后,AP組和WP組于每天進行Morris水迷宮行為學測試前4 h腹腔注射PNU282987 (1 mg/kg/d),之后利用Morris水迷宮進行行為學測試,Real-time PCR及Western blotting法分別檢測各組小鼠海馬組織中AP180 mRNA及蛋白水平的表達情況。結果與對照組比較,APP/PS1組海馬組織中AP180 mRNA及蛋白表達水平明顯降低(P<0.01,P<0.01);而特異性激動α7 nAChR水平后,WP組中AP180 mRNA和蛋白水平升高(P<0.01,P<0.05);與APP/PS1組相比AP組小鼠大腦海馬組織中AP180 mRNA和蛋白水平明顯升高(P<0.01,P<0.01)。結論特異性激動APP/PS1轉基因小鼠海馬組織中α7 nAChR能夠使網格蛋白介導的突觸囊泡內吞過程中的關鍵蛋白AP180表達水平升高。這可能提示了α7nAChR對突觸有一定的保護作用,進一步說明α7nAChR在阿爾茨海默病的發病中起著重要作用。
α7神經型尼古丁受體; 阿爾茨海默病; APP/PS1轉基因鼠; AP180
阿爾茲海默病(Alzheimer’s disease,AD)是一種以進行性認知功能障礙和記憶損傷為主要臨床特征的致死性神經退行性疾病,最早定義為大腦神經元和突觸的損傷[1]。AD的核心病理因素β淀粉樣蛋白(Aβ)的聚集直接或間接的導致了突觸結構和功能的破壞[2],也有研究表明,突觸數量的減少與認知能力的減退有密切的聯系[3],從一定程度上表明突觸的結構損壞和功能障礙在AD認知功能下降與年齡相關的神經元結構和功能改變中扮演了重要角色[4]。α7膽堿能尼古丁受體(α7nAchR)在AD的發病中具有神經保護性作用而成為AD研究中的重要對象[5],而AP180是網格蛋白介導的突觸囊泡內吞過程中的關鍵蛋白之一。本研究旨在以銜接蛋白180(AP180)為例,探討α7nAchR水平對小鼠海馬組織中突觸相關蛋白表達的影響,為AD的防治新思路提供依據。
1 材料與方法
1.1 實驗動物 雄性阿爾茨海默病APPswe/PSldE9雙轉基因模型小鼠(B6.Cg-Tg)及同背景雌性野生型小鼠,體重20~30 g,均購自上海南方模式生物公司[動物許可證號:SCXK(滬)2014-0002]。繁殖采用1只雄鼠與3只雌鼠同居的方式進行。成功繁殖后,當新生的小鼠年齡達到2~3 w時剪鼠尾提取小鼠DNA,進行小鼠基因型鑒定。第19~20天后子鼠雌雄分籠飼養。實驗動物遵照國家實驗動物飼養和使用指南,采用獨立通風柜飼養,動物房常年溫度控制在(24±2)℃,濕度40%~60%,光照時間8:00~21:30。所有動物均飼養于貴州醫科大學實驗動物中心SPF級環境。所有實驗操作均獲得貴州醫科大學實驗動物倫理委員會的許可。
1.2 試劑 Morris水迷宮、Morris水迷宮監測軟件購于中國醫學科學院藥物研究所;TANON 4200全自動化學發光成像分析系統購于上海天能科技有限公司;兔抗鼠α7 nAchR多克隆抗體(sc-5544)、辣根過氧化物酶標記的羊抗鼠二抗均購于美國SANTA CRUZ公司;兔抗鼠AP180多克隆抗體(ab33898)購于英國Abcam公司;辣根過氧化物酶標記的羊抗兔二抗購于美國cell signaling公司;PNU282987購于Sigma公司;BCA蛋白定量分析試劑盒購于美國Thermo公司;RIPA高效裂解液購于北京索萊寶公司;ECL發光試劑、PVDF膜均購于美國Millipoe公司;RNA逆轉錄試劑盒購于Thermo公司;Trizol試劑購于Invitrogen公司;β-actin、AP180定量PCR的引物均由上海生物工程有限公司設計并合成。
