SGK1在血清和母胎界面的表達及其與稽留流產的關系
2018-04-02 01:27:37顧文婷劉福民韓秋峪
中國婦幼健康研究 2018年3期
顧文婷,劉福民,韓秋峪
(1.徐州醫科大學研究生學院,江蘇 徐州 221000;2. 徐州醫科大學附屬醫院婦產科,江蘇 徐州 221000)
稽留流產是因各種因素使胚胎停止發育或死亡滯留宮腔的一種自然流產形式。目前稽留流產在我國的發生率高達13.4%,并且呈逐年增加的趨勢。盡管已有許多相關研究,但稽留流產的發生機制尚不十分明確。血清和糖皮質激素誘導蛋白激酶-1(serum-and glucocorticoid-regulated kinase,SGK1)是絲氨酸/蘇氨酸蛋白激酶的亞家族成員,已被證實在中樞神經系統、心血管系統等多種系統疾病的發生發展中發揮作用。其參與子宮內膜容受性的維持,與人類妊娠有密切關系。Toll樣受體(toll-like receptors,TLRs)是機體內重要的免疫受體。TLR4(Toll-like receptor 4)是第一個在人類中被發現的Toll樣受體家族成員,其不僅介導先天性免疫,還介導獲得性免疫應答,并參與妊娠的維持和妊娠疾病的發生。研究顯示TLR4轉導來自革蘭陰性菌毒素脂多糖(lipopolysaccharide,LPS)的信號,隨后激活核因子-κB(nuclear factor-κB,NF-κB)以誘導促炎細胞因子的表達,不利于妊娠[1]。而SGK1活化可抑制脂多糖(lipopolysaccharide,LPS)/TLR4介導的細胞凋亡,并促進了母胎界面的Th2反應[2]。目前SGK1與稽留流產相關性的研究少有報道,其機制亦不清楚。因此,本研究旨在通過檢測稽留流產患者血清SGK1與TLR4的水平,同時檢測絨毛和蛻膜組織中SGK1、TLR4和NF-κB的表達,探討SGK1與稽留流產的相關性及其與流產發生的可能機制。
1資料與方法
1.1臨床資料
選擇2017年2至7月期間于徐州醫科大學附屬醫院婦產科住院行清宮術的稽留流產患者及正常早孕婦女為研究對象,患者年齡18~46歲,平均29.66±5.91歲。稽留流產組入組條件:①孕8~12周通過超聲發現胚胎停止發育,無心管搏動;②夫婦雙方和(或)胚胎染色體正常;③月經周期正常;④性激素、甲狀腺功能、血糖、胰島素等內分泌檢查均正常;⑤排除生殖道感染性疾病,如霉菌、支原體、衣原體等感染;⑥巨細胞病毒、弓形蟲等病原體檢查IgM均為陰性;⑦檢查抗磷脂抗體為陰性排除抗磷脂綜合征。正常早孕組入組條件:①此次妊娠期間無陰道流血、腹痛等先兆流產癥狀和體征;②超聲證實胚胎發育正常,有心管搏動。排除標準:①既往有自然流產、死胎、死產史;②有家族遺傳病史;③婦科檢查、超聲檢查器質性病變和生殖器官解剖畸形遺傳;④有與入組條件不符合的情況。經嚴格納入排除標準篩選后留取稽留流產組32例、正常早孕組30例,兩組患者間年齡、停經天數、孕次、產次比較,差異均無統計學意義(均P>0.05)。患者均知情同意,并簽署知情同意書,本研究經醫院倫理委員會審核同意。
1. 2研究方法
1.2.1標本制備
所有研究對象入院24h內空腹采集其肘靜脈血3mL,室溫靜止1h后,3 000 r/min離心10min,取上清,-80℃冰箱保存。所有研究對象在清宮術后取新鮮蛻膜及絨毛組織置于10%中性甲醛固定24~48h后常規石蠟包埋、切片、烤片,切片厚度4μm,進行免疫組化染色。
1.2.2 血清中 SGK1、TLR4表達檢測
采用酶聯免疫吸附試驗(試劑購自武漢華美生物工程公司)檢查研究對象血清中SGK1與TLR4的水平,所有實驗操作均嚴格按照試劑盒說明書進行。
1.2.3絨毛及蛻膜組織中 SGK1、TLR4和NF-κB的表達檢測
采用免疫組化方法(SGK1免疫組化試劑盒購自Abcam公司,TLR4和NF-κB免疫組化試劑盒購自武漢華美生物工程公司),具體方法為:石蠟切片脫蠟至水,加入500mL檸檬酸鹽緩沖液,置于微波爐加熱修復抗原,PBS沖洗,室溫冷卻;加入3%過氧化氫溶液阻斷內源性過氧化物酶活性,室溫孵育20min,PBS沖洗;滴加適當比例稀釋的一抗(SGK1 1:100,TLR4 1:150,NF-κB 1:150),4℃孵育過夜,PBS沖洗,滴加適量二抗,PBS沖洗DAB,DAB顯色,蘇木素復染,水沖洗,梯度乙醇脫水,二甲苯透明,中性樹膠封片。實驗中用PBS取代一抗作為陰性對照。
1.3結果判定
1.3.1 ELISA結果判定
