百里醌對阪崎克羅諾腸桿菌的抑制作用
2018-01-08 02:47:11孫慧慧張文婷陳怡飛夏效東
食品科學 2018年1期
石 超,孫慧慧,孫 正,郭 都,張文婷,孫 怡,陳怡飛,夏效東*
(西北農林科技大學食品科學與工程學院,陜西 楊凌 712100)
百里醌對阪崎克羅諾腸桿菌的抑制作用
石 超,孫慧慧,孫 正,郭 都,張文婷,孫 怡,陳怡飛,夏效東*
(西北農林科技大學食品科學與工程學院,陜西 楊凌 712100)
為探究百里醌對阪崎克羅諾腸桿菌的抑制作用及可能的抑制機理,檢測了百里醌對阪崎克羅諾腸桿菌的最小抑菌濃度(minimal inhibitory concentrations,MIC)及對其生長動力學模型的影響,并測定百里醌處理后阪崎腸桿菌胞內ATP濃度、胞內pH值、膜電位、細胞膜完整性的變化,最后利用場發射掃描電子顯微鏡觀察細胞形態的改變。結果表明:百里醌對阪崎克羅諾腸桿菌的MIC為0.3~0.6 mg/mL;百里醌使阪崎克羅諾腸桿菌的生長遲滯期延長,最大生長速率減小;百里醌影響了阪崎克羅諾腸桿菌細胞膜的通透性,表現為:質量濃度為MIC和2×MIC的百里醌使細胞內ATP濃度由6.52 μmol/L分別降低為0.27 μmol/L和0.17 μmol/L,胞內pH值分別由5.69降低為5.22和4.99,細胞膜完整菌體比例分別降低至80%和22%,引起細胞膜膜電位超極化;場發射掃描電子顯微鏡觀測表明百里醌使阪崎克羅諾腸桿菌細胞膜褶皺,菌體干癟。綜上所述,百里醌是通過影響細胞膜通透性和改變細胞形態抑制阪崎克羅諾腸桿菌。這些結果表明百里醌有潛力作為控制阪崎克羅諾腸桿菌的天然抑菌劑應用于食品中。
阪崎克羅諾腸桿菌;百里醌;細胞膜;膜電位
阪崎克羅諾腸桿菌(Cronobacter sakazakii)是一種革蘭氏陰性、桿狀、無芽孢、兼性厭氧、周生鞭毛的食源性條件致病菌[1],它能造成新生兒腦膜炎、菌血癥、壞死性小腸結腸炎等多種疾病,致死率高達50%以上[2]。免疫力低下的成年人(主要是老人)食用被阪崎克羅諾腸桿菌污染的食物也能導致骨髓炎、膿毒癥、腳潰瘍等多種疾病的發生。目前在臨床治療中多使用抗生素治療阪崎克羅諾腸桿菌感染,雖可以使患者康復,但是常常伴隨有嚴重的神經系統后遺癥和發育障礙等癥狀[3]。鑒于阪崎克羅諾腸桿菌的巨大潛在危害,2 0 0 2年食品微生物標準委員會(I n t e r n a t i o n a l Commissionon Microbiological Specifications for Foods,ICMSF)把阪崎腸桿菌描述為“對特定人群造成嚴重危害,危及生命或引起長期慢性實質性后遺癥的細菌”[4]。
預防和控制食源性致病菌對于各國的公共健康衛生具有重要的意義。為抑制和殺滅食品中的病原微生物,食品加工生產過程中通常以添加天然合成防腐劑來保證食品安全、延長食品貨架期。然而由于合成的物質往往具有潛在的毒性,近年來部分研究者將目光集中到植物來源,尤其是水果、蔬菜和中草藥等的活性物質上[5]。消費者在消費過程中也傾向于選擇“零添加”或低含量防腐劑的產品[6]。植物源活性物質以其消費者接受度高及來源范圍廣被認為是合成防腐劑的最佳替代品[7]。
百里醌(thymoquinone,TQ),是黑種草(Nigella sativa)中的主要成分,占黑種草油含量的28%~57%[8]。黑種草,又稱為黑小茴香,是一種毛莨科一年生草本芳香植物[9],被用作香料、食品添加劑在食品生產中廣為應用,同時它也被作為藥物用來治療哮喘、腹瀉、頭痛、鼻塞、背部疼痛、高血壓和胃腸道問題[9-10],目前黑種草已被美國食品藥品監督管理局確認為公認安全的食品成分(GRAS 182.10)。研究表明,百里醌具有多種生物活性作用,包括抗腫瘤、抗炎、抑菌、抗氧化等[8,11-12]。
盡管百里醌已被報道對多種微生物有抑制效果,但其對阪崎克羅諾腸桿菌的抑制作用及機理鮮有研究。鑒于此,本實驗旨在探討百里醌對阪崎克羅諾腸桿菌的抑菌效果,并通過檢測其對細菌生長動力學模型、胞內ATP、胞內pH值、膜電位、細胞膜完整性及細胞形態的影響探討其可能的抑菌機理。
1 材料與方法
1.1 菌株與試劑
