金堂黑山羊FSH信號調節對卵巢基因表達譜影響的研究
2017-12-01 09:55:24趙素君王秋實廖黨金
西南農業學報 2017年11期
曹 冶,趙素君,謝 晶,王秋實,廖黨金
(1.四川省畜牧科學研究院,四川 成都 610066;2. 動物遺傳育種四川省重點實驗室,四川 成都 610066)
金堂黑山羊FSH信號調節對卵巢基因表達譜影響的研究
曹 冶1,2,趙素君1,2,謝 晶1, 2,王秋實1,2,廖黨金1,2
(1.四川省畜牧科學研究院,四川 成都 610066;2. 動物遺傳育種四川省重點實驗室,四川 成都 610066)
【目的】本研究旨在搞清楚金堂黑山羊FSH信號調節機制的作用模式。【方法】將10只生殖周期相同的健康空懷的金堂黑山羊成年母羊隨機分成2組,一組為對照組,一組為試驗組。在發情后的第2天對試驗組金堂黑山羊進行FSH肌肉注射100IU,并在注射24 h后,摘取金堂黑山羊的卵巢;對照組發情后的第3天摘取卵巢。分別提取它們的總RNA,進行表達譜測序。【結果】經測序分析:對照組和試驗組分別得到有效序列30 906 038和27 763 272條; 經統計分析后,得到歸一化序列30 216 946和27 124 882條,與山羊基因組的符合率分別達85.30 %和86.13 %。比對后發現239個基因出現顯著性差異表達,其中139個基因上調了表達,100個基因下調了表達。GO功能分類發現在86個生物學過程、細胞組分、分子功能出現了顯著性差異。KEGG功能分類發現有6個調節途徑發生了顯著性改變。【結論】FSH信號強度變化作用在卵巢上主要通過調節卵巢激素分泌、調節卵巢上FSH信號傳導通路、調節脂代謝來響應,從而調節了動物的繁殖性能。
金堂黑山羊;FSH信號;卵巢;基因表達譜
【研究意義】金堂黑山羊屬我國優良的地方黑山羊品種,其繁殖力顯著高于其他一些山羊品種,據報道金堂黑山羊的產羔數達2.40,大大高于世界上其它優秀山羊品種[1]。【前人研究進展】在前期研究中,發現它的繁殖器官中有兩條FSH受體基因(FSHR)同時表達,通過形成異源二聚體顯著增加了FSH信號強度,促進了排卵,從而表現出高繁殖力。基于以上研究,可以發現動物高繁殖力的主要控制因素在于FSH信號通路。然而,由于FSH是由腦垂體分泌的激素,經血液循環進入靶器官,進行生殖調節,在這個過程中,該信號通路會受到較多的條件因素(如FSH劑量、FSH受體表達、FSH與受體結合以及結合后下游因子作用等)影響。這就使得FSH信號作用在卵巢進行排卵時出現差異,進而使得家畜在每一胎產仔時,產仔數量卻存在差異。這說明動物機體對FSH信號通路的調節存在著一定的調節機制,可能是受研究手段的限制,到目前為止尚未見這方面的詳細報道。【本研究切入點】只有完全搞清楚這個調節機制的作用模式,才能有效利用山羊中影響產羔數的主效基因和有效的分子標記,進行山羊高產仔數的分子育種選擇,在分子水平研究金堂黑山羊FSH信號通路的調節機制,對于優良地方品種的高效選育和改良具有十分重要意義。【擬解決的關鍵問題】因此,通過改變金堂黑山羊的FSH信號強度,來測定卵巢基因表達譜的變化,為徹底搞清楚FSH信號的調節機制打下堅實基礎。
1 材料與方法
1.1 實驗動物
在金堂黑山羊原種場隨機選取年齡在3~5歲,體重30~40 kg,營養良好,前3胎產仔數在2個以上的,生殖周期相同的健康空懷的金堂黑山羊成年母羊10只作為試驗用羊,供試羊在欄內散放飼養,自由采食、飲水。實驗前,觀察發情情況,連續觀察30 d。
1.2 主要試劑
Trizol為Invetrogen公司(美國)產品;FSH購自于寧波激素一廠;化學試劑除注明外,均為國產分析純。
1.3 FSH處理
