北美海篷子三萜皂苷bigelovii E抑制mTOR信號通路誘導乳腺癌MCF-7細胞凋亡
2017-12-02 08:26:08黃真真張丹玉管福琴王奇志
中國藥理學通報 2017年12期
關鍵詞:乳腺癌
黃真真,張丹玉,師 琪,管福琴,王奇志,王 鳴,馮 煦,單 宇
(江蘇省中國科學院植物研究所,江蘇省農業種質資源保護與利用平臺天然產物研究中心,江蘇 南京 210014)
乳腺癌是女性最常見的惡性腫瘤之一。據全國腫瘤登記中心預計,2015年中國乳腺癌發病率和死亡率分別占女性癌癥發病和死亡的15%和7%[1],并呈現逐年上升的趨勢[2]。臨床研究表明,一些天然產物在藥理學和癌癥治療中具有潛在的應用,具有直接性和毒性作用小等特點,可直接作用于病變部位,減少了對正常組織和細胞的損害。因此,從天然藥物中尋找抗腫瘤的新型藥物具有重要意義[3]。北美海篷子(SalicorniabigeloviiTorr.)生長于鹽堿地、鹽湖旁及海邊[4],可直接利用海水灌溉[5],栽培海蓬子還能吸收與利用海水水產養殖池排污物的營養,構建生態濕地,為野生禽類提供食物和棲息地,有益于改良生態環境[6]。現代藥理學研究表明,北美海篷子具有抗菌、抗炎、抗氧化、抗凝血的作用[7-8]。此外,研究發現,從北美海篷子分離得到多種皂類成分,都具有很好的抗腫瘤作用[9]。bigelovii E是從北美海篷子中分離得到的一種三萜類化合物,結構見Fig 1[10]。本課題組研究發現,bigelovii E對乳腺癌細胞MCF-7具有明顯的體外抑制作用,且對正常細胞的毒性較低,但其作用機制尚不明確。因此,本研究進一步觀察了bigelovii E對MCF-7細胞凋亡的影響,分析bigelovii E對MCF-7細胞線粒體凋亡通路的調控及其作用機制。