阪崎克羅諾腸桿菌(Cronobacter sakazakii)ATCC 29544、ATCC 29004、ATCC 12868和ATCC BAA-894購于美國模式菌株收集中心;阪崎克羅諾腸桿菌分離菌株12-2、14-15、18-7、18-8和18-13由西北農林科技大學食品科學與工程學院食品微生物研究團隊分離自市售嬰幼兒乳粉及米粉。
百里醌(色譜級,純度≥9 8%)、熒光探針DiBAC4(3)及cFDA-SE 美國Sigma公司;胰蛋白胨大豆瓊脂(tryptone soya agar,TSA)、胰蛋白胨大豆肉湯(tryptone soya broth,TSB)培養基 北京陸橋技術有限公司;ATP檢測試劑盒 上海碧云天生物技術公司;LIVE/DEAD?BacLightTM細菌活性檢測試劑盒 美國賽默飛世爾科技公司;其他所用有機溶劑均為國產分析純。
1.2 儀器與設備
InfiniteTMM200 PRO多功能酶標儀 瑞士Tecan公司;SmartSpecTMPlus分光光度計 美國Bio-Rad公司;5804R低溫冷凍離心機 德國Eppendorf公司;S-4800場發射掃描電子顯微鏡 日本Hitachi公司;微生物全自動生長曲線分析儀 芬蘭Bioscreen公司;SCIENTZ-IID超聲波裂解儀 寧波新芝生物科技股份有限公司。
1.3 方法
1.3.1 菌種活化
將凍存于-80 ℃的阪崎克羅諾腸桿菌采用劃線法在TSA平板上活化,挑取單菌落接種于30 mL TSB培養基中,將其置于37 ℃培養18 h使細菌處于對數生長后期,培養后的菌液經5 000×g離心15 min,去除上清液,用磷酸鹽緩沖液(phosphate buffered saline,PBS)洗滌菌體沉淀,反復洗滌3 次后,用一定量的PBS懸浮菌體沉淀,測定并調整菌懸液的OD600nm為0.5,使菌液濃度約為108CFU/mL。
1.3.2 最小抑菌濃度測定
最小抑菌濃度(minimal inhibitory concentrations,MIC)的測定采用瓊脂稀釋法[13]。將冷卻至45 ℃的TSA培養基加入24 孔培養板中,向其中加入百里醌并進行稀釋,使百里醌質量濃度為0.80、0.60、0.40、0.30、0.20、0.10、0.05 mg/mL,充分吹打混勻,培養基在室溫下放置至凝固。吸取2 μL菌懸液(1.3.1節方法制備)接種至24 孔板各孔中央。將24 孔板放置于37 ℃培養箱中培養24 h后觀測結果,其中可抑制阪崎克羅諾腸桿菌出現增長的最低質量濃度即為百里醌的MIC。實驗以不含有百里醌的培養基作為陰性對照,含有1 mg/mL氨芐西林的培養基為陽性對照。
1.3.3 百里醌對阪崎克羅諾腸桿菌生長動力學模型影響的測定
按照1.3.1節中的方法制備ATCC 29544菌懸液,調整菌懸液OD600nm為0.5并稀釋100 倍,使菌懸液濃度約為106CFU/mL。向96 孔培養板中每孔加入125 μL的菌懸液和125 μL不同質量濃度的百里醌溶液(TSB培養基配制),使百里醌的最終質量濃度為2×MIC、MIC、1/2×MIC、1/4×MIC、1/8×MIC、1/16×MIC和1/32×MIC,背景空白對照組添加250 μL TSB培養基,樣品對照組添加125 μL菌懸液及125 μL TSB培養基。設置微生物全自動生長曲線分析儀培養溫度為37 ℃并每隔1 h檢測600 nm波長處的OD值。
以培養時間為橫坐標,菌懸液OD600nm值為縱坐標,繪制生長曲線。選取修正Gompertz模型(式(1~3))擬合其生長狀況,并表征菌株的生長參數。利用SPSS軟件中的非線性回歸方法擬合A、B、μ、λ、M和μmax等參數值,以相關系數R2作為評估因子模型的擬合度。
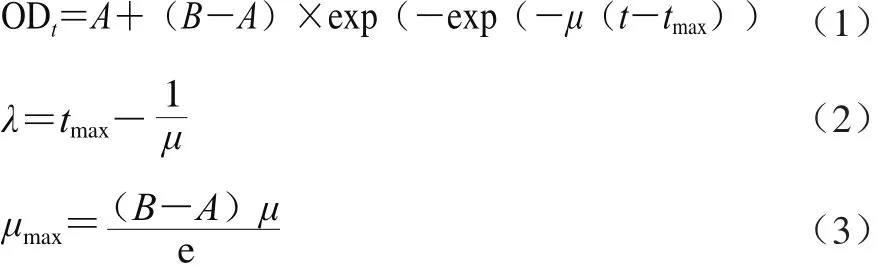
式中:ODt為在t時菌液的OD600nm值;t為培養時間/h;B為最大菌液濃度時的OD600nm值;A為初始菌液濃度時的OD600nm值;tmax為菌株達到指數期所用時間/h;μ為指數期的相對生長速率/h-1;λ為遲滯期/h;μmax為最大生長速率/h-1。
1.3.4 百里醌對阪崎克羅諾腸桿菌胞內ATP的影響
