澤蛙原蛙片蟲的形態(tài)學(xué)研究
2018-03-30 03:33:04汪潤秋李文祥吳山功王桂堂
水生生物學(xué)報(bào) 2018年2期
鄒 紅 李 燦 汪潤秋 李文祥 吳山功 王桂堂 李 明
(1. 中國科學(xué)院水生生物研究所, 農(nóng)業(yè)部淡水養(yǎng)殖病害防治重點(diǎn)實(shí)驗(yàn)室, 淡水生態(tài)與生物技術(shù)國家重點(diǎn)實(shí)驗(yàn)室, 武漢 430072; 2.武漢輕工大學(xué)動(dòng)物科學(xué)與營養(yǎng)工程學(xué)院, 武漢 430023)
蛙片蟲是一類多核、無口, 滲透性營養(yǎng)的鞭毛類原生動(dòng)物。絕大多數(shù)寄生在變溫脊椎動(dòng)物體內(nèi),特別是無尾兩棲動(dòng)物(蛙類和蟾蜍的成體及蝌蚪)的腸道內(nèi)[1]。由于其皮層構(gòu)造與纖毛蟲頗為相似, 該類群長期以來一直被歸為較原始的纖毛蟲類(Protociliate)[2—4]。后來發(fā)現(xiàn)這類原生動(dòng)物沒有大小核的分化, 無性生殖為縱二分裂, 有性生殖能產(chǎn)生具鞭毛的配子, 故將其納入鞭毛蟲類, 并單列為蛙片亞門(Opalinata)[5,6]; 下分1綱、1目、1科, 即: 蛙片綱(Opalinatea), 蛙片目(Opalinida), 蛙片科(Opalinidae)[7]。根據(jù)蛙片蟲形狀和細(xì)胞核數(shù)目的不同, 可分為兩個(gè)亞科, 即原蛙片蟲亞科(Protoopalininae)和蛙片蟲亞科(Opalininae)。
原蛙片蟲屬(Protoopalina)隸屬于原蛙片蟲亞科, 是兩棲動(dòng)物腸道中最常見的蛙片蟲類。該屬由Metcalf在1918年建立, 其典型特征是蟲體常呈長棍狀, 具2個(gè)或4個(gè)圓形或紡錘形胞核[3]。澤蛙原蛙片蟲(P. limnocharis)是Nie(1932)首次從澤蛙腸道內(nèi)發(fā)現(xiàn)并命名的[8]。囿于當(dāng)時(shí)染色技術(shù)和觀測工具的落后, 一些關(guān)鍵的形態(tài)特征如縫線等未能很好地予以揭示。因而, 利用現(xiàn)代技術(shù)手段對(duì)其形態(tài)學(xué)特征進(jìn)行重新描述和修正顯得尤為必要。
1 材料與方法
1.1 宿主蛙來源
澤蛙(Fejervarya limnocharis)成體210只于2016年6月采自湖北省洪湖市(北緯29°40′—29°58′N;東經(jīng)113°12′—113°26′E), 活體運(yùn)回并暫養(yǎng)于實(shí)驗(yàn)室。
1.2 光鏡觀察
活體觀察將宿主蛙用100%酒精麻醉后解剖、取腸道, 置于體視顯微鏡Zeiss Stemi SV6/Axio Cam MRc5下觀察腸道內(nèi)含物。用0.65%生理鹽水浸漬內(nèi)含物后收集部分蟲體置凹玻片上, 置于光學(xué)顯微鏡下觀察其活體形態(tài)。
