不同濃度CaCl2對宜昌潤楠幼苗的抗寒性研究
2018-01-30 15:37:57況紅玲喻峰杜丹張欣王昆賀心茹費永俊
湖北林業科技
2017年6期
況紅玲+喻峰+杜丹+張欣+王昆+賀心茹+費永俊

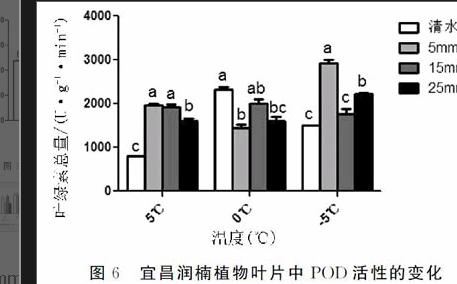
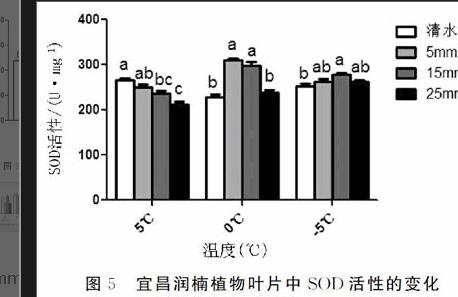
摘要:本文以多年生宜昌潤楠(Machilusichangensis)為材料,采取室內控溫系統模擬低溫脅迫條件,研究不同濃度CaCl2處理對低溫脅迫下宜昌潤楠幼苗葉綠素含量、丙二醛含量、可溶性蛋白含量、可溶性糖含量、超氧化物歧化酶活性、過氧化物酶活性以及激素ABA含量的影響,初步探究CaCl2對潤楠幼苗抗寒性的調節機理。結果表明:(1)宜昌潤楠幼苗在-5℃條件下噴施5mmol/LCaCl2后葉綠素含量最高。(2)宜昌潤楠幼苗在5℃條件下噴施25mmol/LCaCl2后丙二醛(MDA)含量最低。(3)宜昌潤楠幼苗在低溫脅迫條件下,通過噴施CaCl2能影響可溶性蛋白的含量。(4)宜昌潤楠幼苗在-5℃條件下噴施25mmol/LCaCl2后可溶性糖含量最高。(5)宜昌潤楠幼苗在低溫脅迫條件下,通過噴施CaCl2能提高SOD和POD的活性,其中在0℃條件下,宜昌潤楠幼苗經5mmol/LCaCl2噴施后的SOD活性達到最高,在-5℃條件下宜昌潤楠幼苗經5mmol/LCaCl2噴施后的POD活性達到最高。(6)宜昌潤楠幼苗在低溫脅迫條件下,通過噴施CaCl2能提高激素ABA含量,在-5℃條件下噴施5mmol/LCaCl2后ABA含量最高。
關鍵詞:宜昌潤楠;幼苗;CaCl2;抗寒性
中圖分類號:S79227文獻標識碼:A文章編號:1004-3020(2017)06-0014-06DifferentConcentrationsofCaCl2onColdResistanceofYichangRunanSeedlingKuangHongling(1)YuFeng(1)DuDan(1)ZhangXin(1)WangKun(1)HeXinru(2)FeiYongjun(2)
(1.FayeEcologicalEnvironmentGroupco.,LTD.Wuhan430206;2.YangtzeUniversityJingzhou434100)
Abstract:Inthispaper,theeffectsofdifferentconcentrationsofCaCl2onmalondialdehyde(MDA)content,solubleproteincontentandsolublesugarcontentofYunnanseedlinginYichangwerestudiedbyusingthetemperaturecontrolsystemundertheconditionoflowtemperaturestress.Theactivityofchlorophyll,thecontentofchlorophyllandtheABAcontentofthehormonewerestudied.ThemechanismofCaCl2onthecoldresistanceofRunonseedlingwasexplored.Theresultsshowedthat:(1)Thecontentofchlorophyllwasthehighestwhenspraying5mmol/LCaCl2at-5℃.(2)Thecontentofmalondialdehyde(MDA)wasthehighestwhenspraying25mmol/LCaCl2at5℃undertheconditionof5℃.(3)ThecontentofsolubleproteinwasincreasedbysprayingCaCl2undertheconditionoflowtemperaturestress.(4)Thecontentofsolublesugarwasthehighestwhenspraying25mmol/LCaCl2at-5℃.(5)TheactivityofSODandPODcouldbeincreasedbysprayingCaCl2undertheconditionoflowtemperaturestress,andtheactivityofSODwasthehighestaftersprayingwith5mmol/LCaCl2at0℃.TheactivityofPODwasthehighestafterspraying5mmol/LCaCl2at-5℃.(6)ThecontentofABAwasincreasedbysprayingCaCl2undertheconditionoflowtemperaturestress,andthecontentofABAwasthehighestat5℃for5mmol/LCaCl2.
Keywords:YichangRunan;seedling;CaCl2;coldresistance
潤楠屬的植物在全球大約有100個種,主要散布在東部的熱帶、亞洲東南部和亞熱帶區域。我國大約有68種3個變種,分布于西南部和南部地區[1]。潤楠屬植物包括宜昌潤楠(M.ichangensis)、黃枝潤楠(M.versicolora)、滇潤楠(M.yun-nanensis)、梨潤楠(M.pomifera)、華潤楠(M.chinensis)、絨毛潤楠(M.velutina)、刨花潤楠(M.pauhoi)、黃絨潤楠(M.grijsii)和粉葉潤楠(M.glaucifolia)等。
宜昌潤楠為樟科(Lauraceae)潤楠屬(Machilus),是常綠闊葉樹種,分布于陜西南部、湖北、四川等地,是我國的特有樹種,屬于國家二級保護植物。……
登錄APP查看全文