將10只試驗用羊隨機分成2組,每組5只羊。一組為對照組,不進行FSH注射。另一組為試驗組。在發情后的第2天對試驗組金堂黑山羊進行FSH肌肉注射,注射劑量為100 IU,并在注射24 h后,摘取金堂黑山羊的卵巢。同時在對照組發情后的第3天摘取卵巢。卵巢液氮速凍后在-80 ℃條件下保存,用于基因表達譜分析。
1.4 總 RNA 提取
用Trizol試劑提取卵巢總RNA。提取步驟按照操作說明進行。將提取的卵巢總RNA送上海美吉生物有限公司進行表達譜測序分析。將得到的數據進行質控分析,包括A/T/G/C堿基含量分布統計、堿基質量分布分析、飽和度分析、冗余序列分析以及基因覆蓋度分析,從而確定表達譜測序分析的質量。
1.5 GO(Gene Ontology)功能分類和KEGG PATHWAY代謝途徑分析
將測得的基因表達譜用軟件Goatools (https://github.com/tanghaibao/GOatools) 進行GO功能富集分析,使用Fisher精確檢驗。為控制計算的假陽性率,使用多重檢驗方法(Bonferroni)對P值進行了校正,通常情況下,當經過校正的P值(P_fdr)≤0.05時,認為此GO功能存在顯著富集情況[2]。將測得的基因表達譜用KOBAS(http://kobas.cbi.pku.edu.cn/home.do) 進行KEGG PATHWAY 富集分析,使用Fisher 精確檢驗進行計算。為控制計算假陽性率,采用BH(FDR) 方法進行多重檢驗,經過校正的P值(CorrectedP-Value)以0.05 為閾值,滿足此條件的KEGG 通路定義為在差異表達基因中顯著富集的KEGG 通路[3]。
2 結果與分析
2.1 基因表達
本研究采用RNA-seq技術對兩組經不同FSH處理的金堂黑山羊卵巢樣品進行表達譜測序。
分別得到有效序列30906038和27763272條。將這些數據進行A/T/G/C堿基含量分布統計,發現該文庫堿基分布均勻,模糊堿基N %在合理范圍之內。同時,進行堿基質量分布統計,得出一個綜合的值,即質量值(Q),用來評估測序的質量好壞,結果見表1,發現所獲得測序數據達到后續分析要求。堿基錯誤率分布統計分析發現數據的堿基錯誤率均可以達到低于0.1 %。經統計分析后,得到歸一化序列30216946和27124882條。
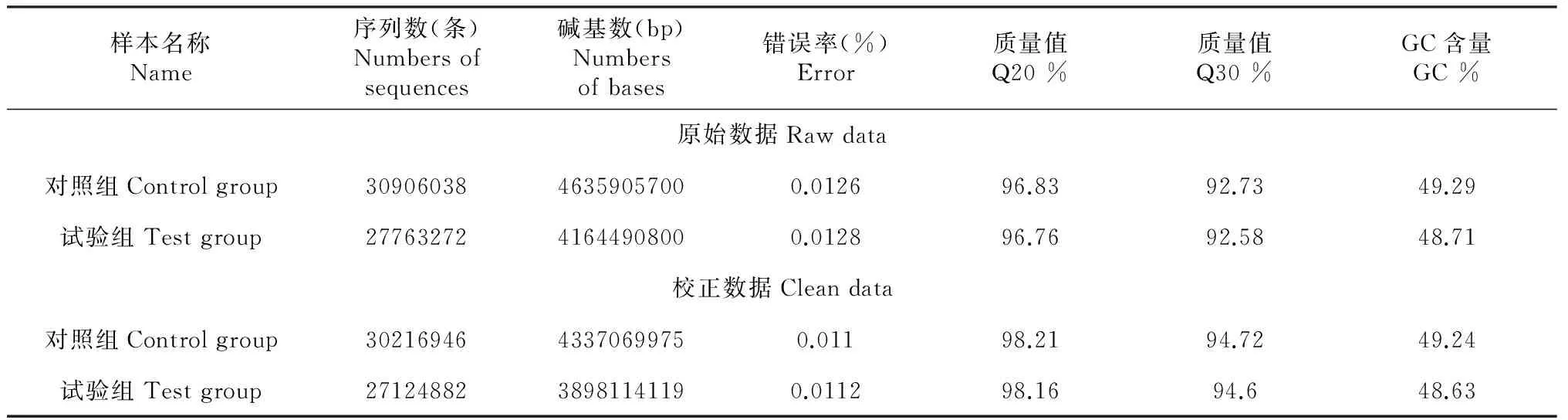
表1 測序數據統計

表2 與參考基因組比對
將質控后得到的高質量測序序列使用(http://tophat.cbcb.umd.edu/)與指定的參考基因組比對,參考基因組來自于http://www.ncbi.nlm.nih.gov/genome/genomes/10731。如表2所示,發現符合率分別達85.30 %和86.13 %。
分析顯示飽和度總體質量較高,該測序量能夠覆蓋絕大多數的表達基因。冗余序列分析顯示此樣本冗余序列含量正常。基因覆蓋度分析顯示結果不具有偏向性,結果較均一。
2.2 表達差異分析
將這些序列進行比對后發現239個基因出現顯著性差異表達,有139個基因上調了表達,100個基因下調了表達。其中,gene17948(muscle, skeletal, receptor tyrosine kinase)上調表達最高,提高了127倍; gene43088(Glutathione S-transferase A2 (EC 2.5.1.18)(Glutathione S-transferase alpha-2)(GST class-alph [...])下調表達最多,下調了144倍。
2.3 差異表達基因的GO功能分類
