HOG1對格特隱球菌應對壓力應激、毒力因子和抗藥性的影響
2017-11-06 06:08:27皇幼明樊一斌陶小華潘衛利
中國麻風皮膚病雜志 2017年10期
皇幼明 樊一斌 陶小華 潘衛利
·論著·
浙江省人民醫院皮膚科,杭州醫學院附屬人民醫院,浙江杭州,310014
潘衛利,E-mail:doctorpan163@163.com
HOG1對格特隱球菌應對壓力應激、毒力因子和抗藥性的影響
皇幼明 樊一斌 陶小華 潘衛利
目的明確HOG1基因對格特隱球菌應對壓力應激、毒力因子產生和抗藥性的影響。方法比較格特隱球菌原始株、hog1Δ菌株和重建株在含高滲透壓培養基、抗真菌藥物培養基中的生長差異,以及在YEPD培養基中莢膜的合成和咖啡因培養基中黑素產生的差異。結果hog1Δ菌株在高壓力和抗真菌培養基中生長受限,在YEPD培養基中莢膜合成減弱,在咖啡因培養基中黑素生成減少。結論HOG1基因在格特隱球菌應對壓力應激、毒力因子產生、抗藥性中有重要作用。
格特隱球菌; HOG1基因; HOG-MAPK通路
隱球菌是擔子菌類酵母真菌微生物,在自然界分布廣泛。據統計,每年隱球菌病例約100萬例,致死率50%以上,在非洲艾滋病患者中,隱球菌病已超過結核病成為最主要的致病病因[1]。隱球菌在宿主中生存和繁殖必須要適應外界環境應激如高滲、氧化反應。隱球菌對環境應激的調節通過多種信號途徑來完成,而HOG-MAPK信號途徑是其中之一。新生隱球菌是隱球菌病的主要致病菌,國外研究表明,該途徑同樣參與調控新生隱球菌對抗菌藥物、溫度等應激反應[2],同時也調控兩個重要的隱球菌毒力因子——莢膜和黑素的生成[3]。而作為它的姐妹種屬,格特隱球菌在形態特征、生化性質方面均有所不同。本文將通過比較格特隱球菌HOG1基因敲除后菌株對外界應激反應、毒力因子、對抗真菌藥物的敏感性等方面的變化來驗證HOG1在隱球菌致病機制中的作用。
1 材料和方法
1.1 材料
1.1.1 菌株 格特隱球菌(血清B型)臨床株CZ2012原始株、hog1Δ菌株(缺陷株)和重建株(均來自第二軍醫大學隱球菌保藏中心)。
1.1.2 培養基 RPMI1640培養液、SDA及改良型SDA培養基、YEPD固體培養基、咖啡酸玉米培養基、尿素瓊脂培養基等均按照標準配方配制。
1.1.3 主要試劑 兩性霉素B、酮康唑、伊曲康唑、氟康唑和5氟胞嘧啶。
1.1.4 主要儀器和器材 2.5 μL、10 μL、100 μL、1000 μL 各式加樣器;0.02 μm孔徑過濾器濾頭;pH計;血細胞計數板。
2 方法
