結核分枝桿菌耐環絲氨酸相關基因研究進展
2018-02-15 07:44:32何興陳玲
實用心腦肺血管病雜志 2018年11期
何興,陳玲
耐多藥結核病(MDR-TB)指同時對利福平和異煙肼耐藥的結核病。據統計,2017年全球范圍內MDR-TB患者數量約為160 684例,其中廣泛耐藥結核病(XDR-TB)患者數量約占8.5%[1],MDR-TB成為全球范圍內結核病防控工作面臨的主要挑戰。環絲氨酸是1952年研發的一種抗生素,具有較強的抗結核分枝桿菌作用,因而WHO于2011年推薦環絲氨酸作為治療MDR-TB的二線核心口服抗結核藥物之一[2-4]。與其他抗結核藥物相比,環絲氨酸耐藥率較低,但近年來結核分枝桿菌對環絲氨酸耐藥增多。有研究者于1993—2007年對荷蘭131例MDR-TB患者進行藥敏試驗發現,僅1例患者對環絲氨酸耐藥,耐藥率為0.8%[5];2005年俄羅斯結核分枝桿菌對環絲氨酸耐藥率為7.4%;1995—2011年中國香港結核分枝桿菌對環絲氨酸耐藥率為8.6%[6]。在結核病治療新藥匱乏情況下,2012年5月國家食品藥品監督管理局批準我國“全球基金耐多藥結核病項目”開展地區使用環絲氨酸[7]。目前,結核分枝桿菌對環絲氨酸耐藥的具體機制尚未完全明確,本文綜述了結核分枝桿菌耐環絲氨酸相關基因,旨在為結核分枝桿菌對環絲氨酸的耐藥機制研究提供參考。
1 作用機制
環絲氨酸具有廣譜抗菌作用,對多數革蘭陰性菌及革蘭陽性菌均具有一定抑制作用,對結核分枝桿菌具有較強的抑制作用并有利于防止結核分枝桿菌對丙硫異煙胺產生耐藥[8]。自1999 年以來,伊朗、土耳其、印度等國家采用含環絲氨酸抗結核化療方案治療MDR-TB并取得良好效果。伊朗學者曾報道,采用含環絲氨酸抗結核化療方案治療的MDR-TB患者可達到治愈效果[9];土耳其學者曾報道,采用含環絲氨酸抗結核化療方案治療的MDR-TB患者治愈率為77%[10];日本學者曾報道,采用含環絲氨酸、吡嗪酰胺、乙胺丁醇等抗結核化療方案治療MDR-TB孕婦療效確切[11];西班牙學者曾報道,采用環絲氨酸、左氧氟沙星治療的17例MDR-TB患者中12例達到治愈效果,且無復發或死亡患者[12];印度學者曾報道,采用含環絲氨酸抗結核化療方案治療的39例MDRTB患者中29例治療6個月后痰菌轉陰并維持到治療結束[13]。
環絲氨酸抗結核分枝桿菌的血藥濃度參考范圍為1.5~30.0 mg/L[14],其對體外H37Rv菌株的最低抑菌濃度(MIC)為 25.0 mg/L[15],環絲氨酸劑量為250.0~500.0 mg/次時血藥峰濃度可維持在20.0~35.0 mg/L,不僅可發揮良好的抗結核分枝桿菌作用,還有利于減少不良反應的發生,因此血藥峰濃度<15.0 mg/L時需增加環絲氨酸劑量,血藥峰濃度>40.0 mg/L需減少環絲氨酸劑量[16]。