對這些差異表達基因進行了GO功能分類,發現在86個生物學過程、細胞組分、分子功能出現了顯著性差異(表3)。其中細胞組分GO功能顯著差異11個[極顯著差異7個(P≤0.01),差異顯著4個(P≤0.05)],分子功能GO功能顯著差異11個[極顯著差異8個(P≤0.01),差異顯著3個(P≤0.05)],生物學過程GO功能顯著差異64個[極顯著差異45個(P≤0.01),差異顯著19個(P≤0.05)]。在生物學過程GO功能中,涉及類固醇激素合成代謝的過程有11個極顯著差異;涉及脂肪合成代謝的過程有10個,其中極顯著差異7個(P≤0.01),顯著差異3個(P≤0.05);涉及小分子合成代謝的過程有3個極顯著差異(P≤0.01);涉及物質轉運過程6個顯著差異(P≤0.05)。
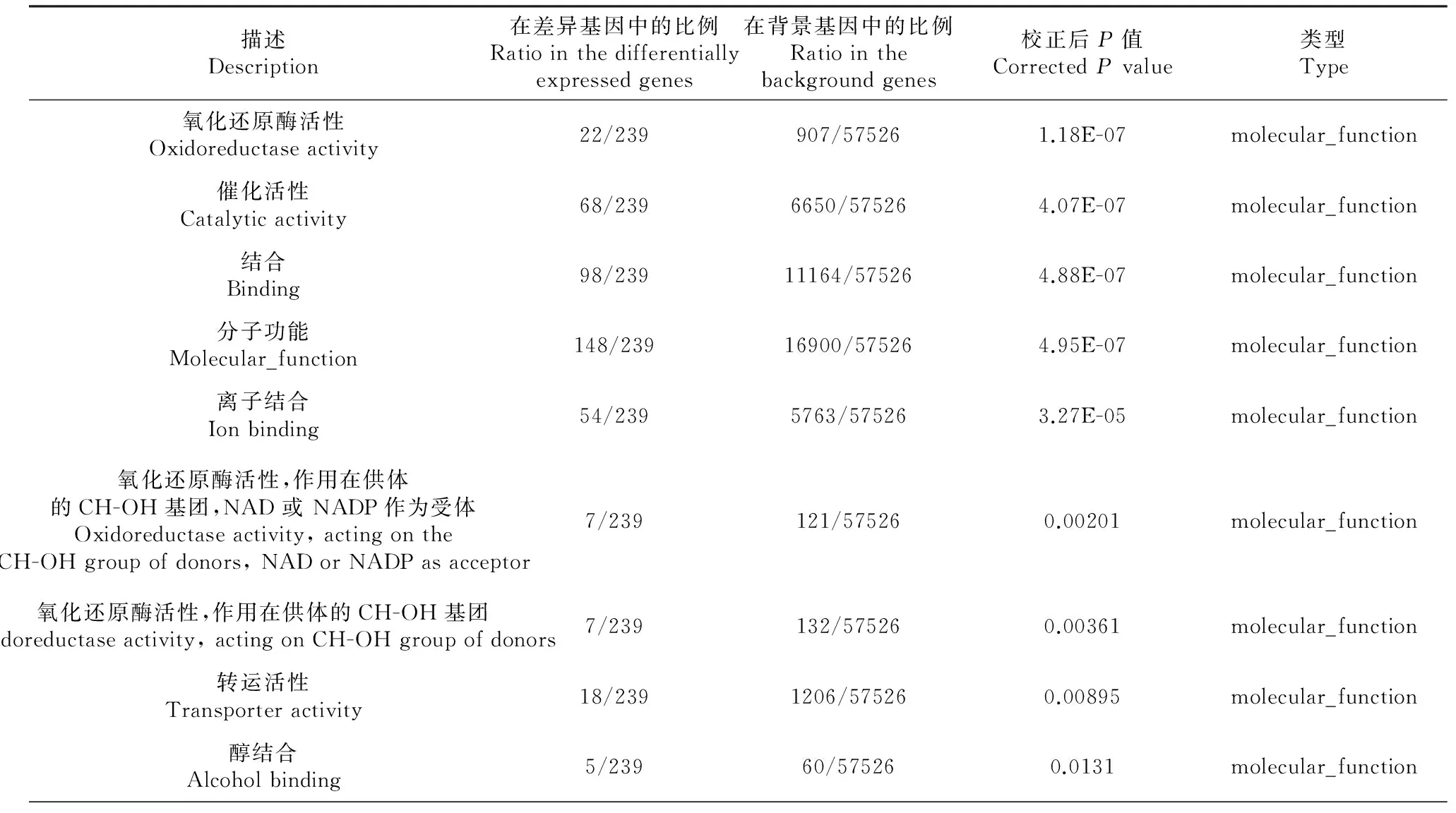
表3 金堂黑山羊卵巢差異表達基因的GO功能分類
續表3 Continued table 3
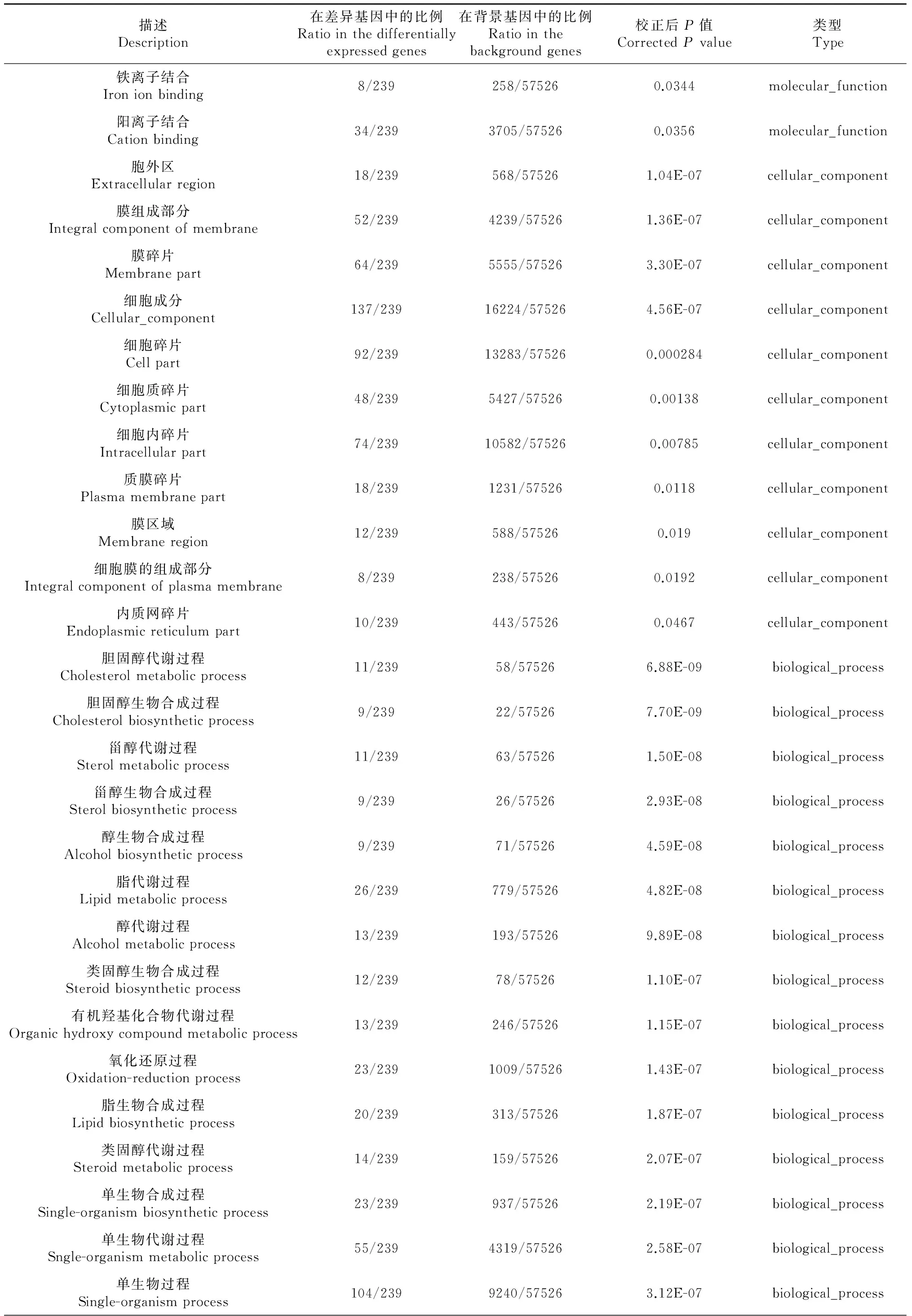
描述Description在差異基因中的比例Ratiointhedifferentiallyexpressedgenes在背景基因中的比例Ratiointhebackgroundgenes校正后P值CorrectedPvalue類型Type鐵離子結合Ironionbinding8/239258/575260.0344molecular_function陽離子結合Cationbinding34/2393705/575260.0356molecular_function胞外區Extracellularregion18/239568/575261.04E-07cellular_component膜組成部分Integralcomponentofmembrane52/2394239/575261.36E-07cellular_component膜碎片Membranepart64/2395555/575263.30E-07cellular_component細胞成分Cellular_component137/23916224/575264.56E-07cellular_component細胞碎片Cellpart92/23913283/575260.000284cellular_component細胞質碎片Cytoplasmicpart48/2395427/575260.00138cellular_component細胞內碎片Intracellularpart74/23910582/575260.00785cellular_component質膜碎片Plasmamembranepart18/2391231/575260.0118cellular_component膜區域Membraneregion12/239588/575260.019cellular_component細胞膜的組成部分Integralcomponentofplasmamembrane8/239238/575260.0192cellular_component內質網碎片Endoplasmicreticulumpart10/239443/575260.0467cellular_component膽固醇代謝過程Cholesterolmetabolicprocess11/23958/575266.88E-09biological_process膽固醇生物合成過程Cholesterolbiosyntheticprocess9/23922/575267.70E-09biological_process甾醇代謝過程Sterolmetabolicprocess11/23963/575261.50E-08biological_process甾醇生物合成過程Sterolbiosyntheticprocess9/23926/575262.93E-08biological_process醇生物合成過程Alcoholbiosyntheticprocess9/23971/575264.59E-08