1.3 動物飼養繁殖及分組 APP/PS1雙轉基因雄鼠同相同背景的野生型雌鼠雜交進行繁殖,對新生小鼠剪尾提取DNA鑒定,APP/PS1轉基因小鼠飼養至24 w,野生型小鼠相同條件下飼養至24 w。將16只經過鑒定的APP/PS1小鼠隨機分為APP/PS1組和APP/PS1+PNU282987組(AP組),每組各8只;另將16只野生型小鼠隨機分為空白對照組(Control組)和PNU282987組(WP組),每組各8只。
1.4 方法
1.4.1 Morris水迷宮行為學檢測 小鼠飼養至24周齡后,各組進行Morris水迷宮測試,測試持續5 d,其中APP組和WP組于每天測試前腹腔注射α7 nAchR激動劑PNU282987,劑量為1 mg/kg。記錄Morris測試過程中小鼠的逃避潛伏期及穿越平臺次數。實驗內容包括:(1)定位航行實驗:共進行4 d,每只小鼠每天完成4次訓練,每次從不同象限將小鼠面向池壁放入水中,觀察并記錄其穿越平臺的時間、次數以及停留平臺的時間。如在60 s內未能尋找到平臺,則將其引導至平臺并停留10 s。(2)空間探索實驗:第5天測試,撤除平臺。記錄小鼠在60 s內各象限尋找平臺的次數和逗留平臺象限時間,找到平臺次數越多和逗留平臺象限時間長,說明空間記憶能力好(見圖1)。
1.4.2 組織處理及蛋白提取 各組小鼠在行為學測試結束后,腹腔注射4%水合氯醛(0.2 ml/10 g)進行麻醉, 開胸經左心室快速灌注預冷0.1 mol/L PBS 40 ml后,快速除去顱蓋,小心取出腦組織在冰上分離得到海馬組織,于-80 ℃保存備用。
1.4.3 Real-time PCR檢測AP180 mRNA表達水平 運用Trizol一步法提取細胞總RNA后將mRNA逆轉錄成為cDNA備用,應用ABI Step One Plus型實時熒光定量PCR儀以逆轉錄得到的cDNA作為模板進行Real-time PCR,所用引物由生工合成(引物序列見表1)。運用SDS 2.1軟件收集和分析AP180、β-actin基因擴增各循環產生的熒光信號,并計算上述基因在各實驗組與對照組比較的相對表達水平(RQ=2-ΔΔCt)。重復3次獨立實驗,各樣本每次3個復孔。
1.4.4 Western 印跡法檢測海馬組織蛋白中AP180蛋白水平 收集小鼠海馬組織,加入裂解液(RIPA∶PMSF=100∶1),12000 r、4 ℃離心20 min,收集上層清液即為蛋白質。用BCA蛋白定量試劑盒進行蛋白定量,Western印跡法檢測各組中AP180蛋白表達水平,以各組蛋白條帶像素灰度值與內參蛋白β-actin的比值表示各組蛋白的相對表達水平。重復3次獨立檢測,每次3個復孔。
2 結 果
2.1 24周齡APP/PS1雙轉基因小鼠的學習記憶能力測試 24周齡各組小鼠通過Morris水迷宮測試,在第5天的定位航行試驗中,以各組小鼠在目標象限持續時間(s)、平均逃避潛伏期(s)及跨越平臺次數(N)為認知能力指標進行比較,結果顯示,24周齡APP/PS1雙轉基因小鼠在目標象限持續時間縮短、逃避潛伏期明顯延長、穿越平臺次數少,與野生型小鼠比較均有統計學意義(P<0.01),表明24周齡APP/PS1雙轉基因小鼠確實存在學習記憶能力方面的損害;而且AP組與APP/PS1組比較,平臺所在象限持續時間延長,逃避潛伏期縮短,穿越平臺次數增多,差異均具有統計學意義(P<0.01),提示注射PNU282987激動α7 nAchR具有保護APP/PS1雙轉基因小鼠學習記憶能力的作用;另外,WP組與野生型小鼠比較,平臺所在象限持續時間延長,逃避潛伏期縮短,穿越平臺次數增多,差異具有統計學意義(P<0.01),也在一定程度上提示了激動α7 nAchR具有保護小鼠學習記憶能力的作用。