每個標準品和樣本的OD值與空白孔的OD值相減后作圖,以OD值為橫坐標,以標準品的濃度為縱坐標,繪制出標準曲線圖,根據樣品OD值的大小,依據標準曲線查出對應的濃度,再乘以稀釋之后的倍數即是樣品實際的濃度。
1.3.2免疫組化結果判定
每張切片隨機觀察5個400倍視野,采用染色強度與陽性細胞百分率法相結合的方法來判讀結果,二者相乘的結果作為最后得分:A.陽性細胞的染色強度按照著色強度進行評分:無著色、淡黃色、棕黃色以及棕褐色分別計分為 0~3分;B.染色細胞的數量評分標準:當染色結果為陽性的細胞數<5%時為 0分;5%~25%時為1分,>25%~50% 時為2分,>50% 為3分。A×B得分結果判斷:0~3分為陰性,4分及以上為陽性,計算每組切片的陽性率。
1.4統計學方法
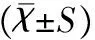
2結果
2.1兩組實驗對象臨床資料的比較
對正常早孕組及稽留流產組的臨床一般資料進行統計學分析,兩組患者在年齡、停經天數、孕次及產次均無統計學意義(均P>0.05),見表1。
表1 正常早孕組和稽留流產組臨床資料比較Table 1 Comparison of clinical data of normal pregnancy group and missed abortion group
2.2 SGK1和TLR4在患者血清中的表達
SGK1和TLR4兩種蛋白在正常早孕婦女血清中的表達量分別為(54.20±15.92)pg/mL和(8.31±2.75)ng/mL,在稽留流產患者血清中的表達量分別為(25.23±7.78)pg/mL和(15.88±6.11)ng/mL。與正常早孕組相比,SGK1在稽留流產組中的表達量顯著降低(t=8.145,P<0.05);TLR4在稽留流產組中的表達量顯著升高(t=4.919,P<0.05),差異均具有統計學意義。
2.3血清中SGK1和TLR4表達的相關性分析
Pearson相關性分析結果顯示,在稽留流產組的血清中,SGK1和TLR4的表達呈負相關(r=-0.358,P= 0.016<0.05)。
2.4 SGK1、TLR4和NF-κB在絨毛和蛻膜組織中的表達部位
免疫組化結果顯示,SGK1主要表達于絨毛合體滋養細胞的細胞質、蛻膜基質細胞細胞質和細胞膜,少數表達于細胞核。TLR4主要表達于絨毛合體滋養細胞和蛻膜基質細胞的細胞質,血管內皮細胞和絨毛間質細胞的細胞質內也有表達,少數見于細胞核。NF-κB主要表達于絨毛合體滋養細胞和蛻膜基質細胞的細胞質、細胞膜,少數見于細胞核,見圖1、圖2。
2.5 SGK1、TLR4和NF-κB在絨毛和蛻膜組織中的表達水平
在32例稽留流產中,SGK1表達陽性者11例,陽性率為34.38%(11/32)。30例正常早孕中,SGK1表達表達陽性者21例,陽性率為70.00%(21/30)。稽留流產組中SGK1表達陽性率明顯低于正常妊娠組,兩組SGK1表達陽性率比較差異有統計學意義(χ2=7.869,P<0.05)。
在32例稽留流產中,TLR4表達陽性者20例,陽性率為62.50%(20/32)。30例正常早孕中,TLR4表達陽性者6例,陽性率為20.00%(6/30)。稽留流產組中TLR4表達陽性率明顯高于正常妊娠組,兩組TLR4表達陽性率比較差異有統計學意義(χ2=11.486,P<0.05)。
在32例稽留流產中,NF-κB表達陽性者23例,陽性率為71.88%(23/32)。30例正常早孕中,NF-κB表達陽性者7例,陽性率為23.33%(7/30)。稽留流產組中NF-κB表達陽性率明顯高于正常妊娠組,兩組NF-κB表達陽性率比較差異有統計學意義(χ2=14.609,P<0.05)。

注:1A正常早孕組絨毛中SGK1的表達;1B稽留流產組絨毛中SGK1的表達;1C正常早孕組絨毛中TLR4的表達;1D稽留流產組絨毛中TLR4的表達;1E正常早孕組絨毛中NF-κB的表達;1F稽留流產組絨毛中NF-κB的表達。
圖1 SGK1、TLR4、NF-κB在正常早孕組和稽留流產組絨毛組織中的表達(免疫組化×400)
Fig.1 The expressions of SGK1, TLR4 and NF-κB in villi(免疫組化×400)

注:2A正常早孕組蛻膜中SGK1的表達;2B稽留流產組蛻膜中SGK1的表達;2C正常早孕組蛻膜中TLR4的表達;2D稽留流產組蛻膜中TLR4的表達;2E正常早孕組蛻膜中NF-κB的表達;2F稽留流產組蛻膜中NF-κB的表達。