Fig 1 Chemical structure of bigelovii E
1 材料與方法
1.1材料
1.1.1細胞株 人類癌細胞系(MCF-7、Hela、HepG2、A549、B16、Lovo、LN229)和正常細胞株(HLF-1)均購自中國科學院上海細胞庫。
1.1.2試劑 Bigelovii E由本課題組從北美海篷子中分離純化獲得;胰蛋白酶和DMEM培養基購自以色列BioInd公司;胎牛血清(FBS)購自北京索萊寶科技有限公司;MTT、二甲基亞砜(DMSO)、碘化丙啶(PI)、Hoechst 33258、羅丹明123、DCFDA、NAC均購自美國Sigma Aldrich公司;Annexin V-PE/7-ADD細胞凋亡試劑盒,購自美國BD公司;BCA蛋白測定試劑盒、RIPA裂解緩沖液,購自江蘇碧云天生物技術研究所;一抗PARP、cleaved-caspase-3、cleaved-caspase-9、cleaved-caspase-7、Mcl-1、Bax、Bak、Bcl-xl、mTOR、p-mTOR、p-p70S6K、p-4-EBP、β-actin,Tubulin以及二抗,購自美國CST公司;ECL化學發光液購自美國Thermo Fisher Scientific公司。所有其他化學試劑均購自沈陽國藥化學試劑有限公司。
1.2方法
1.2.1細胞培養 細胞在含有10% FBS、青霉素100 kU·L-1、鏈霉素100 g·L-1的DMEM或RPMI 1640中培養,培養條件為37℃、5% CO2。培養液2~3 d更換1次。
1.2.2MTT法 分別取處于對數生長期的細胞,鋪于96孔培養板中,每孔100 μL,含有細胞5 000個,分別用(3.125、6.25、12.5、25、50、100 mg·L-1)的 bigelovii E處理,每個濃度重復3孔,在37℃、含5% CO2的培養箱中孵育。72 h后,向各孔中加10 μL MTT(5 g·L-1),培養箱中繼續孵育4 h后,每孔中加入100 μL DMSO,振搖5 min,用酶標儀在波長570 nm下測定每孔的吸光值,進行至少3次獨立實驗。通過確定每個細胞系的IC50值,檢測 bigelovii E對腫瘤細胞和正常細胞活力的影響。
1.2.3集落克隆實驗 將細胞以400個細胞/孔的密度接種在6孔板上。 24 h后,用 bigelovii E(2.5、5、10 mg·L-1)或DMSO處理MCF-7細胞。培養液每3 d更換1次。再培養10 d,觀察到細胞集落形成后,去除培養液,PBS洗2次,4%多聚甲醛固定10 min,結晶紫染色后計數。進行至少3次獨立實驗。
1.2.4Hoechst 33258熒光染色 取處于對數生長期的乳腺癌MCF-7細胞,常規消化,制成單細胞懸液,計數,以2.5×107·L-1接種于6孔板中,用不同濃度的 bigelovii E(2.5、5、10 mg·L-1)處理48 h后,細胞用PBS洗滌2次,固定液固定10 min,然后用0.5 mL Hoechst 33258染色5 min,去除培養液,PBS洗2次,在倒置熒光顯微鏡下分析形態學變化。
1.2.5Annexin V-PE/7-ADD雙染法測定細胞凋亡率 取對數生長期細胞,1×105個接種于6孔板,用bigelovii E(2.5、5、10 mg·L-1)處理細胞48 h,并用PBS洗滌。隨后收集細胞,并重懸于100 μL結合緩沖液中。將細胞懸浮液與Annexin V-PE和7-ADD染液在黑暗中孵育15 min,加入400 μL結合緩沖液,流式細胞儀檢測,采用Accuri C6 software獲取并分析數據。
1.2.6檢測線粒體膜電位(mitochondrial membrane potential,MMP) 將MCF-7細胞(1×105/孔)接種到6孔板,用bigelovii E(2.5、5、10 mg·L-1)處理。48 h后棄去培養基,用PBS清洗3次,然后在含有10 μmol·L-1RH123探針的無血清DMEM培養基中37℃孵育30 min。細胞用PBS洗滌3次,常規消化,制成單細胞懸液后,用流式細胞儀檢測熒光強度。
1.2.7細胞內活性氧簇(reactive oxygen species,ROS)水平的測量 將對數生長期細胞(1×105/孔)接種在6孔板中,然后用含有不同藥物的培養基處理。收集細胞后,加入10 μmol·L-1DCFDA,在黑暗中37℃孵育30 min。流式細胞儀測定細胞內ROS含量。
1.2.8Western blot 檢測相關蛋白的表達 細胞用不同濃度的 bigelovii E處理48 h后,收集細胞,提取各實驗組細胞總蛋白。等量的蛋白(30 μg)在10% SDS-聚丙烯酰胺凝膠上電泳,并轉移到PVDF膜,用 5% 脫脂奶粉室溫封閉2 h,然后加適當的一抗4℃孵育過夜,TBST洗滌3次,每次10 min,二抗室溫孵育2 h,ECL發光液反應1 min后,顯影,定影。
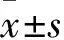
2 結果
2.1BigeloviiE對腫瘤細胞和正常細胞增殖的影響使用MTT法檢測 bigelovii E對人腫瘤細胞和正常細胞增殖的抑制作用。bigelovii E對人乳腺癌細胞MCF-7、宮頸癌細胞Hela、肺癌細胞HepG-2、肝癌細胞A549、黑素瘤細胞B16、結腸癌細胞Lovo、腦膠質瘤細胞LN229的IC50值分別為8.17、8.67、13.14、10.64、18.35、9.48、34.93 mg·L-1(Fig 2)。其中,bigelovii E對MCF-7細胞株最敏感,并具有劑量依賴效應,同時對人正常細胞株HLF-1的存活率無明顯影響(Fig 3),表明bigelovii E對MCF-7具有較好的選擇性。
2.2BigeloviiE抑制MCF-7的集落形成集落形成可以體現細胞獨立生存能力,常用于抗癌藥物敏感實驗、腫瘤放射生物學實驗。Fig 4結果顯示,與對照相比,bigelovii E明顯抑制MCF-7的集落形成,且濃度越高效果越明顯,顯示了劑量依賴效應。