固定染色標(biāo)本觀察將部分蟲體置胚胎皿中, 飽和升汞(HgCl2)溶液固定, 分別用埃利希蘇木精(Ehrlich’s haematoxylin)和蛋白銀(Protargo)染色,加拿大樹膠封片, 用于觀察蟲體皮層及內(nèi)部構(gòu)造。全部觀察、測量、拍照均使用德國蔡司顯微成像系統(tǒng)Zeiss Axioplan 2 imaging and Axiophot 2完成。
1.3 掃描電鏡觀察
將游離出的大量蟲體4000×g離心10min, 2.5%(v/v)戊二醛水溶液與2% (w/v)鋨酸水溶液的1∶1混合液于4℃固定2h。用0.2 mol/L磷酸緩沖液(pH 7.4)清洗后置于涂有1%多聚賴氨酸1 cm×1 cm的玻片上。常規(guī)脫水、乙酸異戊酯置換處理后, 臨界點(diǎn)干燥2h (HCP-2 Crtical Point Dryer)。離子濺射儀(EikoIB-3 Ion Coater)鍍膜噴金后, 掃描電鏡Quanta 200 (FEI, Holand)觀察。
2 結(jié)果
在我們采集的210只澤蛙中, 有38只感染了澤蛙原蛙片蟲。感染率為18.1%, 寄生于澤蛙直腸。固定染色標(biāo)本保存于中國科學(xué)院水生生物研究所寄生蟲學(xué)與協(xié)同進(jìn)化學(xué)科組。
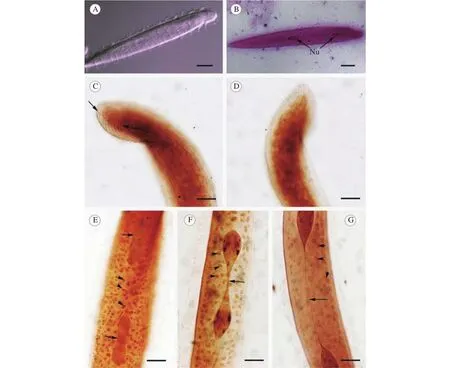
圖 1 澤蛙原蛙片蟲的光鏡顯微照片F(xiàn)ig. 1 Light microscope images of P. limnocharis
澤蛙原蛙片蟲蟲體呈長棒狀, 略扁平(圖 1A,B和2A)。前端凸出、稍狹窄, 約占蟲體1/10 (圖2A, C); 其后為蟲體最寬處, 至尾端漸趨細(xì)削(圖2A, D)。體長為313.8 μm (283.2—350.4 μm, n=30),體寬27.1 μm (21.6—38.4 μm, n=30)。長寬比約為10∶1。蟲體周身密被鞭毛, 游動(dòng)迅速且常翻轉(zhuǎn)前行(圖 2C)。體表皮褶形成的條紋與蟲體縱軸傾斜。縫線位于蟲體頂端, 長約20 μm。所有體動(dòng)基列(約30—40列)均從縫線兩側(cè)發(fā)出, 沿蟲體前端一直延伸至尾部(圖 1C, D)。
胞核2個(gè), 梨形; 其一在蟲體前端約1/6處, 另一個(gè)在位于蟲體中部略前處(圖 1B)。核長19.3 μm(14.4—24.0 μm, n=30)), 寬5.5 μm (4.2—8.4 μm,n=30)。胞核分裂時(shí)從中部縊縮(圖 1E), 逐漸向兩端拉長并形成一條細(xì)絲(圖 1F)。此細(xì)絲在蟲體整個(gè)生活周期都一直存在, 成熟蟲體中其長度為112.92 μm (76.8—127.2 μm, n=30)(圖 1G)。另外,在胞質(zhì)中可見大量大小不一、圓球狀的顆粒(圖1E, F和G)。