圖2 SGK1、TLR4、NF-κB在正常早孕組和稽留流產組(免疫組化×400)
Fig. 2 The expressions of SGK1, TLR4 and NF-κB in decidua (免疫組化×400)
2.6絨毛蛻膜組織中SGK1、TLR4和NF-κB三者的相關性分析
Spearman等級相關分析結果顯示:在稽留流產組的絨毛蛻膜組織中,SGK1和TLR4的表達呈負相關(r= -0.527,P=0.003),SGK1與NF-κB的表達呈負相關(r=-0.662,P=0.000),TLR4與NF-κB的表達呈正相關(r=0.520,P=0.002)。
3討論
3.1 SGK1與母胎界面免疫微環境
SGK1參與妊娠的多個過程,包括維持子宮內膜容受性、促進囊胚植入與滋養細胞侵襲、調節母胎界面細胞分化和增殖與存活等[3]。有研究發現,與月經期子宮內膜相比,在人類早期妊娠蛻膜中,SGK1濃度明顯升高[4]。也有研究表明,在早期自然流產患者的蛻膜中,SGK1的表達減少[5]。本研究結果顯示稽留流產患者絨毛和蛻膜組織中SGK1的表達均明顯低于正常早孕組,提示SGKl低表達與胚胎停止發育有關。
妊娠期母胎界面的免疫細胞與細胞因子參與免疫網絡構成,維持免疫微環境的平衡。其中Thl/Th2平衡是維持正常妊娠的重要因素之一。早期適量的 Th1 類細胞因子如干擾素-γ(interferon-γ,IFN-γ)、腫瘤壞死因子-α(tumor necrosis factor,TNF-α)等有利于滋養細胞形成, 并在著床期促進血管生成,但Th1 類細胞因子也為促炎性細胞因子,可介導細胞免疫、急性排斥反應和炎癥反應等,過度表達時則會影響胚胎的存活,導致流產的發生[6]。
SGK1在免疫與炎癥反應中具有重要的作用。研究顯示SGK1能夠誘導分化和穩定Th2細胞,抑制 Th1 細胞的分化[7]。殷輝等[8]在研究小鼠膿毒血癥時,用脂多糖刺激SGKl抑制劑預處理過的小鼠,發現白介素-12(interleukin -12,IL-12)和TNF-α的表達較未使用SGKl抑制劑預處理的小鼠明顯升高,說明SGK1能夠抑制Th1類細胞因子的表達。
3.2 SGK1參與稽留流產發生的可能機制
國內外已有多項研究證明TLR4參與妊娠的維持和妊娠疾病的發生[9]。TLR4被激活后,通過一系列跨膜信號轉導激活NF-κB,誘導Th1型細胞因子分泌增加,抑制Th2型細胞因子分泌,破壞Thl/Th2平衡[10],導致流產的發生[11]。本研究中稽留流產組絨毛和蛻膜組織中TLR4、NF-κB的表達水平均高于正常早孕組,相關性分析顯示TLR4與NF-κB的表達成正相關,與以往結果研究一致。Zhou等[12]研究內毒素所致器官衰竭時提出SGK1可能通過抑制TLR4下游的轉化生長因子β激酶1(transforming growth factor β kinase 1,TAK1),阻止I-κB激酶(I- kappa B kinase,IKK)磷酸化,抑制NF-κB活性,負調控TLR4介導的炎性活動。本研究中稽留流產患絨毛蛻膜組織中SGK1的表達均明顯低于正常早孕組,并與TLR4、NF-κB的表達水平均呈負相關,提示SGK1低表達可能通過TLR4/NF-κB信號通路活化,導致母胎界面Th1/ Th2平衡向Th1偏倚,參與了流產的發生。
本研究同時檢測了稽留流產患者血清中SGK1和TLR4的表達及其相關性,結果顯示血清中SGK1的表達與絨毛和蛻膜組織一致,較正常早孕組明顯下降(P<0.05),而TLR4的表達較正常早孕組明顯上升(P<0.05),且二者在血清中的表達呈負相關(P<0.05),提示稽留流產患者血清中SGK1與母胎界面的表達具有一致性。檢測孕婦血清中SGK1的水平可能成為判斷流產的指標。但本研究樣本量較小,仍需大樣本研究來證實其實用性。
綜上所述,與正常早孕組相比,稽留流產患者的血清、絨毛和蛻膜組織中SGK1表達均降低,提示SGK1與稽留流產的發生具有相關性,其機制可能與SGK1表達減少,對TLR4介導的炎癥反應的負調節減弱,母胎界面免疫微環境失衡有關。SGK1有可能成為臨床檢測流產的指標或治療靶點。