結核分枝桿菌細胞壁是一種網狀復雜結構,主要由肽聚糖、霉菌酸、阿拉伯半乳聚糖等物質通過共價鍵連接而成,D-丙氨酸是結核分枝桿菌細胞壁中肽聚糖的重要成分[17],丙氨酸消旋酶(Alr)是結核分枝桿菌細胞壁合成過程中必不可少的成分[5],且Alr以磷酸毗哆醛(PLP)為輔酶催化L-丙氨酸和D-丙氨酸的轉化;此外,在丙氨酸合成代謝途徑中,丙氨酸連接酶催化二分子D-丙氨酸合成D-丙氨酸二聚體(ddl),因此丙氨酸連接酶在丙氨酸合成代謝中發揮著重要作用。環絲氨酸為D-丙氨酸結構類似物,主要通過抑制丙氨酸合成代謝途徑中alr基因編碼的Alr、ddlA基因編碼的丙氨酸連接酶而減少肽聚糖的產生,繼而抑制結核分枝桿菌細胞壁形成并減弱其耐酸能力,達到抗結核分枝桿菌的目的[18-19]。
2 耐藥機制
目前已知的結核分枝桿菌耐環絲氨酸基因包括ald基因(Rv2780)、alr基因(Rv3423C)、cycA基因(Rv1704C)、ddlA基因(Rv2981C,ddl)等。
2.1 ald基因 ald基因ID為888493,全長1 116 bp,編碼L-丙氨酸脫氫酶并參與丙氨酸代謝。研究發現,ald基因缺失的結核分枝桿菌無法將L-丙氨酸轉化成丙酮酸并導致L-丙氨酸含量增加,而L-丙氨酸是環絲氨酸競爭性抑制物質前體,L-丙氨酸含量增加可能會導致Alr、丙氨酸連接酶持續活動而降低環絲氨酸的競爭性抑制作用,最終造成結核分枝桿菌產生大量肽聚糖并對環絲氨酸產生耐藥性[19-20]。藥敏試驗結果證實,ald基因缺失與結核分枝桿菌對環絲氨酸耐藥有關,而補充ald基因后部分結核分枝桿菌會保存對環絲氨酸的敏感性[19-20]。
2.2 alr基因 alr基因ID為887634,全長1 227 bp,編碼Alr并參與丙氨酸代謝途徑中L-丙氨酸向D-丙氨酸的轉化。研究發現,alr基因啟動子突變的結核分枝桿菌對環絲氨酸的耐藥性較高(MIC>60 μg/ml)[19];D-丙氨酸消旋酶(alrA)過度表達可導致牛型分枝桿菌卡介苗對環絲氨酸產生耐藥[21],同時超量產生的突變體alrA啟動子出現G-T置換可導致B-半乳糖苷酶基因表達[22]。DESIGARDINS等[23]研究發現,體外培養的alrA基因突變的結核分枝桿菌對環絲氨酸的耐藥性增加。此外,還有研究證實結核分枝桿菌alrA非同義突變與環絲氨酸耐藥表型有關[24-25],有學者在24株耐環絲氨酸結核分枝桿菌中發現1株alrA同義突變,1株非同義突變,涉及的突變位點及氨基酸改變為261位絲氨酸-天冬酰胺,而對環絲氨酸敏感的結核分枝桿菌337位發生GGT-GGC,為甘氨酸同義突變[26](見表1)。
2.3 cycA基因 cycA基因ID為888812,全長1 671 bp,編碼D-絲氨酸、D-丙氨酸等運載通透蛋白并參與環絲氨酸的轉運吸收[27]。BAISA等[28]研究發現,在甘油培養基和微量葡萄糖中大腸埃希菌cycA基因突變株CFT073和K-12對環絲氨酸的耐藥性增加。目前研究已證實耐環絲氨酸大腸埃希菌變異株存在cycA基因單一位點突變[29],早期曾有文獻報道cycA基因點突變可導致大腸埃希菌對環絲氨酸的耐藥性增加[30-31],而在牛型分枝桿菌卡介苗中發現cycA基因非同義突變(Gly122Ser)可以解釋部分環絲氨酸固有耐藥現象[27];FéHERT等[29]對20株耐環絲氨酸大腸埃希菌進行研究發現,其cycA基因突變位點均不相同。此外,還有研究在24株耐環絲氨酸結核分枝桿菌中發現3株cycA基因同義突變,3株cycA基因非同義突變,其中非同義突變包括第188位CCTGCT〔(脯氨酸-丙氨酸〕、第318位CGA-CTA〔精氨酸-亮氨酸〕及第508位GCA-TCA(丙氨酸-絲氨酸),但并未發現突變熱點區域;在對環絲氨酸敏感的結核分枝桿菌中發現cycA基因第406位TCG-TCA(絲氨酸)及第521位CGTCGC(精氨酸),均為同義突變[26](見表1)。