2.2 各組中AP180 mRNA及蛋白表達水平 Real-time PCR檢測AP180 mRNA表達水平,結果顯示:與對照組比較,WP組AP180 mRNA表達水平增高,APP/PS1組AP180 mRNA表達水平降低,差異具有統計學意義(P<0.01);AP組與APP/PS1組比較,AP180 mRNA表達水平增高,差異具有統計學意義(P<0.01)。運用Western blot檢測各組小鼠海馬AP180蛋白表達情況,結果顯示:與對照組相比較,WP組AP180蛋白表達水平均增高(P<0.05),APP/PS1組AP180蛋白表達水平降低,差異具有統計學意義(P<0.01);AP組與APP/PS1比較,AP180蛋白表達水平增高,差異具有統計學意義(P<0.01)(見圖2)
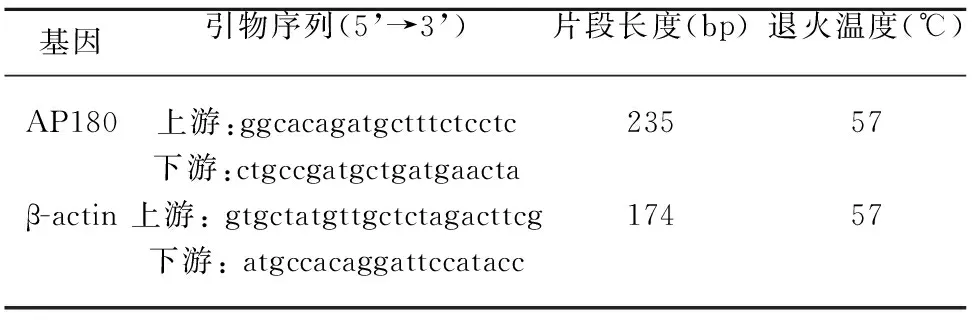
表1 熒光定量PCR引物序列及產物片段
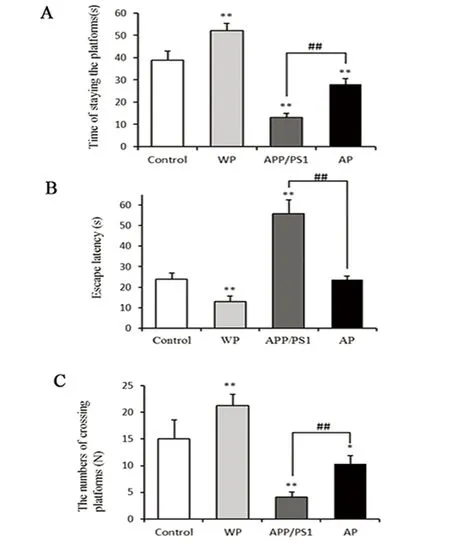
與對照組相比差異有統計學意義*P<0.05;與對照組相比差異有高度統計學意義**P<0.01;與APP/PS1組之間比較差異具有高度統計學意義##P<0.01
圖1 各組小鼠Morris水迷宮學習記憶能力測試結果
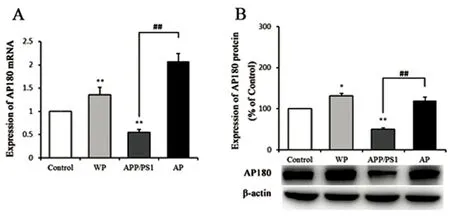
與對照組相比差異有統計學意義*P<0.05;與對照組相比差異有高度統計學意義**P<0.01;與APP/PS1模型組相比差異有高度統計學意義##P<0.01
圖2 各組小鼠海馬組織中AP180 mRNA(A)及蛋白表達水平(B)
3 討 論
阿爾茨海默病(AD)是一種越來越常見的神經退行性病變,多發于老年人,在65歲以上的人群中,AD的發病率已經達到約5%~10%[6]。AD的主要臨床特征為進行性的認知功能損害和記憶能力下降。在AD患者大腦存在幾個典型的病理特征:細胞外的淀粉樣斑塊沉積、細胞內的神經纖維纏結以及基底前腦膽堿能神經元變性和突觸丟失,其中突觸丟失與認知功能下降有更好的相關性[7]。APP/PS1雙轉基因小鼠可表達突變的人類早老素(DeltaE9)和人鼠淀粉樣前蛋白(APPswe)融合體。APP/PS1雙轉基因小鼠APP和Psl基因的突變可以導致Aβ水平的升高,模擬AD的神經病理過程[8]。膽堿能系統功能降低在AD早期階段已有表現,在AD的發生發展中有一定作用[9]。膽堿能系統中的神經型尼古丁受體(Neuronal nicotinic acetylcholine receptor,nAChR)在大腦的學習、認知及記憶功能方面有一定的調節作用,并參與許多信號傳遞的調控,具有顯著的神經保護作用;nAChR是第二信使非依賴性、門控性離子通道偶聯受體,可與神經遞質結合,本身也是離子通道,其開放不需要第二信使的參與。nAChR由α和β兩種亞基組成,不同種類的亞基以不同的組合形式組成不同的五聚體結構,主要調節細胞內外Na+、K+和Ca2+等離子的流動,其表達量的改變與Aβ呈負相關,在AD的發展中起關鍵的調控作用[10,11]。