[1]Xue P,Zheng M,Gong P,etal.Single administration of ultra-low-dose lipopolysaccharide in rat early pregnancy induces TLR4 activation in the placenta contributing to preeclampsia[J].PLoS One,2015,10(4):e0124001.
[2]Lou Y,Hu M,Wang Q,etal.Estradiol suppresses TLR4-triggered apoptosis of decidual stromal cells and drives an anti-inflammatory TH2 shift by activating SGK1[J].Int J Biol Sci,2017,13(4):434-448.
[3]Lou Y,Hu M,Mao L,etal.Involvement of serum glucocorticoid-regulated kinase 1 in reproductive success[J].FASEB J,2017,31(2):447-456.
[4]Lv Y,Gao S,Zhang Y,etal.miRNA and target gene expression in menstrual endometria and early pregnancy decidua[J].Eur J Obstet Gynecol Reprod Biol,2016,197:27-30.
[5]Wang Y,Lv Y,Wang L,etal.MicroRNAome in decidua: a new approach to assess the maintenance of pregnancy[J].Fertil Steril,2015,103(4):980-989.
[6]李莉,喬杰,王海燕.不明原因復發性流產免疫學發病機制的研究進展[J].中華生殖與避孕雜志,2017, 37(2):160-165.
[7]Heikamp E B,Patel C H,Collins S,etal.The AGC kinase SGK1 regulates TH1 and TH2 differentiation downstream of the mTORC2 complex[J].Nat Immunol,2014,15(5):457-464.
[8]殷輝,寧會彬,曾艷麗,等.血清和糖皮質激素調節蛋白激酶1通過核因子-κB抑制Toll樣受體4介導的炎性反應[J].中華傳染病雜志,2016,34(4):242-247.
[9]Koga K,Izumi G,Mor G,etal.Toll-like receptors at the maternal-fetal interface in normal pregnancy and pregnancy complications[J].Am J Reprod Immunol,2014,72(2):192-205.
[10]Wei C B,Tao K,Jiang R,etal.Quercetin protects mouse liver against triptolide-induced hepatic injury by restoring Th17/Treg balance through Tim-3 and TLR4-MyD88-NF-κB pathway[J].Int Immunopharmacol,2017,53:73-82.
[11]趙先蘭,李莎.稽留流產患者絨毛組織中TLR4、NF-κB和HBD2的表達及其相關性[J].中華婦產科雜志,2011,46(11):864-866.
[12]Zhou H,Gao S,Duan X,etal.Inhibition of serum- and glucocorticoid-inducible kinase 1 enhances TLR-mediated inflammation and promotes endotoxin-driven organ failure[J].FASEB J,2015,29(9):3737-3749.
[專業責任編輯:于學文]
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中老年保健(2021年3期)2021-08-22 06:50:04
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