2.1 對外界應激如滲透壓、溫度、氧化反應測試 配制用于不同應激反應測試的YEPD培養基(壓力測試:含1 mol/L、1.5 mol/L KCl的YEPD固體培養基;氧化應激:含H2O25 mmol/mL的YEPD固體培養基;含20%甘油、20%葡萄糖、20%甘油果糖的YEPD培養基),配制各組菌懸液,并依次倍比稀釋,取5 μL菌懸液點種相應的培養基中,3~5天后觀察結果。
2.2 莢膜和黑素生成的測定 配制各組菌液,調整菌懸液至5×106個/mL,點種改良型SDA平板上,分別置于30℃、37℃條件下培養48 h,然后挑取菌落制備菌懸液,墨汁染色,顯微鏡下觀察,拍照。利用photoshop軟件計算菌體莢膜相對大小。然后SPSS 17.0軟件進行分析;取同種菌液,調整同樣濃度,點種于固體咖啡酸培養基上,分別于30℃、37℃溫度下培養3 d、7 d后觀察結果。
2.3 對尿素酶生成的影響 調整菌懸液濃度至5×106個/mL,離心,PBS純化3次后,每組取5 μL點種于尿素培養基(Christensen urea agar),重復三次,分別30℃和37℃條件下培養,1~2 d后觀察結果。
3 統計學分析
本實驗中主要通過測量原始株、hog1Δ缺陷株和重建株之間的莢膜相對面積比,分析HOG1基因對莢膜的影響,因此采用單因素方差分析(one-way ANOVA analysis)比較三種菌株之間的差異,若三種菌株間存在差異(P<0.05),之后采用Bonfferoni檢驗方法比較兩兩菌株間的差異;本實驗所用統計軟件為SPSS 17.0。
4 結果
4.1 HOG1對外界應激如滲透壓、溫度、氧化反應的影響
4.1.1 三種菌株對5 mmol/L H2O2沒有表現出明顯的差異,hog1Δ菌株對1 mol/L KCl、1.5 mol/L KCl的抵抗能力明顯減弱(圖1)。
4.1.2hog1Δ菌株對抵抗唑類藥物應激的敏感性升高,而5氟胞嘧啶的敏感性差別較小(圖2)。
4.1.3hog1Δ菌株在高濃度甘油、高糖、高甘露醇應激下活力下降(圖3)。
4.2 HOG1對莢膜和黑素生成的有正向調節作用
4.2.1 莢膜觀察 在30℃和37℃條件下,hog1Δ菌株的莢膜生成能力明顯減弱(圖4、5)。
同樣地,比較莢膜相對面積大小(30℃:WT 0.45、hog1Δ0.13、HOG1+hog1Δ0.41;37℃:WT0.43、hog1Δ0.10、HOG1+hog1Δ0.36),在30℃和37℃條件下,hog1Δ菌株在同等條件下莢膜相對面積較小,而原始株和重建株莢膜相比,差異有顯著意義(圖6;*:P<0.001,vshog1Δgroup)。
4.2.2 黑素生成比較 格特隱球菌CZ2012hog1Δ菌株與原始株和重建株相比,hog1Δ菌株在不同溫度和不同時間點的黑素生成均明顯減少(圖7) 。