蟲體相關(guān)形態(tài)學(xué)特征測量值見表 1,模式圖見圖 3 。
3 討論
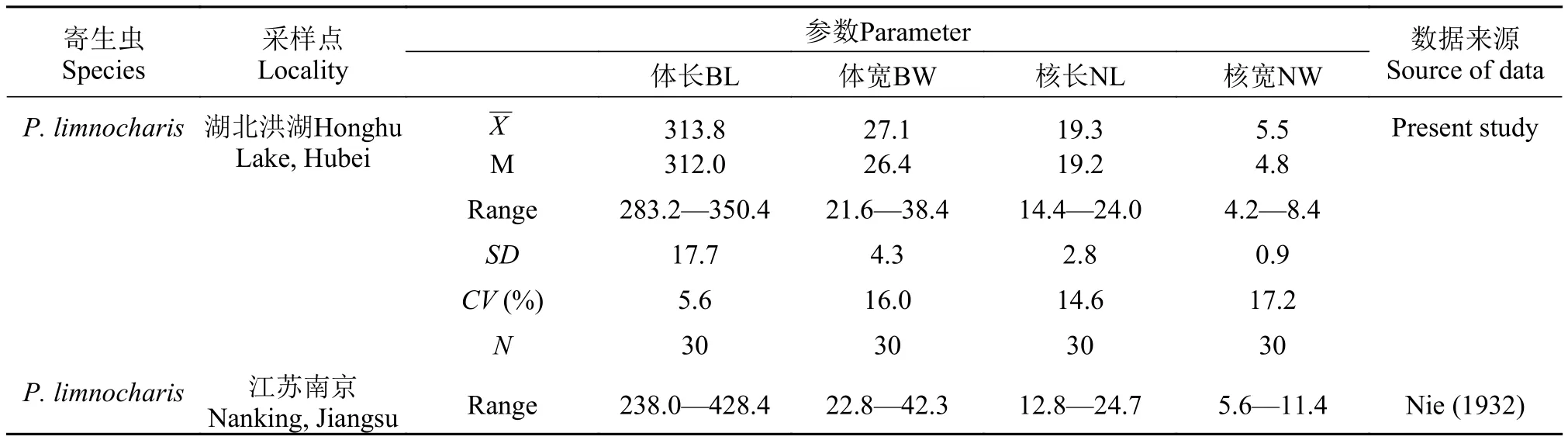
表 1 澤蛙原蛙片蟲形態(tài)學(xué)特征測量值(單位: 微米)Tab. 1 Biometrical values (in μm) on Protoopalina limnocharis inhabiting Fejervarya limnocharis (=Rana limnocharis)
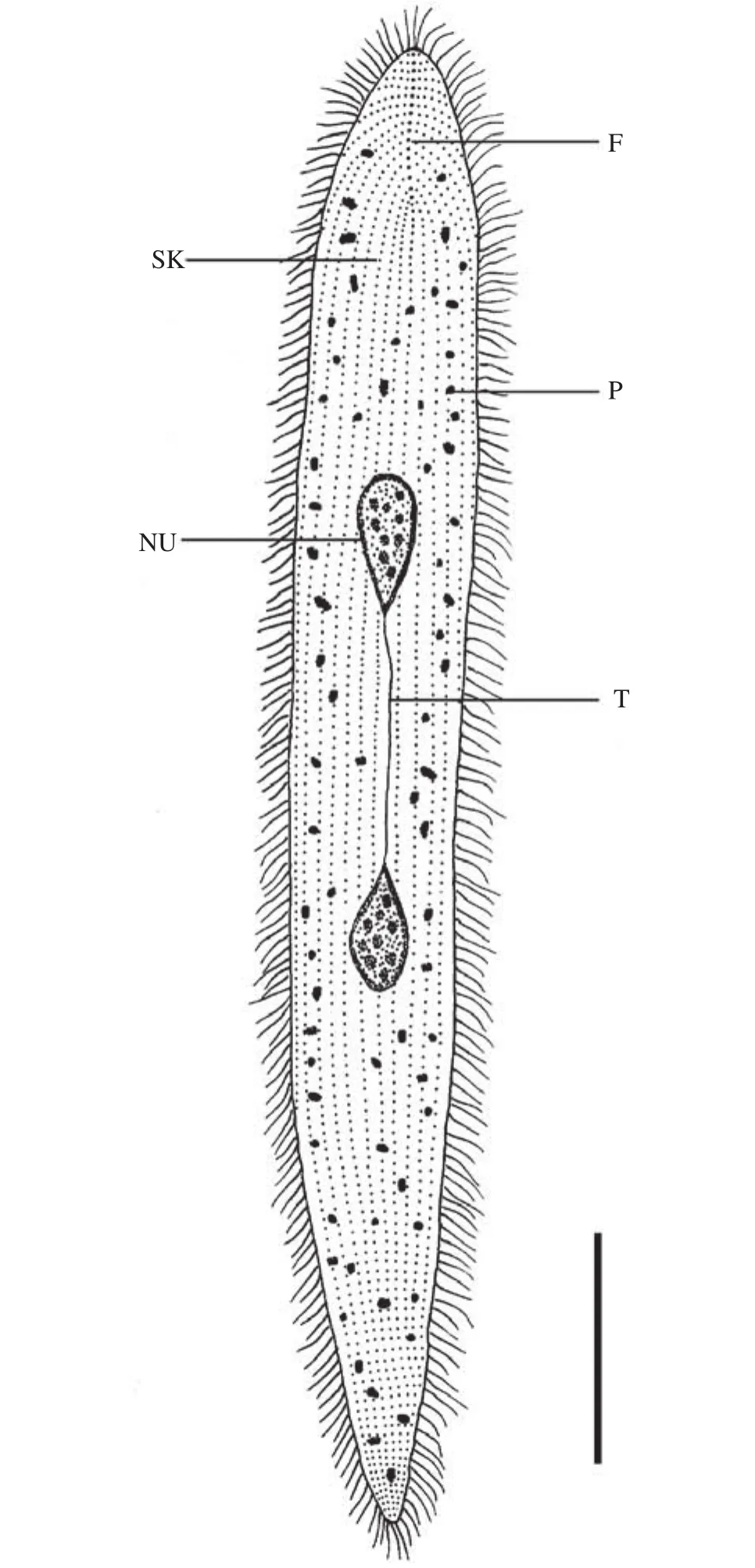
圖 3 澤蛙原蛙片蟲的模式圖Fig. 3 Schematic drawing of P. limnocharis
澤蛙原蛙片蟲是由Nie (1932)首次從澤蛙腸道內(nèi)發(fā)現(xiàn)并命名的[8]。該蛙片蟲目前僅在澤蛙中發(fā)現(xiàn), 應(yīng)該具有較為嚴(yán)格的宿主專一性。本研究中測量蟲體樣本所得形態(tài)特征參數(shù)值與Nie所記述值頗為相符(包括體長、體寬, 核長、核寬; 表 1)。然而,有2個(gè)重要的分類特征需要進(jìn)行補(bǔ)充和修正: (1)縫線位于蟲體頂端, 貫穿背腹側(cè); 所有體動(dòng)基列均從縫線兩側(cè)發(fā)出, 這在前人的研究中沒有描述。該結(jié)構(gòu)特征與我們已經(jīng)研究過的寄生于沼蛙和黑斑蛙腸道內(nèi)的原蛙片蟲Protoopalina pingi的縫線結(jié)構(gòu)是一致的[9], 而且它們的體動(dòng)基列均與蟲體縱軸平行延伸, 所不同的只是兩者體動(dòng)基列的數(shù)目不同: P.limnocharis有30—40列, P. pingi有18—29列[9]。(2)胞核分裂時(shí)在兩新核間產(chǎn)生的連接細(xì)絲, 在蟲體整個(gè)生活周期一直存在。這一結(jié)構(gòu)在其他已知的原蛙片蟲屬種類中未見報(bào)道, 其有何生物學(xué)功能亦不清楚。此外, 我們在觀察分裂完成不久產(chǎn)生的P.limnocharis新生蟲體時(shí), 發(fā)現(xiàn)它們體內(nèi)的2個(gè)胞核距離均較近且有細(xì)絲牽連; 但不同新個(gè)體中胞核的位置是有區(qū)別的: 其中一部分蟲體內(nèi)2個(gè)胞核均位于身體前端, 另一部分則處于蟲體中部位置, 這2種位置分別對(duì)應(yīng)成熟蟲體中2個(gè)胞核的位置。據(jù)此推測P. limnocharis子代蟲體中的2個(gè)胞核均來自于母體中同一個(gè)核, 而其兩核間的連接細(xì)絲可能是胞核分裂進(jìn)化歷程中比較原始的殘跡。