按照1.3.1節方法制備菌懸液,用PBS調整菌懸液的OD600nm為0.5。樣品實驗組中添加菌懸液及百里醌溶液(0、MIC和2×MIC),背景空白對照組添加PBS及百里醌溶液,將樣品置于37 ℃培養箱中培養30 min。隨后裂解超聲,具體方法如下:將樣品冰浴,逐個超聲,每超聲3 s停頓5~7 s,一輪超聲10 次,超聲4 輪;超聲時,探頭置于液面下約1 cm處,同時與離心管底部保持一定距離。每個樣品超聲結束后立即將離心管于100 ℃溫度下處理2~3 min使樣品中ATP酶滅活。將樣品離心取上清液,根據ATP檢測試劑盒說明書進行實驗,使用多功能酶標儀測定樣品上清液的熒光強度。同時,使用ATP檢測裂解液將ATP標準溶液稀釋成0.01、0.10、1.00、10.00 μmol/L的4 個濃度梯度,繪制熒光強度與ATP濃度間的標準曲線,并根據標準曲線計算樣品中ATP的濃度。
1.3.5 百里醌對阪崎克羅諾腸桿菌腸桿菌胞內pH值的影響
菌液制備同1.3.1節,將培養后的菌液4 ℃、5 000×g離心5 min,去除上清液,并用磷酸鉀緩沖液洗滌一次,調整菌懸液的OD600nm為0.5。用4-羥乙基哌嗪乙磺酸緩沖液洗滌菌體2 次,加入磷酸鉀緩沖液重懸浮,加入cFDA-SE熒光探針使其終濃度為3 μmol/L(37 ℃培養箱中孵育20 min),隨后用磷酸鉀緩沖液洗滌一次,重新懸浮,加入葡萄糖使其終濃度為10 μmol/L(37 ℃培養箱中孵育30 min),最后用磷酸鉀緩沖液洗滌菌體2 次并重懸浮菌體。實驗使用磷酸鉀緩沖液配制百里醌溶液,實驗組將百里醌溶液添加至菌懸液中,使百里醌質量濃度分別為0、MIC和2×MIC,背景空白對照組為磷酸鉀緩沖液配制的百里醌溶液,樣品于37 ℃培養箱中培養20 min。
為構建標準曲線,使用50 mmol/L甘氨酸、50 mmol/L氯化鉀、50 mmol/L檸檬酸、50 mmol/L磷酸二氫鈉溶液配制pH值為3、4、5、6、7、8、9、10的緩沖液,用氫氧化鈉、鹽酸溶液調節pH值。標準曲線組每孔加入200 μL上述緩沖液、30 μL菌液、10 μL尼日利亞菌素(10 μmol/L)和10 μL纈氨霉素(10 μmol/L)。使用多功能酶標儀測定發射波長520 nm,激發波長分別為490、440 nm時的熒光強度,繪制490 nm和440 nm時熒光強度的比值與pH值間的線性關系,并根據標準曲線計算樣品組胞內pH值。
1.3.6 百里醌對阪崎克羅諾腸桿菌膜電位的影響
細菌膜電位的測定參照Sanchez等[14]的方法,菌液制備同1.3.1節,以PBS洗滌菌體2 次并調整菌懸液OD600nm為0.5。將125 μL菌懸液加入黑色96 孔酶標板中,置于37 ℃培養箱中培養30 min。隨后向每孔中加入細胞膜電位熒光探針DiBAC4(3)(1 μmol/L),并再次37 ℃孵育30 min。然后將125 μL百里醌溶液加入黑色酶標板各孔中,使百里醌的質量濃度分別為0、MIC和2×MIC。百里醌作用5 min后使用多功能酶標儀檢測熒光強度,激發、發射波長分別為492、515 nm。激發和發射縫隙寬度分別為3、5 nm。通過計算百里醌作用組與對照組的相對熒光強度(處理組檢測所得熒光強度減去對照組檢測所得熒光強度),分析細菌膜電位的變化。若細胞內熒光強度增加,即膜電位增加,表示細胞去極化;反之,細胞內熒光強度降低,即膜電位降低,表示細胞超極化[24]。
1.3.7 百里醌對阪崎克羅諾腸桿菌細胞膜完整性的影響
參照Shi Chao等[15]的方法,使用LIVE/DEAD?BacLightTM細菌活性檢測試劑盒檢測百里醌對阪崎克羅諾腸桿菌細胞膜完整性的影響。具體方法如下:菌液制備同1.3.1節,將菌液4 ℃、10 000×g離心15 min,使用0.85% NaCl溶液洗滌菌體2 次,隨后加入2 mL 0.85% NaCl溶液。將重懸菌懸液分別取1 mL加入到20 mL 0.85% NaCl溶液(活菌組)和70%異丙醇溶液(死菌組)中,于25 ℃培養箱中培養1 h。使用0.85% NaCl溶液洗脫2~3 次后將兩組樣品調整至OD600nm為0.5(兩組誤差不超過0.01)。添加百里醌于0.85% NaCl溶液處理的菌液中,使百里醌質量濃度為0、MIC和2×MIC,并在37 ℃孵育30 min后,離心去掉上清液,加入相同體積的0.85% NaCl溶液重懸。