Fig 2 IC50 of bigelovii E on different cancer cells

Fig 3 Inhibitory effects of bigelovii E on proliferation of MCF-7 and HLF-1 cells
*P<0.05,**P<0.01vscontrol

Fig 4 The inhibitory effects of bigelovii E on clongenic potential of MCF-7
2.3BigeloviiE誘導MCF-7細胞凋亡Hoechst 33258染色結果顯示(Fig 5),bigelovii E 2.5、5、10 mg·L-1處理后,隨著藥物濃度增加,細胞可以觀察到更明亮的熒光和典型的凋亡特征,如細胞生長減少、體積縮小、變形為圓形,而對照組呈圓形或橢圓形的正常形態。將bigelovii E按不同終濃度(2.5、5、10 mg·L-1)處理MCF-7細胞48 h后,Annexin V-PE/7-ADD 雙染結果表明(Fig 6),與對照組相比,用bigelovii E處理后MCF-7細胞凋亡率明顯增加。Bigelovii E以濃度依賴性方式誘導細胞凋亡。
Fig 7的Western blot結果表明,不同濃度bigelovii E(2.5、5、10 mg·L-1)處理MCF-7細胞48 h后,活化的PARP、caspase-3、caspase-7、caspase-9表達水平明顯增加,進一步證明bigelovii E促進了細胞凋亡。

Fig 5 Effect of bigelovii E on MCF-7 morphological change (×200)
A: Control; B: Bigelovii E 2.5 mg·L-1; C: Bigelovii E 5 mg·L-1; D: Bigelovii E 10 mg·L-1

Fig 6 Effects of bigelovii E on MCF-7 by Annexin V-PE and 7-ADD double staining
A: Control; B: Bigelovii E 2.5 mg·L-1; C: Bigelovii E 5 mg·L-1; D: Bigelovii E 10 mg·L-1
2.4BigeloviiE誘導MCF-7細胞線粒體功能紊亂為了檢測bigelovii E誘導的MCF-7細胞凋亡是否通過線粒體功能介導,通過RH123染色和流式細胞儀分析測定Δψm。Fig 8結果顯示,2.5 mg·L-1bigelovii E處理MCF-7細胞24 h后,線粒體膜電位出現明顯降低。隨著作用濃度升高至5、10 mg·L-1,流式細胞儀檢測到更明顯的去極化峰。這一結果證明,bigelovii E作用細胞后,引起線粒體膜電位下降,導致線粒體功能紊亂,進而激發線粒體凋亡,誘導細胞凋亡。

Fig 7 Effects of bigelovii E on expression of apoptosis perform protein in MCF-7

Fig 8 Effects of bigelovii E on integrity of mitochondrial membrane in MCF-7
A: Control; B: Bigelovii E 2.5 mg·L-1; C: Bigelovii E 5 mg·L-1; D: Bigelovii E 10 mg·L-1
ROS不平衡可能促進線粒體功能障礙,并引發線粒體介導的細胞凋亡。Fig 9結果表明,與對照相比,bigelovii E作用后,細胞內ROS水平增加,而這種增加效應在加入ROS阻斷劑NAC后有所回落。說明bigelovii E可引發細胞內ROS水平增加,使線粒體功能受損。
通過Western blot進一步檢測了 bigelovii E對Bcl-2家族的影響,發現抗凋亡蛋白Bcl-xL和Mcl-1表達明顯降低,而促細胞凋亡蛋白Bax表達明顯增加(Fig 10)。說明bigelovii E可通過調控線粒體凋亡通路相關蛋白,誘導MCF-7細胞凋亡。
2.5BigeloviiE調節mTOR通路蛋白的表達Fig 11的Western blot結果表明,mTOR及其下游靶點p70S6K和4-EBP磷酸化水平均明顯降低,但mTOR蛋白總量并未發生明顯變化。因此推測bigelovii E可通過調控mTOR的磷酸化位點,抑制mTOR活化,進而降低其下游靶點p70S6K和4-EBP磷酸化水平,激活線粒體凋亡通路,最終誘導細胞凋亡。