就胞核數(shù)量而言, 已知原蛙片蟲的種類絕大多數(shù)為2個(gè)核, 如P. limnocharis、P. caudata michyla[10]、P. heleophrynes[11]、P. pomacantha[12]等。也有個(gè)別種類核數(shù)不定, 如P. pingi多為四核, 有時(shí)兩核或六核[9]; P. quadrinucleata核數(shù)最多時(shí)可達(dá)8個(gè)[13]。作者認(rèn)為這種核數(shù)不定的現(xiàn)象應(yīng)該是在蟲體分裂過程中胞質(zhì)分裂和核分裂不同步造成的; 核數(shù)不定的種類應(yīng)該是原蛙片蟲屬中進(jìn)化程度較低的種類, 而核數(shù)穩(wěn)定為兩核的種類進(jìn)化程度較高。
另外, 關(guān)于蛙片蟲類的分類依據(jù), 長期以來一直爭議頗多; 但主流上基本沿用Metcalf的界定方法[4]: 雙核、圓柱狀種類歸為Protoopalina, 雙核、扁平狀種類歸為Zelleriella, 多核、圓柱狀種類歸為Cepedea, 多核、扁平狀種類歸為Opalina。根據(jù)我們目前已有的形態(tài)學(xué)工作基礎(chǔ)(特別是銀染標(biāo)本)和分子生物學(xué)數(shù)據(jù)(全rDNA序列), 我們認(rèn)為這種分類方法是基本正確的。這將在我們的研究中進(jìn)一步補(bǔ)充、求證并最終得到較為合理的結(jié)論。
參考文獻(xiàn):
[1]Mignot J P. Patterning in opalinids. I. Implications of new morphological and ultrastructural findings on the genesis of kinetics [J]. European Journal of Protistology, 1994,30(2): 196—210
[2]Boisson C. Deux nouvelles opalines de l’Ouest africain[J]. Bulletin de i’Institut Fran?ais d’Afrique Noire, 1965,27A: 858—865
[3]Metcalf M M. Opalina and the origin of the Ciliata [J].Anatomical Record, 1918, 14: 88—89
[4]Metcalf M M. The opalinid ciliate infusorians [J]. Bulletin of the United States National Museum, 1923, 120:1—484
[5]Corliss J O. The opalinid infusorians: flagellates or ciliates [J]? Journal of Eukaryotic Microbiology, 1955,2: 107—114