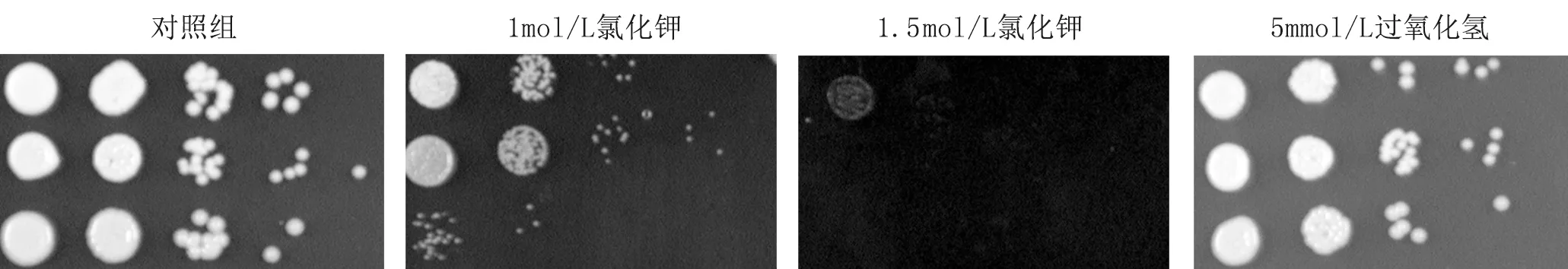
圖1 上至下三種菌株分別為WT、hog1Δ、HOG1+hog1Δ
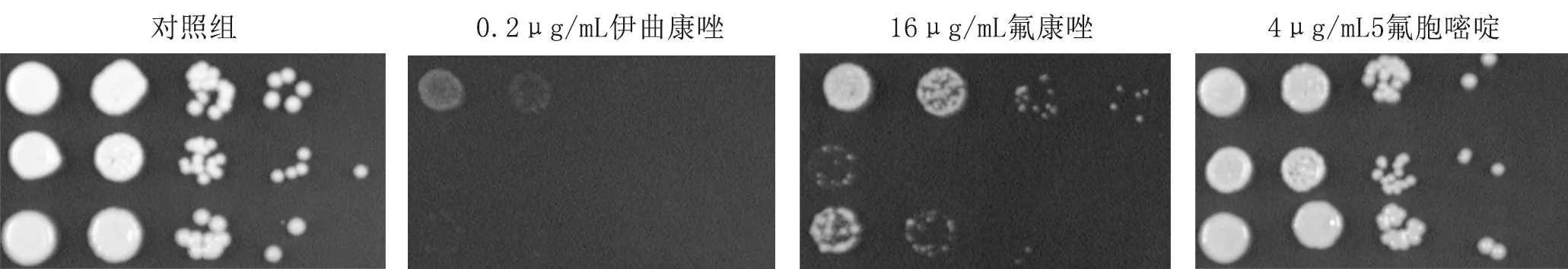
圖2 上至下三種菌株分別為WT、hog1Δ、HOG1+ hog1Δ
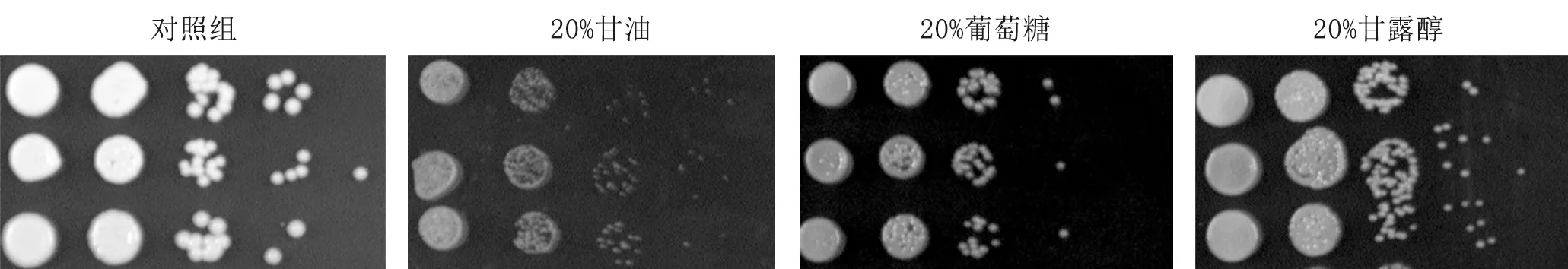
圖3 上至下三種菌株分別為WT、hog1Δ、HOG1+ hog1Δ
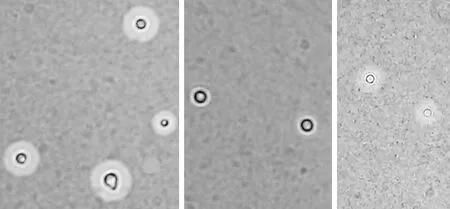
圖4 30℃條件下,hog1Δ菌株的莢膜生成能力明顯減弱
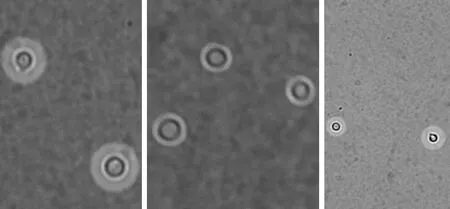
圖5 37℃條件下,hog1Δ菌株的莢膜生成能力明顯減弱
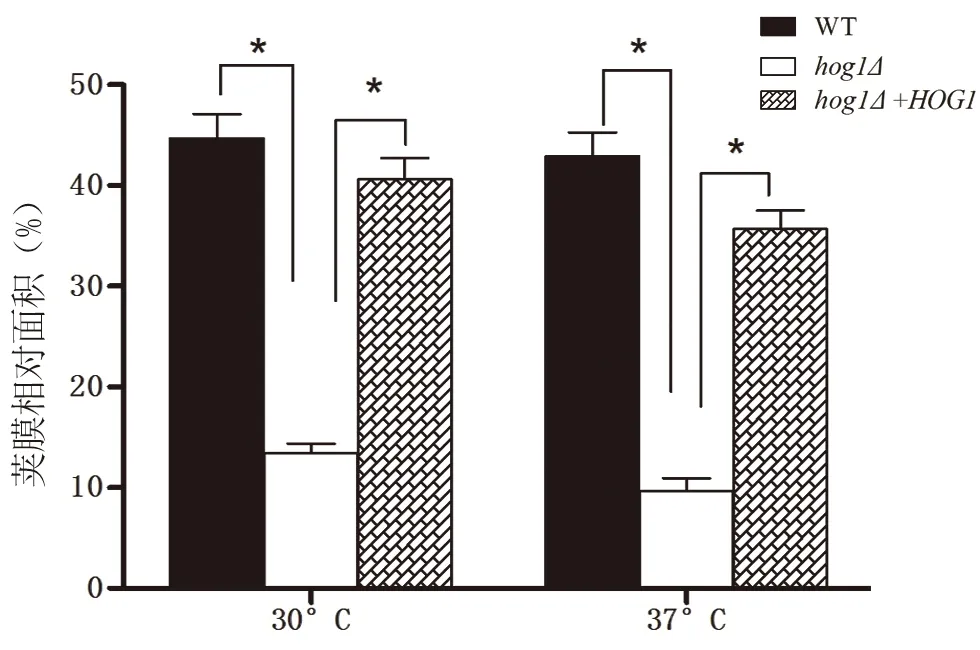
圖6 在30℃和37℃條件下,hog1Δ菌株在同等條件下莢膜相對面積較小
5 討論
格特隱球菌主要感染免疫正常人群。該病原菌以前被認為主要分布在熱帶和亞熱帶地區,而近來國內也有越來越多的格特隱球菌感染病例報道[4]。HOG-MAPK信號通路在國外的研究主要限于新生隱球菌,國內關于格特隱球菌致病性研究的試驗研究尚少。在本研究中,我們通過比較格特隱球菌原始株、hog1Δ菌株及重建株的毒力方面的差異來驗證HOG1在格特隱球菌致病中的作用。
通過觀察hog1Δ菌株在正常條件和高滲條件下的生長情況,我們發現hog1Δ菌株對外部高滲透性離子、20%糖、20%甘露醇均表現為對高滲透壓的抵抗力減弱,表明hog1Δ菌株對高滲透壓表現出較強的敏感性。Jung等[5]在新生隱球菌H99中發現,hog1Δ菌株在富含高糖的高Na+、K+的YEPD培養基表現出明顯的差別,低糖、低PH的高滲培養基中生長顯著受限。所以,我們認為本實驗的hog1Δ菌株對外部高滲應激的抵抗力明顯降低,證實HOG1蛋白作為HOG-MAPK通路中重要的因子在細胞抵抗外界高滲壓力中發揮著重要的作用。在新生隱球菌中,HOG-MAPK在菌體抵抗多種氧化應激損害中有重要作用,本研究中,hog1Δ菌株并沒有表現出明顯的變化,提示在菌株中,HOG1在菌體對外界氧化應激反應中發揮的作用有限,這可能與隱球菌有多個信號途徑和調控機制在應對壓力刺激時發揮作用有關。

圖7 上至下三種菌株分別為WT、hog1Δ、HOG1+ hog1Δ