α7nAChR 是神經型尼古丁受體中重要的成員[12],有報道指出在AD中最脆弱的神經元是那些高表達神經型尼古丁受體的神經元,特別是含有α7 nAChR亞單位的[13],且在 AD患者膽堿能系統損傷與α7nAChR水平下降和突觸損傷有關[14],其在調節神經可塑性方面具有獨特的作用,在神經保護中起了重要的作用。因此,α7nAChR成為近年來研究的熱點。課題組前期的研究也表明Aβ 能明顯降低大鼠及培養細胞中α7nAChR的表達,而α7nAChR基因表達沉默能通過影響細胞APP代謝相關酶的表達而增加Aβ的生成[15],二者互為因果。激活的α7nAChR可以調節神經元興奮性和神經遞質釋放、改變突觸可塑性,對維持記憶及認知功能的十分重要[16]。PNU是α7nAChR激動劑中特異性較高的合成物,與其他nAchR亞單位結合微乎其微,與α7nAChR結合,提高和改善AD動物模型的認知和記憶功能[17]。
突觸是大腦中信息存儲和傳遞的基本結構和功能單位。突觸囊泡再循環是腦內的神經遞質進行釋放和循環再利用的一個關鍵過程,它有兩種再循環形式:其一即通過形成短暫存在的融合孔使神經遞質從突觸囊泡釋放;其二為通過網格蛋白介導使突觸囊泡與突出前膜融合以胞吐的形式釋放神經遞質,然后仍然通過網格蛋白介導將神經遞質以胞吞的形式重新吸收形成新的突觸囊泡,完成突觸囊泡再循環。目前尚沒有證據表明前一種循環形式對突觸功能有明顯影響,當前的研究主要以網格蛋白介導的胞吐和胞吞形式為主要模式[7]。AP180是一種神經特異性的蛋白,是網格蛋白包被的主要成分之一,它參與了網格蛋白包被過程,同時也調節著新形成的突觸囊泡的大小。
盡管AD的主要病理學改變是Aβ沉積形成的老年斑和神經元纖維纏結,但近年的研究結果表明突觸丟失是認知功能低下的主要危險因素之一。突觸丟失致使突觸聯系障礙,神經元與神經元之間信息傳遞中斷,學習、記憶和認知能力下降。研究顯示[18,19],AD患者和動物模型的腦組織中存在一定程度的突觸關聯中斷、網格蛋白介導的突觸囊泡內吞功能降低以及神經細胞突觸回收囊泡的能力下降。通過對AD患者尸體解剖研究證實,AD腦組織突觸功能低下。但引起AD腦中突觸丟失及功能低下的具體機制目前還不十分清楚。本實驗主要研究特異性激動APP/PS1轉基因小鼠腦組織中α7nAChR水平后對海馬組織網格蛋白AP180蛋白的影響,從而探討α7nAChR在阿爾茨海默病(Alzheimer disease,AD) 發病機制中的神經保護作用機制。本研究選用24周齡的APP/PS1雙轉基因模型小鼠,行為學Morris水迷宮實驗結果表明:APP/PSl雙轉基因組小鼠和非轉基因組小鼠都具有一定的空間記憶能力,但24周齡APP/PSl雙轉基因小鼠空間探索實驗結果與同月齡小鼠有差異,說明24周齡APP/PSl雙轉基因小鼠學習記憶能力出現下降,這和APP/PSl雙轉基因小鼠行為學特點一致。實驗結果顯示APP/PS1雙轉基因模型小鼠海馬組織中AP180蛋白表達水平下降,表明在AD早期即已出現突觸蛋白表達含量下降。此外,本實驗結果還顯示,特異性激動α7nAChR水平后,與對照組相比WP組小鼠大腦海馬組織中AP180蛋白表達水平升高;與APP/PS1組相比AP組小鼠大腦海馬組織中AP180蛋白表達水平明顯升高。因此,我們推測可能原因是α7nAChR對突觸有一定的神經保護作用,α7nAChR與突觸密切相關。因而,進一步研究α7nAChR對突觸活動的調控有重要的研究意義,為AD的早期診治提供一定的幫助。
[1]余 抒,府偉靈.阿爾茨海默病及診斷學研究進展[J].臨床檢驗雜志,2016,34(1):49-51.