biological_process脂代謝過程Lipidmetabolicprocess26/239779/575264.82E-08biological_process醇代謝過程Alcoholmetabolicprocess13/239193/575269.89E-08biological_process類固醇生物合成過程Steroidbiosyntheticprocess12/23978/575261.10E-07biological_process有機羥基化合物代謝過程Organichydroxycompoundmetabolicprocess13/239246/575261.15E-07biological_process氧化還原過程Oxidation-reductionprocess23/2391009/575261.43E-07biological_process脂生物合成過程Lipidbiosyntheticprocess20/239313/575261.87E-07biological_process類固醇代謝過程Steroidmetabolicprocess14/239159/575262.07E-07biological_process單生物合成過程Single-organismbiosyntheticprocess23/239937/575262.19E-07biological_process單生物代謝過程Sngle-organismmetabolicprocess55/2394319/575262.58E-07biological_process單生物過程Single-organismprocess104/2399240/575263.12E-07biological_process
續表3 Continued table 3
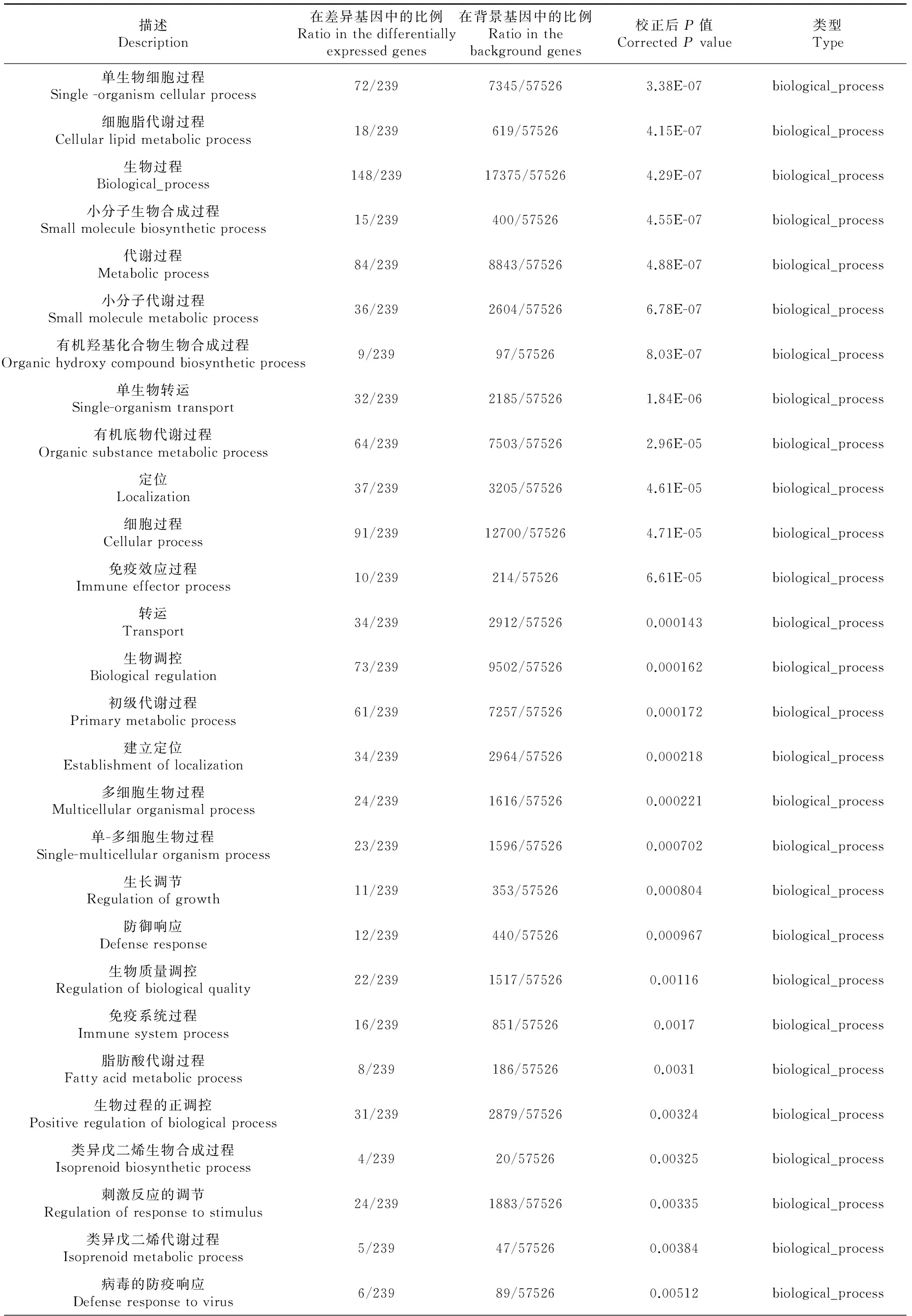
描述Description在差異基因中的比例Ratiointhedifferentiallyexpressedgenes在背景基因中的比例Ratiointhebackgroundgenes校正后P值CorrectedPvalue類型Type單生物細胞過程Single-organismcellularprocess72/2397345/575263.38E-07biological_process細胞脂代謝過程Cellularlipidmetabolicprocess18/239619/575264.15E-07biological_process生物過程Biological_process148/23917375/575264.29E-07biological_process小分子生物合成過程Smallmoleculebiosyntheticprocess15/239400/575264.55E-07biological_process代謝過程Metabolicprocess84/2398843/575264.88E-07biological_process小分子代謝過程Smallmoleculemetabolicprocess36/2392604/575266.78E-07biological_process有機羥基化合物生物合成過程Organichydroxycompoundbiosyntheticprocess9/23997/575268.03E-07biological_process單生物轉運Single-organismtransport32/2392185/575261.84E-06biological_process有機底物代謝過程Organicsubstancemetabolicprocess64/2397503/575262.96E-05