Hog1蛋白是HOG-MAPK通路的重要組分。目前已有國外相關學者對新生隱球菌該通路上游組分進行了相關研究[5,6]。莢膜和黑素主要由cAMP信號通路調控,是隱球菌區別于其他真菌的兩個重要的毒力特征,HOG-MAPK信號通路通過與cAMP交叉作用參與莢膜和黑素的生成[7]。該通路的任何環節的突變都可能會造成莢膜和黑素生成的改變以及毒力的變化。而本實驗hog1Δ缺陷株卻表現為莢膜和黑素的合成明顯減少,表明HOG1可以正向調節莢膜和黑素的合成。這與新生隱球菌的作用相反[7],而同樣的研究中,新生隱球菌不同菌株間HOG1對莢膜和黑素的調控作用也存在差異,證明隱球菌種屬間HOG1對莢膜和黑素的調控機制存在差異,可能HOG1基因缺失后引起HOG-MAPK 通路上下游組分發生變化或同時存在其他平行途徑同時調控這莢膜和黑素的生成。
兩性霉素B和唑類作為經典抗真菌藥物常被用于隱球菌病的一線治療。HOG-MAPK途徑控制菌體細胞麥角固醇的合成,從而影響對兩性霉素B和唑類抗真菌藥物的敏感性。已有研究表明高滲透壓力可導致真菌多種麥角固醇基因合成減少[8],這與我們在臨床藥物治療后對隱球菌菌體在電鏡下觀察到的結果相似。低麥角固醇可引起細胞膜緊縮,靈活性減小,導致胞內甘油等滲透性物質流出減少。HOG-MAPK通路可能通過影響麥角固醇的生成,使新生隱球菌對抗真菌藥物的敏感性產生影響。新生隱球菌耐藥機制復雜,臨床分離的耐藥菌株多與菌體內麥角固醇生成有關[9],可能是該成分主要影響抗真菌藥物(如多烯類和三唑類)的結合位點和細胞膜的通透性有關。本實驗中, CZ2012hog1Δ菌株表現出對兩性霉素B和5FC的敏感性減弱,而對唑類藥物的敏感性增強,這與國外新生隱球菌的情況正好相反[10]。說明在格特隱球菌中可能存在其他的途徑共同調節麥角固醇的合成進而影響抗真菌藥物的敏感性,具體的機制尚需我們進一步進行研究。
總之,本研究主要從體外毒力表型來證實HOG1對新生隱球菌毒力調控的影響。結果HOG1基因敲除后,格特隱球菌對外界應激反應有明顯變化,相關毒力因子如莢膜和黑素合成減少,對兩性霉素B、5-FC的敏感性減弱,而對唑類藥物的敏感性增強。因此,我們認為,HOG1在格特隱球菌應對外界應激變化和毒力調控中發揮著重要作用,但與其在新生隱球菌中的調控機制存在差異性,對該通路中相關靶點的具體作用機制尚需進一步的分子研究來證實。
[1] Park BJ, Wannemuehler KA, Marston BJ, et al. Estimation of the current global burden of cryptococcal meningitis among persons living with HIV/AIDS[J]. AIDS,2009,23(4):525-530.