[2]Upadhaya AR,Capetillo-Zarate E,Kosterin I,et al.Dispersible amyloid β-protein oligomers, protofibrils,and fibrils represent diffusible but not soluble aggregates their role in neurodegeneration in amyloid precursor protein (APP) transgenic mice[J].Neurobiol Aging,2012,33(11):2641-2660.
[3]沈 陽,陳 宇,傅潔瑜,等.老年癡呆癥的分子機制[J].生命科學,2014,6:550-559.
[4]董陽婷,譚龍春,官志忠.β-淀粉樣肽對大鼠原代海馬神經細胞中SIRTs表達的影響[J].貴陽醫學院學報,2016,41(12):1376-1381.
[5]Parri H,Hernandez CM,Dineley KT,et al.Research update:Alpha7 nicotinic acetylcholine receptor mechanisms in Alzheimer’s disease[J].Biochem Pharmacol,2011,82(8):931.
[6]景 鑫,田 新,黃金蘭,等.氧化應激在阿爾茨海默病中的作用及相關藥物研究[J].生物技術世界,2015,10:157-159.
[7]夏志明,胡雅兒.突觸囊泡再循環與阿爾茨海默病[J].生理科學進展,2012,43(1):5-10.
[8]Selkor DJ.Alzheimer’s disease: genes,Proteins,and therapy [J].Physiol Rev,2001,81:741-766.
[9]Medeiros R,Castello NA,Cheng D,et al.α7 nicotinic receptor agonist enhances cognition in aged 3xTg-AD mice with robust plaques and tangles[J].Am J Pathol,2014,184:520-529.
[10]Taly A,Corringer PJ,Guedin D,et al.Nicotinic receptors: allosteric transitions and therapeutic targets in the nervous system[J].Nat Rev Drug Discov,2009,8(9):733-750.
[11]Cao Y,Xiao Y,Ravid R,et al.Changed clathrin regulatory proteins in the brains of Alzheimer’s disease patients and animal models[J].J Alzheimer’s Dis,2010,22(1):329-342.
[12]Tanzi RE.The synaptic abeta hypothesis of Alzheimer’s disease[J].Nat Neurosci,2005,8(8):977-979.
[13]Cao Y,Xiao Y,Ravid R,et al.Changed clathrin regulatory proteins in the brains of Alzheimer’s disease patients and animal models[J].J Alzheimer’s Dis,2010,22(1):329-342.
[14]曹 穎,廖 媛,肖 雁,等.β淀粉樣肽對人SH-SY5Y細胞突觸素、發動蛋白及銜接蛋白180表達的影響[J].中華神經醫學雜志,2011,10(8):774-778.