biological_process定位Localization37/2393205/575264.61E-05biological_process細胞過程Cellularprocess91/23912700/575264.71E-05biological_process免疫效應過程Immuneeffectorprocess10/239214/575266.61E-05biological_process轉運Transport34/2392912/575260.000143biological_process生物調控Biologicalregulation73/2399502/575260.000162biological_process初級代謝過程Primarymetabolicprocess61/2397257/575260.000172biological_process建立定位Establishmentoflocalization34/2392964/575260.000218biological_process多細胞生物過程Multicellularorganismalprocess24/2391616/575260.000221biological_process單-多細胞生物過程Single-multicellularorganismprocess23/2391596/575260.000702biological_process生長調節Regulationofgrowth11/239353/575260.000804biological_process防御響應Defenseresponse12/239440/575260.000967biological_process生物質量調控Regulationofbiologicalquality22/2391517/575260.00116biological_process免疫系統過程Immunesystemprocess16/239851/575260.0017biological_process脂肪酸代謝過程Fattyacidmetabolicprocess8/239186/575260.0031biological_process生物過程的正調控Positiveregulationofbiologicalprocess31/2392879/575260.00324biological_process類異戊二烯生物合成過程Isoprenoidbiosyntheticprocess4/23920/575260.00325biological_process刺激反應的調節Regulationofresponsetostimulus24/2391883/575260.00335biological_process類異戊二烯代謝過程Isoprenoidmetabolicprocess5/23947/575260.00384biological_process病毒的防疫響應Defenseresponsetovirus6/23989/575260.00512biological_process
續表3 Continued table 3
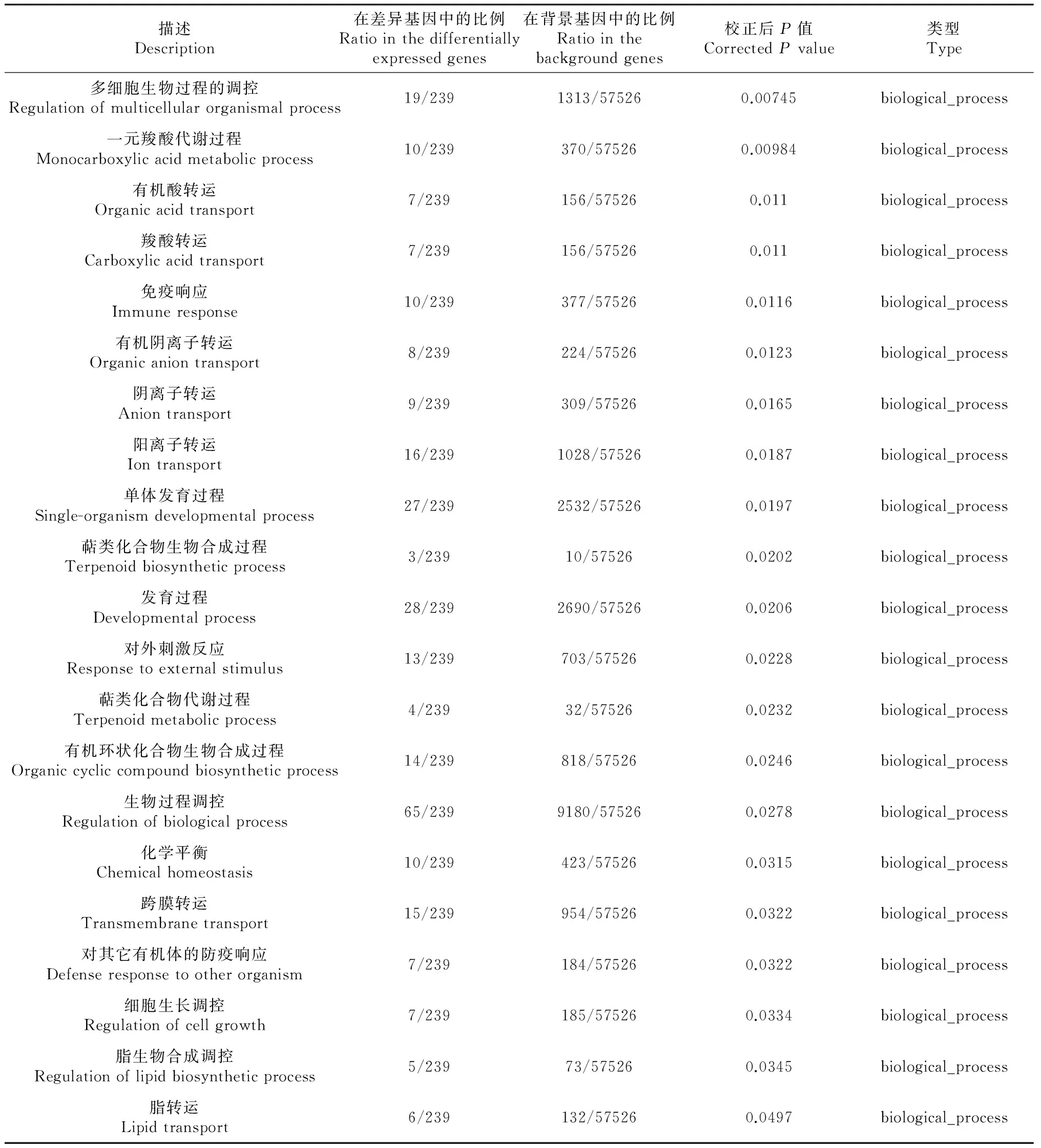