以不同體積活菌組菌懸液和死菌組菌懸液配制不同活菌比例(0%、10%、50%、90%、100%)的菌懸液,在黑色96 孔酶標板中設置樣品空白對照孔(0.85% NaCl溶液)、樣品孔和標準曲線孔,并向所有孔內加入相同體積2×SYTO/PI染料,充分吹打混勻。將96 孔酶標板置于25 ℃條件下培養5~10 min。使用多功能酶標儀檢測熒光強度,SYTO染料的激發及發射波長分別為485、542 nm,PI染料的激發及發射波長分別為485、610 nm。
1.3.8 百里醌對阪崎克羅諾腸桿菌細胞形態的影響
場發射掃描電子顯微鏡對阪崎克羅諾腸桿菌細胞形態的觀測參照Li Guanghui等[16]的方法,具體如下:按1.3.1節方法制備菌懸液,將添加不同質量濃度百里醌(0、MIC和2×MIC)的菌液(OD600nm為0.5)置于37 ℃培養箱中培養,分別在2 h和4 h取出樣品。隨后,使用PBS洗滌菌體2 次,并將菌體重新懸浮于2.5%戊二醛-PBS中固定(4 ℃)。12 h后依次使用PBS、無菌水洗滌菌體,并將菌體置于1%鋨酸中固定5 h,隨后使用不同體積分數(30%、50%、70%、80%、90%和100%)的乙醇梯度脫水,每次10 min。將樣品滴加至專用蓋玻片并貼附于場發射掃描電子顯微鏡載物臺,樣品抽真空脫水2 h后鍍金,使用場發射掃描電子顯微鏡觀測細菌細胞形態。
1.4 數據統計分析
2 結果與分析
2.1 百里醌對阪崎克羅諾腸桿菌的MIC
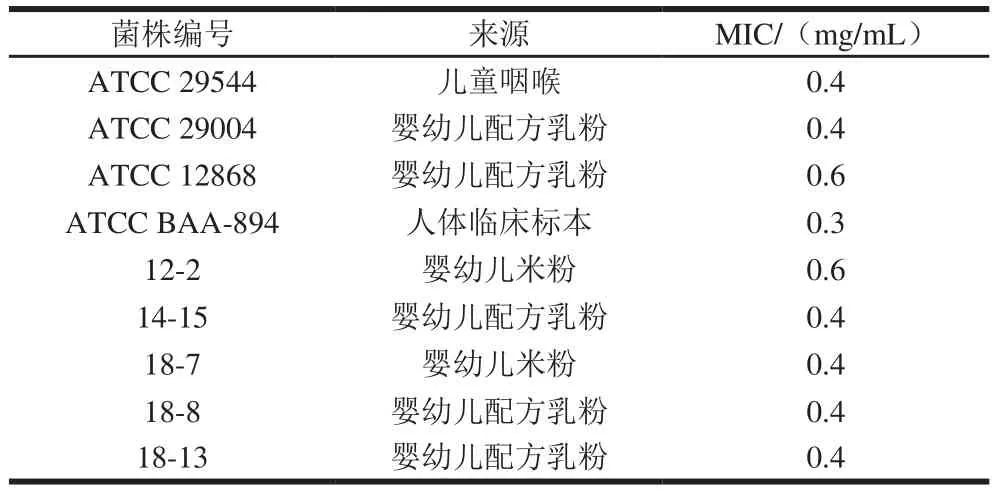
表1 百里醌對阪崎克羅諾腸桿菌的MICTable 1 Minimum inhibitory concentrations of thymoquinone against C. sakazakii
百里醌對9 株食品或臨床來源的阪崎克羅諾腸桿菌有良好的抑制效果。由表1可知,百里醌對阪崎克羅諾腸桿菌ATCC 29544、ATCC 29004的MIC為0.4 mg/mL,對4 種食品來源的分離菌(14-15、18-7、18-8、18-13)的MIC也為0.4 mg/mL。百里醌對ATCC 12868及分離菌12-2的MIC為0.6 mg/mL,對ATCC BAA-894的MIC為0.3 mg/mL。為了進一步探討百里醌對阪崎克羅諾腸桿菌的抑菌機理,選擇ATCC 29544進行后續的實驗。國內外一些研究也報道了植物源抑菌劑對阪崎克羅諾腸桿菌有抑制效果,如香芹酚和百里酚[17]、辛酸、檸檬酸、香草醛[18]、反式-肉桂醛[18]、藍莓花青素和野生藍莓汁[19],對比其他的植物源抑菌劑,百里醌對阪崎克羅諾腸桿菌具有較強的抑菌效果。
2.2 百里醌對阪崎克羅諾腸桿菌生長動力學模型的影響
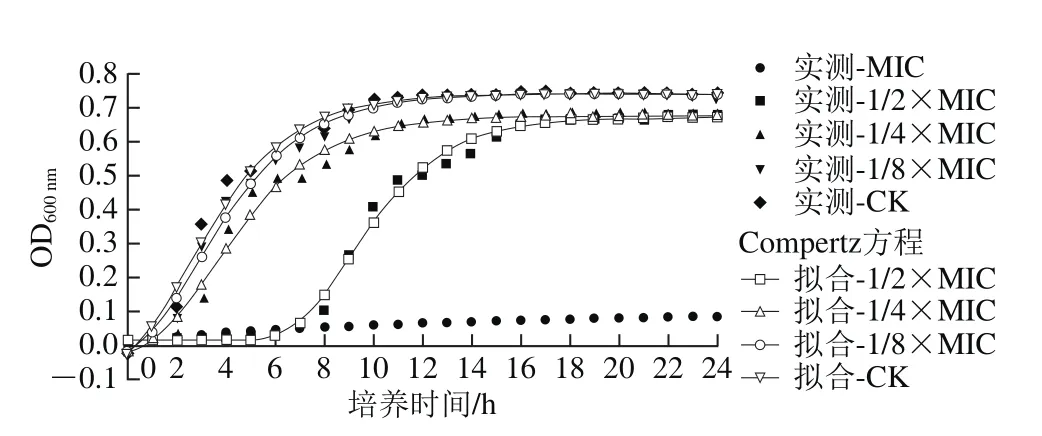
圖1 百里醌對阪崎克羅諾腸桿菌ATCC 29544生長曲線的影響Fig. 1 Growth curves of C. sakazakii ATCC 29544 cultured in TSB with various concentrations of thymoquinone
阪崎克羅諾腸桿菌在含有不同質量濃度百里醌的TSB中的生長曲線如圖1所示,應用修正Gompertz方程擬合ATCC 29544的生長曲線,各擬合方程的R2均大于0.98,表明此方程能夠較好地擬合菌株的生長狀況。由圖1可以看出,在TSB培養液中,質量濃度為MIC的百里醌幾乎完全抑制阪崎克羅諾腸桿菌的生長。當百里醌質量濃度為1/2×MIC時,阪崎克羅諾腸桿菌在6 h內幾乎沒有生長,生長17 h達到細菌數量的最大值。
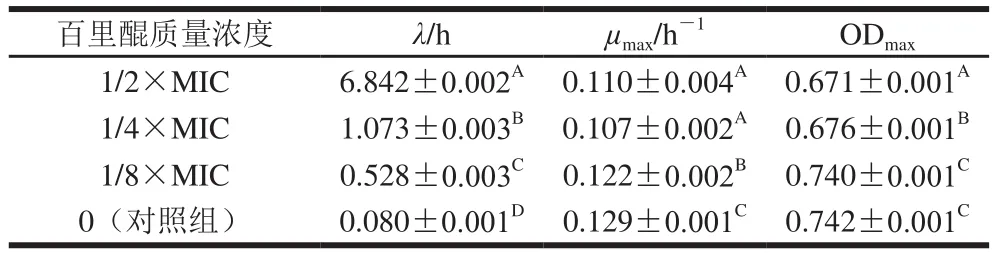
表2 阪崎克羅諾腸桿菌在含有不同質量濃度百里醌的TSB中的生長動力學參數(n=6)Table 2 Kinetic parameters of C. sakazakii cells during growth in TSB with different concentrations of thymoquinone (n= 6)
菌株的生長參數如表2所示,百里醌使阪崎克羅諾腸桿菌在TSB中生長遲滯期增長,最大生長速率減小,并呈現濃度依賴效應。
2.3 百里醌對阪崎克羅諾腸桿菌胞內ATP濃度的影響
螢火蟲熒光素酶催化熒光素產生熒光時需要ATP提供能量,并當螢火蟲熒光素酶和熒光素都過量時,在一定的濃度范圍內熒光的產生和ATP的濃度成正比,據此首先構建標準曲線,得到ATP濃度與相對熒光強度之間呈現良好的線性關系(y=21 365x+2 381;R2=0.999)。