Fig 9 Effects of bigelovii E on intracellular ROS in MCF-7

Fig 10 Effects of bigelovii E on expression of Bcl-2 protein family in MCF-7

Fig 11 Effects of bigelovii E on expression of mTOR signaling pathway in MCF-7
3 討論
目前,乳腺癌已成為世界范圍內的嚴重公共衛生問題。我國雖是乳腺癌低發國家,但其發病率呈上升趨勢[3]。因此,為了減緩這一趨勢,從天然藥物中尋找新型有效的抗乳腺癌藥物具有重要意義。我們課題組研究發現,bigelovii E作用于MCF-7細胞,可調控線粒體凋亡通路,誘導細胞凋亡,具有良好的抗腫瘤效果。
我們的研究認為mTOR是bigelovii E抑制MCF-7細胞增殖的重要靶點。mTOR信號通路參與細胞凋亡過程,在多種疾病中發揮重要作用[11-12]。Yu等[13]實驗證明,mTOR/p70S6K通路通過調控caspase級聯體系和Bcl-2家族中抗凋亡因子的表達,在細胞凋亡的線粒體損傷途徑中發揮作用。本研究發現,經bigelovii E處理后的MCF-7細胞中,p-mTOR、p-p70S6K和p-4-EBP的表達量與對照組相比均明顯減少,并且具有濃度依賴效應,表明mTOR/p70S6K信號轉導通路在bigelovii E誘導MCF-7細胞凋亡作用中具有重要地位。因此,推測bigelovii E可由mTOR介導的線粒體凋亡途徑,提高細胞凋亡執行蛋白的活化水平,促使細胞凋亡。
隨著線粒體調控細胞凋亡的發現,線粒體誘導腫瘤細胞凋亡成為目前研究的熱點[14-15]。線粒體是多種促細胞凋亡信號轉導分子的靶點,同時也是細胞死亡通路的整合元件,細胞凋亡過程中許多重要事件的發生都與線粒體密切相關。流式細胞術檢測表明,bigelovii E作用細胞后,可明顯降低線粒體膜電位,增大膜通透性,促使ROS釋放,進而影響線粒體功能。同時,Western blot 實驗表明,bigelovii E處理后的MCF-7細胞抗凋亡蛋白Bcl-xl、Mcl-1的表達與對照組相比明顯減少,而促凋亡蛋白Bax的表達明顯增加,且濃度依賴效應明顯。因此,推斷bigelovii E可通過上調Bax表達,下調Mcl-1、Bcl-xl,激發ROS釋放、降低線粒體膜電位,進而導致線粒體功能紊亂,誘導MCF-7細胞凋亡。
本實驗初步研究表明,bigelovii E對人乳腺癌MCF-7細胞有明顯的抗腫瘤活性,可激活mTOR介導的線粒體凋亡通路,進而誘導MCF-7細胞凋亡。但其作用途徑尚有待深入研究,以更全面、系統地闡明bigelovii E對MCF-7增殖抑制的分子機制。
(致謝:本研究是在江蘇省中國科院植物研究所完成,衷心感謝各位老師和同學在實驗過程中的幫助與指導。)
[1] Chen W, Zheng R, Baade P D, et al. Cancer statistics in China, 2015[J].CACancerJClin, 2016,66(2):115-32.
[2] 邱宇凡,胡蘊慧,張 瑾.DNA 同源重組修復與乳腺癌的研究進展[J].中國藥理學通報,2016,32(7):910-4.
[2] Qiu Y F, Hu Y H, Zhang J. Research progress of relationship between DNA homologous recombination repair and breast cancer[J].ChinPharmacolBull,2016,32(7): 910-4.
[3] 李依敏,蘇思貞,朱 靜,等.乳腺癌靶向藥物治療研究進展[J]. 山東醫藥,2017,57(9): 107-9.
[3] Li Y M, Su S Z, Zhu J, et al. Research progress in targeted drug therapy for breast cancer[J].ShandongMedJ, 2017,57(9): 107-9.
[4] 呂忠進,Glenn E, Hodges R, et al. 全海水灌溉的作物—北美海蓬子(上) [J]. 世界農業,2001,262(2):14-6.
[4] Lyu Z J, Glenn E, Hodges R, et al. Salicornia bigelovii: Crop of whole sea-water irrigation(Ⅰ)[J].WorldAgric, 2001,262(2):14-6.
[5] 趙可夫,范 海.世界上可以用海水灌溉的鹽生植物資源[J]. 植物學通報,2000,17(3):282-8.
[5] Zhao K F, Fan H. Halophyte resources in the world that allow seawater irrigation[J].ChinBullBotany, 2000,17(3): 282-8.
[6] 魏成金,尹金來,徐陽春.不同氮磷水平對北美海蓮子生長和品質的影響[J]. 南京農業大學學報,2006,29(4):59-63.