描述Description在差異基因中的比例Ratiointhedifferentiallyexpressedgenes在背景基因中的比例Ratiointhebackgroundgenes校正后P值CorrectedPvalue類型Type多細胞生物過程的調控Regulationofmulticellularorganismalprocess19/2391313/575260.00745biological_process一元羧酸代謝過程Monocarboxylicacidmetabolicprocess10/239370/575260.00984biological_process有機酸轉運Organicacidtransport7/239156/575260.011biological_process羧酸轉運Carboxylicacidtransport7/239156/575260.011biological_process免疫響應Immuneresponse10/239377/575260.0116biological_process有機陰離子轉運Organicaniontransport8/239224/575260.0123biological_process陰離子轉運Aniontransport9/239309/575260.0165biological_process陽離子轉運Iontransport16/2391028/575260.0187biological_process單體發育過程Single-organismdevelopmentalprocess27/2392532/575260.0197biological_process萜類化合物生物合成過程Terpenoidbiosyntheticprocess3/23910/575260.0202biological_process發育過程Developmentalprocess28/2392690/575260.0206biological_process對外刺激反應Responsetoexternalstimulus13/239703/575260.0228biological_process萜類化合物代謝過程Terpenoidmetabolicprocess4/23932/575260.0232biological_process有機環狀化合物生物合成過程Organiccycliccompoundbiosyntheticprocess14/239818/575260.0246biological_process生物過程調控Regulationofbiologicalprocess65/2399180/575260.0278biological_process化學平衡Chemicalhomeostasis10/239423/575260.0315biological_process跨膜轉運Transmembranetransport15/239954/575260.0322biological_process對其它有機體的防疫響應Defenseresponsetootherorganism7/239184/575260.0322biological_process細胞生長調控Regulationofcellgrowth7/239185/575260.0334biological_process脂生物合成調控Regulationoflipidbiosyntheticprocess5/23973/575260.0345biological_process脂轉運Lipidtransport6/239132/575260.0497biological_process
2.4 差異表達基因的KEGG功能分類
對這些差異表達基因進行KEGG功能分類,發現有6個調節途徑發生了顯著性改變(表4)。其中,有3個途徑發生了極顯著改變,分別是Steroid biosynthesis,Terpenoid backbone biosynthesis和Ovarian steroidogenesis。
3 討 論