[2] Lee KT, Byun HJ, Jung KW, et al. Distinct and redundant roles of protein tyrosine phosphatases Ptp1 and Ptp2 in governing the differentiation and pathogenicity of Cryptococcus neoformans[J]. Eukaryot Cell,2014,13(6):796-812.
[3] Huston SM, Ngamskulrungroj P, Xiang RF, et al. Cryptococcus gattii capsule blocks surface recognition required for dendritic cell maturation independent of internalization and antigen processing[J]. J Immunol,2016,196(3):1259-1271.
[4] Fang W, Fa Z, Liao W. Epidemiology of Cryptococcus and cryptococcosis in China[J]. Fungal Genet Biol,2015,78:7-15.
[5] Jung KW, Strain AK, Nielsen K, et al. Two cation transporters Ena1 and Nha1 cooperatively modulate ion homeostasis, antifungal drug resistance, and virulence of Cryptococcus neoformans via the HOG pathway[J]. Fungal Genet Biol,2012,49(4):332-345.
[6] Kim SY, Ko YJ, Jung KW, et al. Hrk1 plays both Hog1-dependent and -independent roles in controlling stress response and antifungal drug resistance in Cryptococcus neoformans[J]. PLoS One,2011,6(4):e18769.
[7] Bahn YS, Kojima K, Cox GM, et al. Specialization of the HOG pathway and its impact on differentiation and virulence of Cryptococcus neoformans[J]. Mol Biol Cell,2005,16(5):2285-2300.
[8] Wang M, Chen C, Zhu C, et al. Os2 MAP kinase-mediated osmostress tolerance in Penicillium digitatum is associated with its positive regulation on glycerol synthesis and negative regulation on ergosterol synthesis[J]. Microbiol Res,2014,169(7-8):511-521.
[9] Ngamskulrungroj P, Chang Y, Hansen B, et al. Characterization of the chromosome 4 genes that affect fluconazole-induced disomy formation in Cryptococcus neoformans[J]. PLoS One,2012,7(3):e33022.
[10] Ko YJ, Yu YM, Kim GB, et al. Remodeling of global transcription patterns of Cryptococcus neoformans genes mediated by the stress-activated HOG signaling pathways[J]. Eukaryot Cell,2009,8(8):1197-1217.
TheroleofHOG1geneinstress,virulencefactorsandanti-fungaldrugsensitivityofCryptococcusgattii
HUANGYouming,FANYibin,TAOXiaohua,PANWeili.
DepartmentofDermatology,ZhejiangProvincialPeople'sHospital,People'sHospitalofHangzhouMedicalCollege,Zhejiang310014,China
PANWeili,E-mail:doctorpan163@163.com
Objective: To determine the influence of HOG1 gene on stress, virulence factors and anti-fungal drug sensitivity ofCryptococcusgattii.MethodsThe growth difference ofCryptococcusgattii(wild strain,hog1Δstrain and reconstruction strain) in high osmotic pressure medium and antifungal medium, and the bacterial capsule size in the YEPD medium and melanin production in the caffeine medium were compared.Resultshog1Δstrain showed growth limitation in high osmotic pressure medium and antifungal medium, reduction of capsule synthesis in YEPD medium and less melanin production in caffeine medium.ConclusionHOG1 gene plays an essential role in the environmental stress, production of virulence factors and antifungal drug sensitivity ofCryptococcusgattii.
Cryptococcusgattii; HOG1 gene; HOG-MAPK pathway
(收稿:2017-03-05 修回:2017-07-04)