由圖2可知,百里醌對阪崎克羅諾腸桿菌胞內ATP含量有極顯著的降低作用(P<0.01),并呈現濃度依賴效應。未經百里醌處理的對照組胞內ATP濃度為6.52 μmol/L,經質量濃度為MIC的百里醌處理的實驗組胞內ATP濃度降為0.27 μmol/L,經質量濃度為2×MIC的百里醌處理的實驗組降為0.17 μmol/L。ATP是細胞內能量代謝、酶反應和信號調節的基礎,對細胞生長、分裂和存活等細胞功能有著關鍵作用[20]。本實驗中百里醌使阪崎腸桿菌胞內ATP顯著降低,從而影響細菌存活。Li Guanghui等[16]也有類似的發現,綠原酸處理金黃色葡萄球菌細胞使細菌胞內ATP濃度降低。胞內ATP濃度的降低可以歸因于細菌自身水解速率增加[14],以及細胞膜通透性的損傷,使胞內ATP流出[20]。
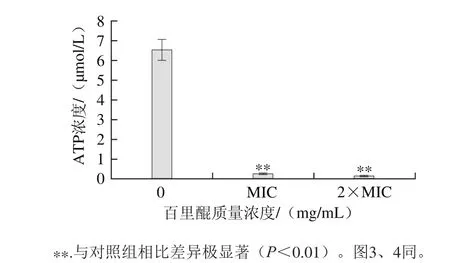
圖2 百里醌對阪崎克羅諾腸桿菌ATCC 29544胞內ATP濃度的影響Fig. 2 Effect of thymoquinone on intracellular ATP production in C. sakazakii ATCC 29544
2.4 百里醌對阪崎克羅諾腸桿菌胞內pH值的影響
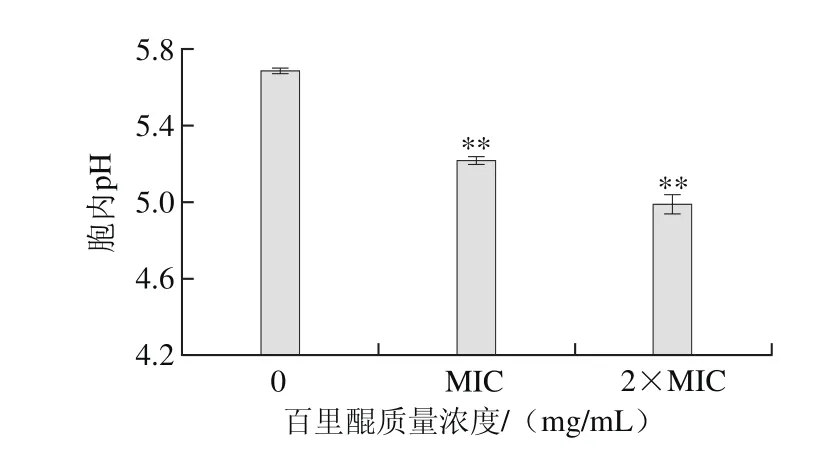
圖3 百里醌對阪崎克羅諾腸桿菌ATCC 29544胞內pH值的影響Fig. 3 Effect of thymoquinone on intracellular pH of C. sakazakii ATCC 29544
由圖3可知,百里醌對阪崎克羅諾腸桿菌胞內pH值有極顯著的影響(P<0.01)。未經百里醌處理的阪崎腸桿菌胞內pH值為5.69±0.01,經質量濃度為MIC的百里醌處理后,阪崎腸桿菌胞內pH值降低為5.22±0.02,經質量濃度為2×MIC的百里醌處理后,阪崎腸桿菌胞內pH值降低為4.99±0.05。
細菌pH值對于細胞內的DNA的轉錄與合成、酶活性及蛋白質合成等有非常重要的影響[21]。Turgis等[22]研究表明了芥末精油使大腸桿菌O157:H7胞內pH值從6.23降低為5.20,使鼠傷寒沙門氏菌胞內pH值從6.59降至5.44,胞內pH值的改變意味著細胞膜的損傷[14]。
2.5 百里醌對阪崎克羅諾腸桿菌細胞膜電位的影響

圖4 百里醌對阪崎克羅諾腸桿菌ATCC 29544細胞膜電位的影響Fig. 4 Effect of thymoquinone on membrane potentials of C. sakazakii ATCC 29544
由圖4可知,經百里醌處理的阪崎克羅諾腸桿菌細胞膜出現超級化(負相對熒光強度),并且經質量濃度為2×MIC百里醌處理比質量濃度MIC的處理后超級化現象更加明顯。靜息膜電位是活細胞的重要參數之一,與抑菌物質殺滅細菌作用密切相關[23]。實驗所使用的陰離子熒光染料DiBAC4(3)是一種細胞膜電位敏感的親脂性陰離子熒光染料,可以指示細胞膜電位的變化[18]。DiBAC4(3)本身無熒光,當進人細胞與胞漿內的蛋白質結合后才發出熒光,DiBAC4(3)進入細胞,細胞內熒光強度增加,即膜電位增加表示細胞去極化;反之,細胞內熒光強度降低即膜電位降低表示細胞超極化[24]。
通過實驗可以得出,百里醌引起阪崎腸桿菌細胞膜超極化,這與紫蘇、白蒿和金合歡提取物引起細菌細胞膜的超極化現象一致[14]。研究表明,出現超極化和去極化現象的原因為:pH值的改變及抑菌物質影響細菌細胞膜上離子的運動,特別是K+,從而影響胞體的自動調節[25]。
2.6 百里醌對阪崎克羅諾腸桿菌細胞膜完整性的影響

表3 不同質量濃度百里醌處理對阪崎克羅諾腸桿菌ATCC 29544細胞膜熒光強度的影響Table 3 Fluorescence of C. sakazakii ATCC 29544 treated with various concentrations of thymoquinone