[15]Haass C,Selkoe DJ.Soluble p rotein oligomers in neur odegeneration:less ons from the Alzheimer’s amyloid β2pep tide[J].Nat Rev Mol Cell Biol,2007,8(2):101-112.
[16]Nordman JC,PhilipsWS,Kodama N,et al.Axon targeting of the alpha 7 nicotinic receptor in developing hippocampal neurons by Gprinl regulates growth[J].J Neurochemist,2014,129(4):649-662.
[17]Vicens P,Heredia L,Torrente M,et al.Behavioural effects of PNU-282987 and stress in an animal model of Alzheimer’s disease[J].Psychogeriatrics,2017,17(1):33-42.
[18]Yao PJ,Zhu M,Pyun EI,et al.Defects in expression of genes related to synaptic vesicle trafficking in frontal cortex of Alzheimer’s disease[J].Neurobiol Dis,2003,12(2):97-109.
[19]Pham E,Crews L,Ubhi K,et al.Progressive accumulation of amyloid-beta oligomers in Alzheimer’s disease and in amyloid precursor protein transgenic mice is accompanied by selective alterations in synaptic scaffold proteins[J].Federation of European Biochemical Societies,2010,277(14):3051-3067.
Theeffectsexpressionofα7neuronalnicotinereceptoragonistPNU282987onAP180proteininhippocampusofAPP/PS1Mice
DENGYuxin,WANGXiaoling,WUChangxue,etal.
[KeyLaboratoryofMedicalMolecularBiology(GuizhouMedicalUniversity),Guiyang550004,China]
ObjectiveTo investigate the expression of AP180 protein in hippocampus of specific agonized APP/PS1 transgenic mice after specific activation of α7nAChR.Methods16 APP/PS1 mice were randomly divided into APP/PS1 group (APP/PS1) and APP/PS1+PNU282987 group (AP),8 mice in each group;16 wild-type mice were randomly divided into wild group (Control) and wild+PNU282987 group (WP),each group of 8.Mice were reached after 24 weeks of age,Morris water maze behavioral test,Real-time PCR and Western blotting were used to detect AP180 mRNA expression and protein level in the hippocampus of mice in each group.ResultsCompared with the control group,AP180 mRNA and protein expression in the hippocampus of APP/PS1 group were significantly decreased (P<0.01,P<0.01).Compared with the control group,the expression of AP180 mRNA and WP1 in the WP group was significantly higher than that in the control group (P<0.01,P<0.05).Compared with APP/PS1 group,AP180 mRNA and protein levels in hippocampus of AP group were significantly increased (P<0.01,P<0.01).ConclusionThe specific activation of α7 nAChR in hippocampus of APP/PS1 transgenic mice could increase the expression level of key protein AP180 in the endocytosis of synovial vesicles mediated by cathepsin.This may suggest that α7nAChR has a protective effect on synapses,further indicating that α7nAChR plays an important role in the pathogenesis of Alzheimer’s disease.
α7 nAChR; AD; APP/PS1 transgenic mouse; AP180
R749.1
A
1003-2754(2017)10-0872-04
2017-08-17;
2017-10-03
國家自然科學基金(81360178);貴州省重大專項計劃[黔科合重大專項字(2014)6008號];貴州省創新計劃項目[黔教合協同創新中心(2014)06];貴州省科技創新人才團隊[黔科通(2016)161號]
[1.地方病與少數民族疾病教育部重點實驗室(貴州醫科大學),貴州 貴陽 550004;2.貴州省醫學分子生物學重點實驗室(貴州醫科大學),貴州 貴陽 550004;3.貴州醫科大學病理學教研室,貴州 貴陽 550004]
齊曉嵐,E-mail:xiaolan76@163.com
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
作文周刊·小學二年級版(2022年20期)2022-05-05 01:33:06
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
學苑創造·A版(2020年9期)2020-10-13 09:41:02
創新作文(小學版)(2019年10期)2019-09-25 08:12:28
人大建設(2019年12期)2019-05-21 02:55:32
小學生學習指導(低年級)(2017年5期)2017-05-04 04:14:38
作文與考試·小學高年級版(2015年17期)2015-05-30 10:48:04
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