[6]Corliss J O. Flagellates, opalinids and the search for the most primitive ciliate and its progenitor [J]. Ceylon Journal of Science, 1979, 13: 65—78
[7]Sheng Y F, Wang J G. Protozoology [M]. Beijing: Science Press. 1999, 223—231 [沈韞芬, 汪建國. 原生動(dòng)物學(xué). 北京:科學(xué)出版社. 1999, 223—231]
[8]Nie D S. On some intestinal ciliates from Rana limnocharis Gravenhorst [J]. Zoological Series, 1932, 6:183—199
[9]Li W, Wang C, Huang F, et al. Redescription of Protoopalina pingi Nie, 1935 inhabiting the recta of Hylarana guentheri and Pelophylax nigromaculatus in China [J].Parasite, 2014, 21: 46
[10]Nie D S. Intestinal ciliates of amphibian of Nanking [J].Contributions from the Biological Laboratory of the Science Society of China: Zoological Series, 1935, 11:67—95
[11]Delvinquier B J, Markus M B, Passmore N I. Opalinidae in African Anura. Genus Protoopalina [J]. Systematic Parasitology, 1995, 30: 81—120
[12]Grim J N, Pérez-Espa?a H, Martínez-Díaz S F. The morphology of Protoopalina pomacantha, n. sp., symbiont in the rectum of the Angelfishes, Pomacanthus zonipectus and Holacanthus passer. A light, scanning electron and transmission electron microscopic study [J]. European Journal of Protistology, 2000, 36(3): 343—350
[13]Lu K. On some parasitic ciliates from frogs of Pehpei [J].Sinensia, 1945, 16: 65—72