綠色熒光強度與細胞膜完整細胞百分比呈現良好的線性關系,線性擬合方程為y=32 769x+10 632(R2=0.99)。由表3可知,與對照組相比,經百里醌處理的阪崎克羅諾腸桿菌細胞膜完整性極顯著下降(P<0.01)。未經百里醌處理的對照組細胞膜完整菌體比例為96%,僅經過質量濃度為MIC的百里醌處理15 min后,細胞膜完整菌體比例下降至80%。經質量濃度為2×MIC百里醌處理后,細胞膜完整的細菌比例僅為22%。Campos等[26]也得到相似結論,添加不同濃度的沒食子酸能夠顯著降低希氏乳桿菌的細胞膜完整性。Cherrat等[27]證明最小抑菌濃度的月桂精油能夠在細菌適宜生長的溫度下損傷大腸桿菌及單增李斯特菌細胞膜。
2.7 百里醌對阪崎克羅諾腸桿菌細胞形態的影響
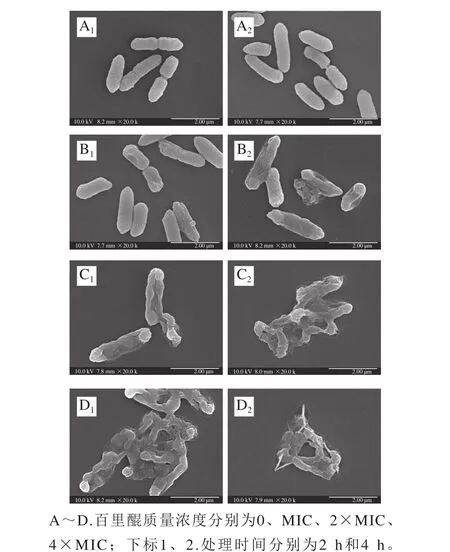
圖5 場發射掃描電子顯微鏡觀測百里醌對阪崎克羅諾腸桿菌ATCC 29544細胞形態的影響(20 000×)Fig. 5 Field emission scanning electron electron microscopic observation of the effect of thymoquinone on morphology of C. sakazakii ATCC 29544 (20 000 ×)
由圖5可知,未經百里醌處理和經由百里醌處理的細菌細胞形態存在明顯差異。未經百里醌處理的阪崎克羅諾腸桿菌表面光滑、飽滿,具有典型的革蘭氏陰性桿菌的形態,并且細胞能夠生長和分裂(圖5A1、A2)。但經過百里醌處理后,阪崎腸桿菌表面褶皺,菌體干癟。經4×MIC的百里醌處理后,阪崎腸桿菌菌體出現大面積塌陷,喪失了其固有形態(圖5D1、D2)。阪崎克羅諾腸桿菌細胞的受損程度和受損的比例隨著百里醌質量濃度的增加而升高。
植物源活性物質對其他多種病原菌細胞形態的改變也有類似報道。di Pasqua等[28]發現丁子香酚能夠破壞大腸桿菌O157:H7的細胞膜,導致細胞內容物流出。然而其他天然物質(香芹酚、肉桂醛、檸檬烯和百里酚)僅能導致外膜形態的改變。Rhayour等[29]發現丁香精油對于不同病原菌細胞形態的作用不同。作用大腸桿菌后,細胞膜表面出現孔洞,但對于枯草芽孢桿菌,菌體僅出現變形。在本實驗中,百里醌使菌體細胞表面塌陷但并沒有造成細胞膜破裂、瓦解。這說明百里醌是通過與細胞膜表面特定位點結合然后滲透到靶位點并影響其目標分子[30]。
3 結 論
本研究以百里醌為研究對象,以阪崎克羅諾腸桿菌為作用主體,探究百里醌對阪崎克羅諾腸桿菌的抑菌作用及可能的抑菌機理。利用瓊脂稀釋法測定了百里醌對9 株阪崎克羅諾腸桿菌的MIC為0.3~0.6 mg/mL。測定百里醌對阪崎克羅諾腸桿菌生長曲線的影響,并利用修正Gompertz模型擬合其生長狀況,表征菌株的生長參數。結果表明百里醌使阪崎克羅諾腸桿菌生長遲滯期顯著增長,最大生長速率顯著減小。接著探索了百里醌對阪崎克羅諾腸桿菌細胞膜通透性的影響:利用螢火蟲熒光素酶檢測百里醌對胞內ATP的影響,利用cFDA-SE熒光探針檢測百里醌對胞內pH值的影響,利用DiBAC4(3)熒光探針檢測百里醌對細胞膜電位的影響,利用LIVE/DEAD?BacLightTM細菌活性檢測試劑盒檢測百里醌對阪崎克羅諾腸桿菌細胞膜完整性的影響。結果表明百里醌影響了阪崎克羅諾腸桿菌細胞膜的通透性,表現為:質量濃度為MIC和2×MIC的百里醌使細胞內ATP濃度由6.52 μmol/L分別降低為0.27 μmol/L和0.17 μmol/L,胞內pH值由5.69分別降低為5.22和4.99,細胞膜完整菌體比例分別降低至80%和22%,引起細胞膜膜電位超極化。最后通過場發射掃描電子顯微鏡觀測百里醌對阪崎克羅諾腸桿菌細胞形態的影響,結果表明百里醌作用使細胞膜皺縮,細胞干癟,部分菌體喪失了原有的細胞形態。上述結果表明:百里醌對阪崎克羅諾腸桿菌有著良好的抑制效果,是通過影響細胞膜的通透性及引起細胞功能紊亂來實現其抑菌作用。研究結果為百里醌作為天然抑菌劑在食品中發揮其控制阪崎克羅諾腸桿菌的作用提供了理論依據。然而,百里醌對于食品感官品質的影響及食品成分對于百里醌抑菌作用的干擾需要在實際應用前進一步探討。
[1] 任立松, 陳卓, 馬龍, 等. 阪崎腸桿菌新疆分離株侵襲乳鼠器官特征研究[J]. 新疆醫科大學學報, 2010, 33(6): 614-616. DOI:10.3969/j.issn.1009-5551.2010.06.007.