金堂黑山羊是一個優秀的地方山羊品種,具有生長發育快,體格大,特別是其繁殖力明顯高于其它山羊品種。山羊是季節性發情動物,分別于春季和秋季發情。只有在發情期,山羊才會啟動卵泡發育。已有研究表明,山羊發情后在一個發情期后出現3~4個卵泡波,第1個FSH卵泡波峰值出現在發情后的第1~5天。張超等[4]研究了不同繁殖力山羊血漿中FSH的分泌規律,發現大足黑山羊在1個發情周期出現3個卵泡波,第1個卵泡波FSH峰值出現在第2天,為8.422 IU/L,而莎能奶山羊在1個發情期有4個卵泡波,第1個卵泡波FSH峰值出現在第1天,為6.227 IU/L。葛世豪等[5]研究了濟寧青山羊的FSH分泌規律,發現濟寧青山羊在1個發情有4個卵泡波,第1個卵泡波FSH峰值出現在第2
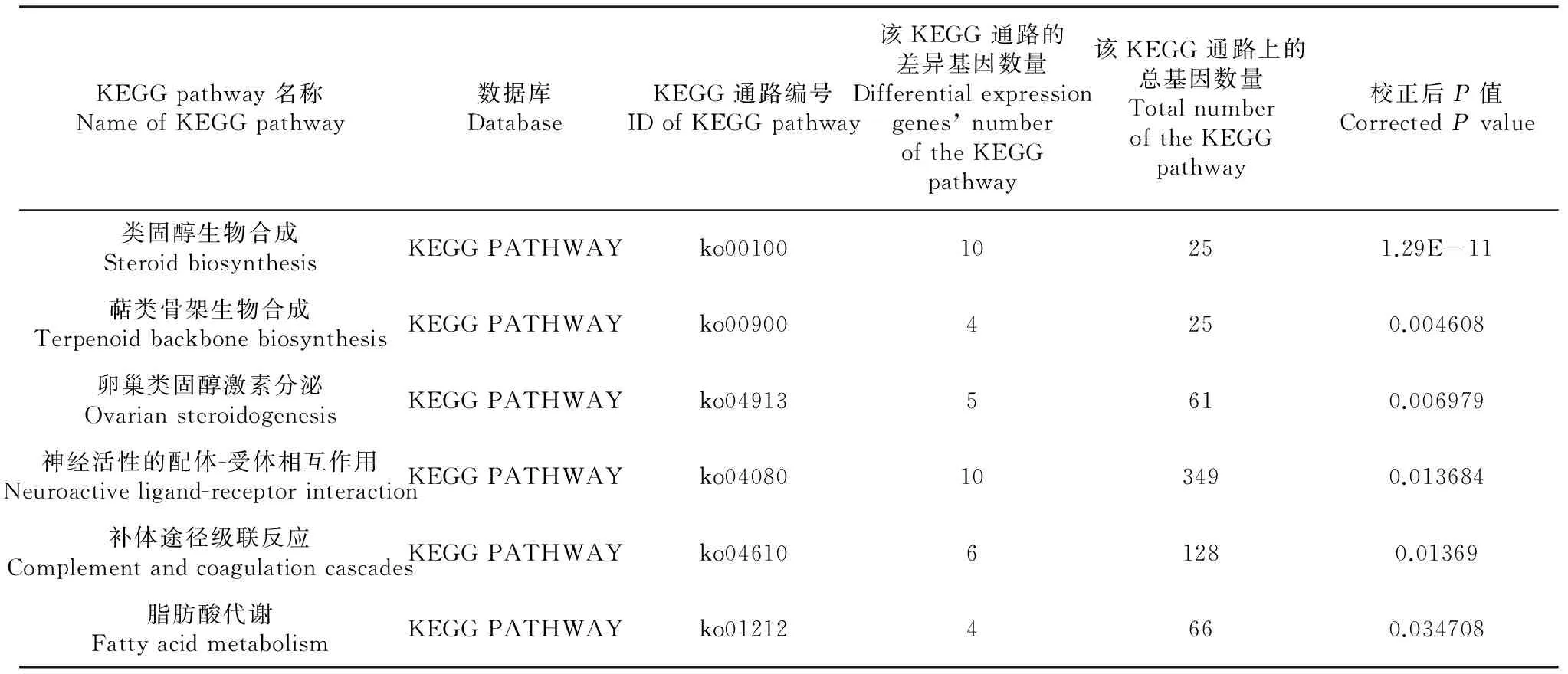
表4 金堂黑山羊卵巢差異表達基因的KEGG功能分類
天,為3.63 IU/L。侯衍猛等[6]研究了萊蕪黑山羊的FSH分泌規律,發現萊蕪黑山羊在1個發情有4個卵泡波,第1個卵泡波FSH峰值出現在第5天,為0.70 IU/L。綜上,可以確定,山羊在發情后第1~5天,將出現第一個卵泡波FSH峰,峰值為0.7~8.4 IU/L。所以本次研究中,在發情后的第2天對試驗組金堂黑山羊進行FSH注射,提高金堂黑山羊的FSH信號強度。按照寧波激素一廠的產品說明,山羊超數排卵為間隔12 h連續4 d肌注FSH,總量為200~250 IU,平均每次注射60~80 IU。為此選擇一次注射,注射劑量為100 IU,并在注射24 h后,摘取金堂黑山羊的卵巢,檢測FSH信號不同強度調節后,卵巢表達譜出現的差異。
FSH是調控動物發育、生長、成熟以及生殖相關的重要垂體激素,是動物下丘腦-垂體-性腺軸中的重要激素,下丘腦GnRH脈沖式釋放,刺激垂體分泌FSH、LH分泌,FSH、LH作用于雌性動物卵巢,刺激其分泌性激素。研究表明,動物卵細胞的發育依賴于FSH和LH的刺激,其中,FSH在原始卵母細胞的征集、類固醇激素的產生、LH受體的表達調節中起著至關重要的作用。在本次研究中,發現隨著FSH信號強度的增加,使金堂黑山羊的卵巢激素合成、分泌以及信號傳導的相關酶的表達量顯著增加,這就意味著,FSH信號傳導對卵巢性激素的釋放有極其顯著的正向調節作用[7],這與以上的結論完全吻合。
在本次研究中,發現FSH信號強度變化也顯著影響卵巢的脂代謝生物學過程。Mruk DD 等[8]發現,在精子生成的過程中,支持細胞對于精子的發生起著嚴格的限制性作用,不僅僅是提供能量和結構性支持,還提供精子細胞的發育和移動,還控制著成熟精子的釋放。在本次研究中發現的脂代謝生物學過程顯著變化也與以上結果類似,卵子成熟也包含著原始卵泡征集、發育、優勢化、排卵等重要生物學過程,在這個過程中,需要大量能量來供給這些生物學過程的進行,所以卵巢的脂代謝過程隨著FSH信號的增強顯著增加。
在本次研究中,發現FSH信號強度變化也顯著影響卵巢的配體受體反應。FSH的生理功能是通過分布于性腺的特異性FSH受體(follicle-stimulating hormone receptor, FSHR) 所介導。支持細胞和顆粒細胞分別是FSH作用于睪丸和卵巢的靶細胞。FSH與位于靶細胞膜上的促卵泡素受體(FSHR)結合,經過受體(FSHR)介導信息傳遞到靶細胞內,它們主要利用胞內第二信使cAMP作為信號,利用PKA途徑進行信號傳導[9-11]。為響應FSH信號強度,那么金堂黑山羊也通過加強卵巢的配體受體反應來提高信號強度,這與觀察到的現象是一致的。
4 結 論
綜上所述,FSH信號強度變化作用在卵巢上主要通過調節卵巢激素來調節排卵時卵巢的內環境,通過調節卵巢上FSH信號傳導通路來響應FSH信號強度的變化,通過調節脂代謝來做好卵巢排卵前的能量準備,從而達到響應FSH信號強度變化,改變動物的排卵數,從而調節了動物的繁殖性能。
[1]李 鍵. 肉山羊安全生產技術[M]. 北京:中國農業出版社,2005.