2.4 ddlA基因 ddlA基因ID為888415,全長1 122 bp,編碼丙氨酸連接酶并催化二分子D-丙氨酸合成ddl、參與丙氨酸代謝。20世紀70年代,有學者對耐環絲氨酸結核分枝桿菌突變體進行分離發現,ddlA基因對環絲氨酸的抗結核分枝桿菌作用影響極小,但后續研究發現ddlA基因、alr基因同時過度表達可導致結核分枝桿菌對環絲氨酸的耐藥性增加[32]。有研究在24株耐環絲氨酸結核分枝桿菌中發現1株ddlA基因同義突變,為第92位CGT-CGC突變(精氨酸)(見表1)[26]。
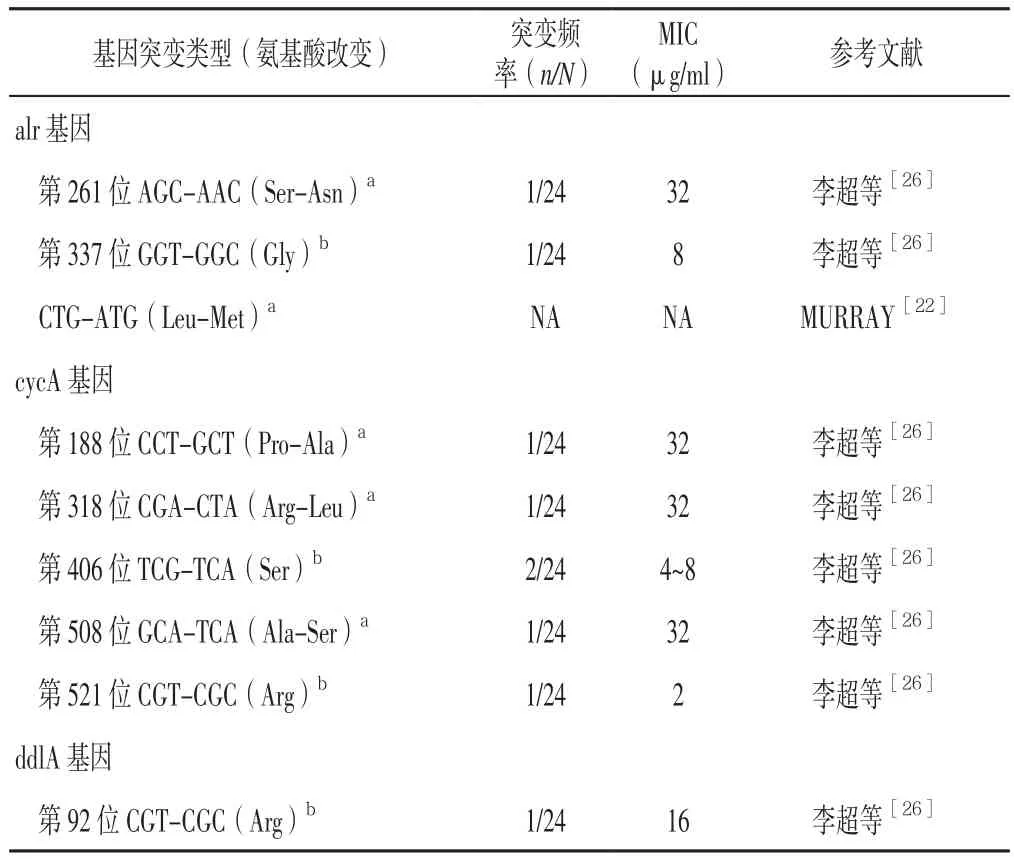
表1 已知的環絲氨酸耐藥基因突變類型、氨基酸改變及MICTable 1 Known drug resistance related gene mutation types to cycloserine,changes of amino acids and the MIC
3 小結與展望
綜上所述,環絲氨酸作為二線核心抗結核口服藥物,具有獨特的結構及作用機制,治療耐藥結核病的前景廣闊,但其耐藥現象不容忽視,應深入研究其耐藥機制以減少耐藥的發生。近年來隨著對結核分枝桿菌耐藥機制的研究深入,發現恥垢分枝桿菌中Alr及丙氨酸連接酶過表達會產生對環絲氨酸的耐藥性,且Alr對環絲氨酸耐藥性的影響程度較丙氨酸連接酶高[33],而alr基因、ddlA基因突變與結核分枝桿菌對環絲氨酸耐藥有關[21,34-35];此外,卡介苗絲氨酸運載體突變也可導致結核分枝桿菌對環絲氨酸產生耐藥性[27,33]。
目前,基因突變可以解釋部分結核分枝桿菌對環絲氨酸耐藥,其中ald基因、alr基因、cycA基因、ddlA基因突變導致結核分枝桿菌對環絲氨酸耐藥已達成一定共識,但各研究中所涉及的突變位點均不相同,因此還可能存在其他突變位點、耐藥靶酶等,因此,結核分枝桿菌對環絲氨酸耐藥的具體機制等仍需進一步研究證實。
猜你喜歡
英語世界(2023年6期)2023-06-30 06:29:10
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
中國生殖健康(2020年2期)2021-01-18 02:51:26
科學大眾(2020年12期)2020-08-13 03:22:22
小學生導刊(2018年13期)2018-06-29 03:49:00
現代檢驗醫學雜志(2016年4期)2016-11-15 02:01:14
中國當代醫藥(2015年17期)2015-03-01 02:03:58
中國神經精神疾病雜志(2014年1期)2014-03-01 03:23:22