[6] Wei C J, Yin J L, Xu Y C. Effect of different nitrogen and phosphorus level on growth and quality ofSalicorniabigeloviiTorr. [J].JNanjingAgricUniv, 2006,29(4):59-63.
[7] Zhang S, Wei M, Cao C, et al. Effect and mechanism ofSalicorniabigeloviiTorr. plant salt on blood pressure in SD rats[J].Food&Function, 2015,6(3): 920-6.
[8] Chung Y C, Chun H K, Yang J Y, et al. Tungtungmadic acid, a novel antioxidant, from Salicornia herbacea [J].ArchPharmRes, 2005,28(10): 1122-6.
[9] 管福琴,單 宇,趙友誼,等.海蓬子屬植物藥理活性及其作用機制研究進展[J]. 中國藥理學通報,2013,29(9) :1188-91.
[9] Guan F Q, Shan Y, Zhao Y Y, et al. Research development on pharmacological activites and mechanism of Salicornia plants[J].ChinPharmacolBull, 2013,29(9): 1188-91.
[10] Shan Y, Li H, Guan F, et al. Triterpenoids from the herbs ofSalicorniabigelovii[J] .Molecules, 2015,20(11): 20334-40.
[11] Hansel D E, Platt E, Orloff M, et al. Mammalian target of rapamycin (mTOR) regulates cellular proliferation and tumor growth in urothelial carcinoma [J].AmJPathol, 2010,176(6): 3062-72.
[12] Sun Y W, Huang W J, Hsiao C J, et al. Methoxychalcone induces cell-cycle arrest and apoptosis in human hormone-resistant prostate cancer cells through PI 3-kinase- independent inhibition of mTOR pathways[J] .Prostate, 2010,70(12): 1295-306.
[13] Yu C C, Wu P J, Hsu J L, et al. Ardisianone, a natural benzoquinone, efficiently induces apoptosis in human hormone-refractory prostate cancers through mitochondrial damage stress and survivin downregulation[J].Prostate, 2013,73(2): 133-45.
[14] Xiao J X, Huang G Q, Zhu C P, et al. Morphological study on apoptosis Hela cells induced by soyasaponins[J].ToxicolInVitro, 2007,21(5): 820-6.
[15] Hermisson M, Wagenknecht B, Wolburg H, et al. Sensitization to CD95 ligand-induced apoptosis in human glioma cells by hyperthermia involves enhanced cytochrome c release[J] .Oncogene, 2000,19(19): 2338-45.
猜你喜歡
中老年保健(2022年6期)2022-08-19 01:41:48
現代臨床醫學(2022年1期)2022-02-12 02:04:58
甘肅科技(2020年20期)2020-04-13 00:30:42
中國生殖健康(2019年2期)2019-08-23 08:11:42
中國生殖健康(2019年6期)2019-01-06 09:20:12
中國生殖健康(2019年5期)2019-01-06 09:16:40
幸福家庭(2019年14期)2019-01-06 09:15:38
祝您健康(2018年5期)2018-05-16 17:10:16
癌癥進展(2016年9期)2016-08-22 11:33:20
中國組織化學與細胞化學雜志(2016年4期)2016-02-27 11:16:08