[2] 劉咪, 楊保偉, 夏效東, 等. 阪崎克羅諾腸桿菌致病性機理研究進展[J]. 食品科學, 2014, 35(9): 329-333. DOI:10.7506/spkx1002-6630-201409064.
[3] 李秀桂. 阪崎腸桿菌污染嬰幼兒食品流行病學特征研究進展[J]. 大眾科技, 2009(9): 137-139. DOI:10.3969/j.issn.1008-1151.2009.09.068.
[4] IVERSEN C, FORSYTHE S. Risk prof i le of Enterobacter sakazakii,an emergent pathogen associated with infant milk formula[J]. Trends in Food Science & Technology, 2003, 14(11): 443-454. DOI:10.1016/S0924-2244(03)00155-9.
[5] CUSHNIE T P, LAMB A J. Recent advances in understanding the antibacterial properties of flavonoids[J]. International Journal of Antimicrobial Agents, 2011, 38(2): 99-107. DOI:10.1016/j.ijantimicag.2011.02.014.
[6] GYAWALI R, IBRAHIM S A. Natural products as antimicrobial agents[J]. Food Control, 2014, 46: 412-429. DOI:10.1016/j.foodcont.2014.05.047.
[7] TAJKARIMI M M, IBRAHIM S A, CLIVER D O. Antimicrobial herb and spice compounds in food[J]. Food Control, 2010, 21(9):1199-1218. DOI:10.1016/j.foodcont.2010.02.003.
[8] GALI-MUHTASIB H, ROESSNER A, SCHNEIDER-STOCK R.Thymoquinone: a promising anti-cancer drug from natural sources[J].International Journal of Biochemistry and Cell Biology, 2006, 38(8):1249-1253. DOI:10.1016/j.biocel.2005.10.009.
[9] RAGHEB A, ATTIA A, ELDIN W S, et al. The protective effect of thymoquinone, an anti-oxidant and anti-inflammatory agent, against renal injury: a review[J]. Saudi Journal of Kidney Diseases and Transplantation, 2009, 20(5): 741-752.
[10] ALI B H, BLUNDEN G. Pharmacological and toxicological properties of Nigella sativa[J]. Phytotherapy Research, 2003, 17(4): 299-305.DOI:10.1002/Ptr.1309.
[11] BADARY O A, AL-SHABANAH O A, NAGI M N, et al. Acute and subchronic toxicity of thymoquinone in mice[J]. Drug Development Research, 1998, 44(2/3): 56-61. DOI:10.1002/(Sici)1098-2299(199806/07)44:2/3<56::Aid-Ddr2>3.0.Co;2-9.
[12] HARZALLAH H J, KOUIDHI B, FLAMINI G, et al. Chemical composition, antimicrobial potential against cariogenic bacteria and cytotoxic activity of Tunisian Nigella sativa essential oil and thymoquinone[J]. Food Chemistry, 2011, 129(4): 1469-1474.DOI:10.1016/j.foodchem.2011.05.117.
[13] FRANKOVA A, MAROUNEK M, MOZROVA V, et al. Antibacterial activities of plant-derived compounds and essential oils toward Cronobacter sakazakii and Cronobacter malonaticus[J]. Foodborne Pathogens and Disease, 2014, 11(10): 795-797. DOI:10.1089/fpd.2014.1737.
[14] SANCHEZ E, GARCIA S, HEREDIA N. Extracts of edible and medicinal plants damage membranes of Vibrio cholerae[J]. Applied and Environmental Microbiology, 2010, 76(20): 6888-6894.DOI:10.1128/Aem.03052-09.
[15] SHI Chao, ZHANG Xiaorong, SUN Yi, et al. Antimicrobial activity of ferulic acid against Cronobacter sakazakii and possible mechanism of action[J]. Foodborne Pathogens and Disease, 2016, 13(4): 196-204.DOI:10.1089/fpd.2015.1992.
[16] LI Guanghui, WANG Xin, XU Yunfeng, et al. Antimicrobial effect and mode of action of chlorogenic acid on Staphylococcus aureus[J].European Food Research and Technology, 2014, 238(4): 589-596.DOI:10.1007/s00217-013-2140-5.
[17] LEE S Y, JIN H H. Inhibitory activity of natural antimicrobial compounds alone or in combination with nisin against Enterobacter sakazakii[J]. Letters in Applied Microbiology, 2008, 47(4): 315-321.DOI:10.1111/j.1472-765X.2008.02432.x.
[18] JANG H I, RHEE M S. Inhibitory effect of caprylic acid and mild heat on Cronobacter spp. (Enterobacter sakazakii) in reconstituted infant formula and determination of injury by flow cytometry[J].International Journal of Food Microbiology, 2009, 133(1/2): 113-120.DOI:10.1016/j.ijfoodmicro.2009.05.009.
[19] JOSHI S S, HOWELL A B, D’SOUZA D H. Cronobacter sakazakii reduction by blueberry proanthocyanidins[J]. Food Microbiology,2014, 39: 127-131. DOI:10.1016/j.fm.2013.11.002.