[2]Tang H, Wang X, Bowers J E, et al. Unraveling ancient hexaploidy through multiply-aligned angiosperm gene maps[J]. Genome Res, 2008, 18: 1944-1954.
[3]Xie C, Mao X, Huang J, et al. KOBAS 2.0: a web server for annotation and identification of enriched pathways and diseases[J]. Nucleic Acids Res 2011, 39: 316-322.
[4]張 超,羅艷梅,張家驊,等. 不同繁殖力山羊血漿中INHB、ACTA和FSH變化規律的研究[J]. 中國畜牧雜志,2011,47(7):24-27.
[5]葛仕豪,高立坤,侯衍猛,等. 濟寧青山羊發情周期內促性腺激素和性激素分泌規律的研究[J]. 西南農業學報,2007,20(6):1348-1352.
[6]侯衍猛,曹洪防,徐云華,等. 萊蕪黑山羊發情周期中FSH、LH、E2和P的分泌規律[J].中國獸醫學報,2006,26(3):340-343.
[7]張家驊. 家畜生殖內分泌學[M]. 北京:高等教育出版社,2007.
[8]Mruk D D, Cheng C Y. Sertoli-Sertoli and Sertoli-germ cell interactions and their significance in germ cell movement in the seminiferous epithelium during spermatogenesis[J]. Endocr Rev,2004,25: 747-806.
[9]Segaloff D L,Ascoli M. The lutropin/choriogonadotropin receptor 4 years later[J]. Endocrine Reviews,1993,14: 324-342.
[10]Christenson L K, Gunewardena S, Hong X, et al. Research resource: preovulatory LH surge effects on follicular theca and granulosa transcriptomes[J]. Molecular Endocrinology, 2013, 27: 1153-1171.
[11]Li Q, Jimenez-Krassel F, Ireland J J, et al. Gene expression profiling of bovine preovulatory follicles: gonadotropin surge and prostanoid-dependent up-regulation of genes potentially linked to the ovulatory process[J]. Reproduction, 2009, 137: 297-307.
(責任編輯 陳 虹)
EffectofFSHSignalRegulationonGeneExpressionProfilingofOvaryinJintangBlackGoat
CAO Ye1,2,ZHAO Su-jun1,2,XIE Jing1,2,WANG Qiu-shi1,2,LIAO Dang-jin1,2
(1.Sichuan Animal Science Academy, Sichuan Chengdu 610066, China; 2.Animal Breeding and Genetics Key Laboratory of Sichuan Province, Sichuan Chengdu 610066, China)
【Objective】The purpose of this study was to understand the model of FSH signal regulation mechanism in Jintang black goat.【Method】10 healthy adult Jintang black goats in the same reproductive cycle were randomly divided into two groups, one was as the control group, and the other was as the test group. On the second day of the estrus, the test group was injected with FSH 100IU, and the ovaries of Jintang black goats were obtained in 24 hours after injection; On the third days of the estrus, the ovaries of the control group were obtained, their total RNA were extracted, and the expression profiles of their total RNA were sequenced. 【Result】The 30 906 038, 27 763 272 effective bands in the control group and the test group were obtained, respectively; After statistical analysis, the normalized sequences were 30216946, 27124882 bands, and the coincidence rate of goat genome were 85.30 %, 86.13 %,respectively. By comparison, 239 genes were found to be significantly different, of which 139 genes were up-regulated, and 100 genes was down-regulated. GO functional classification reveals that there were significant differences in 86 biological processes, cellular components and molecular functions. KEGG function classification indicated that significant changes were found in 6 regulatory pathways.【Conclusion】FSH signal intensity changes had effect on the ovary, which caused mainly to regulate the secretion of ovarian hormones, the FSH signal transduction pathway and lipid metabolism and finally regulated the animal reproductive performance.
Jintang black goat; FSH signal; Ovary; Gene expression profiling
S826
A
1001-4829(2017)11-2595-08
10.16213/j.cnki.scjas.2017.11.035
2017-05-20
四川省應用基礎研究項目“金堂黑山羊FSH信號通路調節機制的研究”(2015JY0008)
曹 冶(1970-),男,博士,研究員,現從事動物分子遺傳研究,E-mail:cy3831@163.com。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
音樂探索(2022年2期)2022-05-30 21:01:37
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