[20] MEMPIN R, TRAN H, CHEN C, et al. Release of extracellular ATP by bacteria during growth[J]. BMC Microbiology, 2013, 13: 1-13.DOI:10.1186/1471-2180-13-301.
[21] BREEUWER P, DROCOURT J, ROMBOUTS F M, et al. A novel method for continuous determination of the intracellular pH in bacteria with the internally conjugated fl uorescent probe 5 (and 6-)-carboxyfluorescein succinimidyl ester[J]. Applied Environmental Microbiology, 1996, 62(1): 178-183.
[22] TURGIS M, HAN J, CAILLET S, et al. Antimicrobial activity of mustard essential oil against Escherichia coli O157:H7 and Salmonella typhi[J]. Food Control, 2009, 20(12): 1073-1079. DOI:10.1016/j.foodcont.2009.02.001.
[23] MATES S M, EISENBERG E S, MANDEL L J, et al. Membrane potential and gentamicin uptake in Staphylococcus aureus[J].Proceedings of the National Academy of Sciences, 1982, 79(21):6693-6697.
[24] WHITEAKER K L, GOPALAKRISHNAN S M, GROEBE D, et al.Validation of FLIPR membrane potential dye for high throughput screening of potassium channel modulators[J]. Journal of Biomolecular Screening, 2001, 6(5): 305-312. DOI:10.1089/108705701753200518.
[25] BOT C, PRODAN C. Probing the membrane potential of living cells by dielectric spectroscopy[J]. European Biophysics Journal with Biophysics Letters, 2009, 38(8): 1049-1059. DOI:10.1007/s00249-009-0507-0.
[26] CAMPOS F M, COUTO J A, FIGUEIREDO A R, et al. Cell membrane damage induced by phenolic acids on wine lactic acid bacteria[J]. International Journal of Food Microbiology, 2009, 135(2):144-151. DOI:10.1016/j.ijfoodmicro.2009.07.031.
[27] CHERRAT L, DUMAS E, BAKKALI M, et al. Effect of essential oils on cell viability, membrane integrity and membrane fl uidity of Listeria innocua and Escherichia coli[J]. Journal of Essential Oil Bearing Plants, 2016, 19(1): 155-166. DOI:10.1080/0972060X.2015.1029986.
[28] DI PASQUA R, BETTS G, HOSKINS N, et al. Membrane toxicity of antimicrobial compounds from essential oils[J]. Journal of Agricultural and Food Chemistry, 2007, 55(12): 4863-4870. DOI:10.1021/Jf0636465.
[29] RHAYOUR K, BOUCHIKHI T, TANTAOUI-ELARAKI A, et al.The mechanism of bactericidal action of oregano and clove essential oils and of their phenolic major components on Escherichia coli and Bacillus subtilis[J]. Journal of Essential Oil Research, 2003, 15(5):356-362.
[30] BAJPAI V K, SHARMA A, BAEK K H. Antibacterial mode of action of Cudrania tricuspidata fruit essential oil, affecting membrane permeability and surface characteristics of food-borne pathogens[J]. Food Control, 2013, 32(2): 582-590. DOI:10.1016/j.foodcont.2013.01.032.
Antimicrobial Activity of Thymoquinone against Cronobacter sakazakii
SHI Chao, SUN Huihui, SUN Zheng, GUO Du, ZHANG Wenting, SUN Yi, CHEN Yifei, XIA Xiaodong*
(College of Food Science and Engineering, Northwest A & F University, Yangling 712100, China)
The objective of the present study was to evaluate the effectiveness of thymoquinone (TQ) against Cronobacter sakazakii strains. The minimum inhibitory concentration (MIC) of TQ against C. sakazakii and its effect on microbial growth kinetics were determined. Changes in intracellular ATP concentration, intracellular pH (pHin), membrane potential and membrane integrity were measured to elucidate the possible antimicrobial mechanism. Cell morphology changes were also observed under a fi eld emission scanning electron microscope. The results showed that the MICs of TQ against C. sakazakii strains ranged from 0.3 to 0.6 mg/mL, and TQ exposure resulted in a longer lag phase and lower specif i c growth rate compared to the control. TQ affected the cell membrane permeability of C. sakazakii, as evidenced by a reduction in intracellular ATP concentration from 6.52 to 0.27 and 0.17 μmol/L and a decrease in pHinfrom 5.69 to 5.22 and 4.99 at MIC and 2 × MIC, respectively. TQ at both tested concentrations caused 80% and 22% reduction in cell integrity, respectively. In addition, cells treated with TQ showed hyperpolarized cell membrane. Field emission scanning electron microscopy illustrated that C. sakazakii cells exposed to TQ exhibited a more wrinkled surface compared with the smooth surface of untreated cells. In conclusion, TQ exerts its antimicrobial action partly by causing excessive permeability of the cell membrane and changing cell morphology. These fi ndings suggest that TQ has potential application in controlling C. sakazakii in foods.
Cronobacter sakazakii; thymoquinone; cell membrane; membrane potential
10.7506/spkx1002-6630-201801009
Q939.9
A
1002-6630(2018)01-0058-07
石超, 孫慧慧, 孫正, 等. 百里醌對阪崎克羅諾腸桿菌的抑制作用[J]. 食品科學, 2018, 39(1): 58-64.
10.7506/spkx1002-6630-201801009. http://www.spkx.net.cn
SHI Chao, SUN Huihui, SUN Zheng, et al. Antimicrobial activity of thymoquinone against Cronobacter sakazakii[J]. Food Science,2018, 39(1): 58-64. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201801009. http://www.spkx.net.cn
2016-09-04
“十二五”國家科技支撐計劃項目(2015BAD16B08);中央高校基本科研業務費專項資金項目(2452017146)
石超(1989—),女,講師,博士,研究方向為天然活性物質的抗菌功能。E-mail:meilixinong@126.com
*通信作者簡介:夏效東(1981—),男,教授,博士,研究方向為天然活性物質的營養及抗菌功能。E-mail:foodscixiaodong@yahoo.com
